Особенности железистого аппарата растений Mentha arvensis L. разного географического происхождения

Автор: Шелепова Ольга Владимировна, Воронкова Татьяна Владимировна, Смирнова Ирина Михайловна, Енина Ольга Леонидовна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Флористика
Статья в выпуске: 1-7 т.14, 2012 года.
Бесплатный доступ
Изучены параметры секреторного аппарата 3 разновидностей, 1 формы и 2 сортов, относящихся к M. arvensis L. разного географического происхождения. Железистый аппарат представлен 10-и клеточными секреторными железками и 3-х клеточными волосками. Наиболее крупными эфиромасличными трихомами и высокой плотностью их расположения на обеих сторонах листа отличалась тетраплоидная форма M. arvensis из природной флоры Вьетнама. Это обеспечило наибольший выход эфирного масла (4,73%). По количеству, размерам и плотности эфиромасличных секреторных желез наиболее перспективными для селекционной работы являются тетраплоидная форма M. arvensis из природной флоры Вьетнама и M. arvensis var. piperascens из природной флоры острова Тайвань.
Секреторные железки, эфирное масло
Короткий адрес: https://sciup.org/148200952
IDR: 148200952 | УДК: 582.949/665.527
Features of the glandular apparatus of Mentha arvensis L. plants of different geographical origin
The parameters of the secretory apparatus of three species, one form and two varieties belonging to M. arvensis L. different geographical origin were studied. Glandular apparatus is represented by a 10-cell secretory glands and 3-cell hairs. The major aromatic trichomes and a high density of their arrangement on both sides differed tetraploid form of M. arvensis of the natural flora of Vietnam. This provided the highest yield of essential oil (4.73%). By the number, size and density of the essential oil secretory glands of the most promising for breeding are tetraploid form of M. arvensis of the natural flora of Vietnam and M. arvensis var. piperascens of the natural flora of the island of Taiwan.
Текст научной статьи Особенности железистого аппарата растений Mentha arvensis L. разного географического происхождения
Mentha arvensis L. – вид широко распространенный во всех областях лесной, лесостепной и степной зон Европейской части России и Западной Сибири, имеет евроазиатский бореальный тип распространения. Для вида характерна значительная экологическая и географическая изменчивость. В своем исследовании «Дикорастущие мяты СССР» (1972) В.В. Макаров выделил 11 разновидностей данного вида, вид крайне полиморфен [3]. Кроме того в естественных условиях часто возникают межвидовые гибриды между M. arvensis и M. aquatica , и M. arvensis и M. longifolia .
M. arvensis – один из видов рода Mentha, широко используемых в мировом производстве для получения эфирного масла с высоким содержанием ментола. Основной фонд продуцируемого мятой эфирного масла формируется в железистом аппарате растения. Это специфическая по своей структуре и функциональной диагностике система. Секреторные железы у растений мяты образуются на ранних этапах онтогенез а на всех органах на всех органах надземной части растений (стеблях, листьях, чашечке, венчике). Кроме того, секреторные железы обнаружены на поверхности корневищ мяты [1]. Для растений мяты характерны 2 типа секреторных образований: секреторные волоски и сидячие 10клеточные железки. Основной фонд продуцируемого растением эфирного масла сосредоточен именно в секреторных железках (трихомах). Наибольшая их плотность наблюдается на листьях возраста от 12 до 20 дней, причем максимальное их количество расположено в базальной части листа, на средней и апикальной частях – меньше. Более высокая плотность трихом выявлена на нижней части листьев, она возрастает от листьев нижнего яруса к листьям верхнего яруса [2, 5].
Формы, гибриды и сорта, относящиеся к разным видам рода Mentha, отличаются параметрами железистого аппарата. Так, согласно данным [4] наиболее крупные трихомы наблюдались у M. piperita (70,5-76,5 мкм), самые мелкие - M. aquatica (60,061,1 мкм).
Крайний полиморфизм M. arvensis позволяет предположить, что для разновидностей, гибридов и сортов, относящихся к данному виду, также будет характерно варьирование параметров железистого аппарата.
Целью данной работы было изучение особенностей железистых трихом 3 разновидностей, 1 формы и 2 сортов, относящихся к M. arvensis . Наши исследования позволяют заполнить недостающие параметры в характеристиках образцов, что даст возможность более эффективного их использования в практической селекции.
МАТЕРИАЛЫ И МЕТОДЫ
Коллекция видов, гибридов и сортов мяты сформировалась в лаборатории физиологии и биохимии растений ГБС РАН в течение последних лет, но ряд сортов и гибридов мяты культивируются с середины 80-х годов прошлого столетия. Новая плантация мяты была заложена весной 2010 г. на экспозиционном участке лаборатории в Москве, растения высаживали черенками с 3-4 парами развитых листьев на делянки по 1 м2 и выращивали при дополнительном поливе в течение вегетационного периода.
Объектами исследования были 3 разновидности M. arvensis: № 1 – var. lapponica (возможно var. arvensis, так как никаких надежных признаков отделения нет) [3] из коллекции Института биологии Коми научного центра УрО РАН (Сыктывкар, Россия); № 2 – var. piperascens из природной флоры Дальневосточного региона; № 3 – var. piperascens из природной флоры острова Тайвань; № 4 – сорт «Памяти Кириченко» (в дальнейшем «ПК»), внесенный в Государственный реестр селекционных достижений 56523/8853145; № 5 – сорт «NT» (оба сорта получены на основе аналитической селекции из образцов M. arvensis природной флоры Вьетнама); № 6 – тетраплоидная форма, полученная обработкой M. arvensis из природной флоры Вьетнама раствором колхицина.
Изучение секреторного аппарата проводили на растениях 2 года вегетации. Для подсчета количества и определения размера секреторных железок были отобраны по 20 штук полностью сформировавшихся листьев 12-14 ярусов от верхней части побега, железки на которых находились в постсекреторной стадии развития [6, 7]. Кроме того, были учтены площадь поверхности и биомасса листа. Затем 20 участков каждого листа с обеих сторон были сфотографированы с помощью видеокамеры Lumenera Infinity 2, присоединенной к световому микроскопу, при увеличении Ч500 и обработаны при помощи программы Infinity camera V:5.0.2. После соответствующей калибровки масштаба на каждом фотоснимке было подсчитано количество секреторных железок на верхней и нижней поверхности листа и определен их диаметр. Полученные результаты были обработаны статистически и рас- считаны к средней площади листа для каждого объекта исследования.
Фенологические наблюдения, учет биомассы и выхода эфирного масла проводили в 2010-2011гг. на растениях 1 и 2 годов вегетации. Определение содержания эфирного масла проводили методом гидродистилляции.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Анализ полученных данных показал, что наиболее крупные листья яйцевидно-округлой формы сформировались у тетраплоидной формы M. arven-sis – средняя площадь листа составила 16,5±0,3 см, соотношение длины к ширине листа – 1,7 (табл. 1). У сортов «ПК» и «NT» размеры листьев были в 2,12,5 раза меньше и они имели яйцевидную форму (соотношение длины к ширине листа 2,0-2,7). И наиболее мелкие листья зафиксированы у всех 3 разновидностей M. arvensis – средняя площадь листа не превышала 5,0 см2, была в 3,6-4,2 раза меньше, чем у тетраплоидной формы, листья имели яйцевидную форму (соотношение длины к ширине листа 2,4-2,5).
Таблица 1 . Отдельные параметры секреторного аппарата и биопродуктивности разновидностей, форм и сортов M. arvensis различного географического происхождения
|
Номер образца |
Площадь листа, см2 |
Отношение длины к ширине листа |
Количество трихом на листе, шт |
Биомасса ли стьев с 1м2 |
Выход эфирного масла, % на возд.-сухое в-во |
|
|
Нижняя по верхность |
Верхняя по верхность |
|||||
|
№ 1 |
4,4±0,8 |
1,92 |
918 |
968 |
56,3 |
0,99 |
|
№ 2 |
3,9±0,5 |
2,35 |
3062 |
1502 |
202,3 |
2,04 |
|
№ 3 |
4,6±0,7 |
2,52 |
4433 |
1834 |
146,3 |
2,51 |
|
№ 4 |
8,1±0,6 |
2,68 |
8536 |
3837 |
314,8 |
3,00 |
|
№ 5 |
6,5±0,7 |
2,04 |
6389 |
2671 |
260,6 |
1,93 |
|
№ 6 |
16,5±1,2 |
1,70 |
18942 |
7244 |
265,5 |
4,73 |
Практически у всех изученных растений количество секреторных трихом на нижней поверхности листа было в 2,0-2,6 раза больше, чем на верхней. Только у var. lapponica на обеих сторонах листа зафиксировано практически одинаковое количество секреторных желез (~ 200 шт/см2) и минимальное их количество на листе (~ 970 шт).
Для характеристики железистого аппарата также важна плотность их расположения на поверхности листа. Наиболее высокой плотностью расположения трихом желез на обеих сторонах листа отличалась тетраплоидная форма M. arvensis (рис. 1). У сортов «ПК» и «NT» она была в 2,3-3,0 (для нижней стороны) и 1,9-2,7 (для верхней) раза меньше, а у разновидностей в 4,3-20,6 и 3,9-7,5, соответственно, раза меньше по сравнению с тетраплоидной формой.
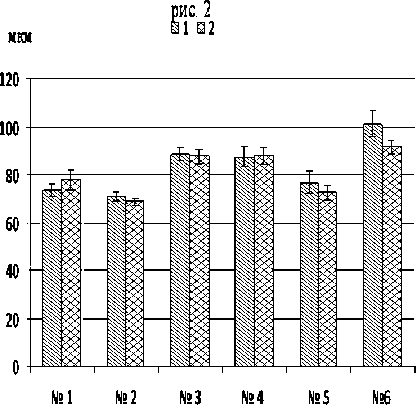
Максимальный размер секреторных желез на обеих сторонах листа зафиксирован у тетраплоид-ной формы – 101,3±5,4 (нижняя) и 91,5±3,2 (верхняя) мкм (рис. 2), что несколько выше показателей характерных для M. piperita [4]. Минимальный размер – у var. piperascens из природной флоры Дальневосточного региона – диаметр трихом не превышал 71 мкм.
В целом аналогичные размеры секреторных трихом на обеих сторонах листа характерны для сорта «ПК» и var. piperascens из природной флоры Дальневосточного региона и острова Тайвань. Только у var. lapponica на верхней поверхности листа сформировались более крупные железы.
Из всех изученных нами разновидностей, форм и сортов, относящихся к M. arvensis , как нам кажется, следует обратить внимание на 3. Это – тет-раплоидная форма M. arvensis , var. piperascens из природной флоры острова Тайвань и сорт «ПК», полученный методом аналитической селекции из M. arvensis из природной флоры Вьетнама.
Тетраплоидная форма M. arvensis – растения прямостоячие с ярко, интенсивно зеленой окраской, практически отсутствует опушенность на стебле. Листья крупные яйцевидно-овальной формы. Чашечка трубчато-колокольчатая, опушенная. Венчик бледно-розового цвета.
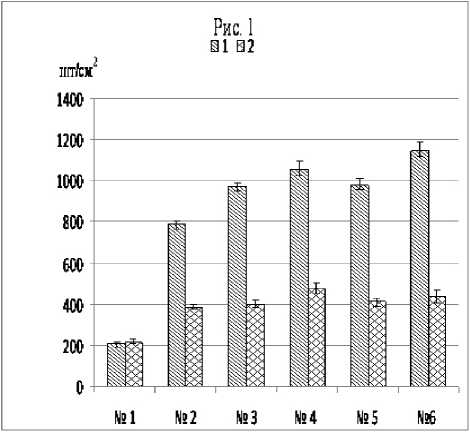
Рис. 1 . Плотность расположения секреторных железок на нижней (1) и верхней (2) сторонах листа растений M. arvensis разного географического происхождения: № 1 - var. lapponica ; № 2 - var. piperascens из природной флоры Дальневосточного региона; № 3 - var. piperascens из природной флоры острова Тайвань; № 4 - сорт «Памяти Кириченко»; № 5 - сорт «NT»; № 6 - тетраплоидная форма M. arvensis .
Рис. 2 . Размеры секреторных железок на нижней (1) и верхней (2) сторонах листа растений M. arvensis разного географического происхождения: № 1 - var. lapponica ; № 2 - var. piperascens из природной флоры Дальневосточного региона; № 3 - var. piperascens из природной флоры острова Тайвань; № 4 - сорт «Памяти Кириченко»; № 5 - сорт «NT»; № 6 - тетраплоидная форма M. arvensis .
Лист наряду с железистыми волосками имеет 10 клеточные трихомы, в субкутикулярном пространстве головок которых формируется основной фонд продуцируемого растением эфирного масла. Они выглядят под микроскопом как блестящие шарики, располагающиеся в небольших углублениях ткани эпидермиса листа. Идеальную форму шара зрелым железкам придает внутреннее давление эфирного масла, содержащееся в этих полостях [6, 7]. Размеры и густота расположения трихом при максимальной площади листьев позволили обеспечить наибольший выход эфирного масла - 4,73 %, при достаточно высокой продуктивности биомассы листьев. По количеству, размерам и плотности эфиромасличных секреторных желез данная форма является наиболее перспективной для селекционной работы.
M. arvensis var. piperascens из природной флоры острова Тайвань - растение прямостоячее с ярко, интенсивно зеленой окраской, стебель с легким красноватым оттенком, опушен. Листья овальноланцетные, удлиненно овальные, длина пластинок превышает ширину в 2,5 раза, на очень коротких черешках.
Анализ данных показал, что для данной разновидности характерны максимальные практически одинаковые размеры на нижней и верхней поверхностях листа секреторных желез, что обеспечило высокий выход эфирного масла (2,51 %) при средней продуктивности биомассы листьев. Данную разновидность также следует признать перспективной для селекционной работы.
Сорт «Памяти Кириченко», полученный из M. arvensis из природной флоры Вьетнама - растения прямостоячие с ярко, интенсивно зеленой окраской, стебель ребристый, практически голый. Листья овально-ланцетные, удлиненно овальные, на очень коротких черешках. Чашечка трубчатоколокольчатая, опушенная. Венчик бледнорозового цвета.
Сформированные на листья секреторные железы имели достаточно значительные размеры при высокой плотности их расположения на обеих поверхностях листа. У данного сорта отмечена максимальная продуктивность биомассы листьев при достаточно высоком содержании эфирного масла (3,0 %), что позволяет рекомендовать данный сорт для интродукции в условиях разных почвенноклиматических районов России.
ЗАКЛЮЧЕНИЕ
Изучены параметры секреторного аппарата 3 разновидностей, 1 формы и 2 сортов, относящихся к M. arvensis разного географического происхождения. Железистый аппарат изученных разновидностей, формы и сортов в основном представлен 10-и клеточными секреторными железками наряду с 3-х клеточными волосками. Наиболее крупными эфиромасличными трихомами отличалась тетрап-лоидная форма M. arvensis из природной флоры Вьетнама. Для нее также характерна высокая плотность расположения секреторных желез на нижней и верхней поверхностях листа, что обеспечило наибольший выход эфирного масла (4,73 %).
По количеству, размерам и плотности эфиромасличных секреторных желез наиболее перспективными для селекционной работы являются тет-раплоидная форма M. arvensis из природной флоры Вьетнама и M. arvensis var. piperascens из природной флоры острова Тайвань.
Работа выполнена при финансовой поддержке гранта РФФИ № 11-04-01820.
Список литературы Особенности железистого аппарата растений Mentha arvensis L. разного географического происхождения
- Кириченко Е.Б. Экофизиология мяты. М.: Наука, 2008. 140 с.
- Лысякова Н.Ю. Некторые морфо-физиологические особенности межвидовых гибридов мяты, определяющие уровень накопления эфирного масла: дис…. канд. сел.-хоз. наук. Симферополь, 1984. 213с.
- Макаров В.В. Дикорастущие мяты СССР: дисс… канд. биол. наук. М., 1972. 179с.
- Теплицкая Л.М., Резникова С.А. Исследование железистого аппарата у мяты в связи с задачами селекции//Труды ВИР. 1975. Т.54, вып.2. С.262-272.
- Теплицкая Л.М. Особенности морфологии и дифференцировки железистого аппарата мяты в связи с интенсивностью и направленностью маслообразовательного процесса: автореф. дис…. канд. биол. наук. Кишинев, 1982. 23с.
- Turner Gl. W., Gershenzon J., Croteau R.B. Development of peltate glandular trichomes of peppermint.//Plant. Physiol. 2000. V.124. P. 665-679.
- Turner Gl. W., Gershenzon J., Croteau R.B. Distribution of peltate glandular trichomes on developing leaves of peppermint.//Plant. Physiol. 2000. V.124. P. 655-664.
