Особенности жизненного цикла и пространственно-демографической структуры ценопопуляций полыни крымской (Artemisia taurica Willd.) в Республике Калмыкия
 в Республике Калмыкия")
Автор: Зенкина Татьяна Евгеньевна, Сагалаев Вадим Александрович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: флора
Статья в выпуске: 1-9 т.14, 2012 года.
Бесплатный доступ
Обсуждаются результаты исследования ценопопуляций полыни крымской в Кумо-Манычской впадине (Республика Калмыкия). Описываются особенности жизненного цикла и возрастной структуры ценопопуляций, характеризуется адаптивная роль пространственно-демографической структуры ценопопуляций в условиях меняющихся факторов окружающей среды.
Ценопопуляции, возрастные состояния, демографические спектры, пространственная структура
Короткий адрес: https://sciup.org/148201039
IDR: 148201039 | УДК: 582.661.15(470.47)
Features of life cycle and spatial-demographic structure of wormwood crimean (Artemisia taurica Willd.) cenopopulation in Kalmykia Republic
Results of research the wormwood crimean cenopopulation in Kumo-Manych hollow (Kalmykia Republic) are discussed. Features of life cycle and age structure of cenopopulation are described, the adaptive role of spatial and demographic structure of cenopopulation in the conditions of changing factors of environment is characterized.
Текст научной статьи Особенности жизненного цикла и пространственно-демографической структуры ценопопуляций полыни крымской (Artemisia taurica Willd.) в Республике Калмыкия
Исследования ценопопуляций (ЦП) растений являются основой для разработки научных рекомендаций по сохранению биологического разнообразия и природных экосистем в изменяющихся условиях окружающей среды. Изучение организации популяций позволяет оценить их состояние, определить способы самоподдержания и устойчивого существования вида. Знание биологии вида и структуры его популяций – необходимая основа для прогноза дальнейшего развития популяций и реакции растений на факторы среды обитания [2, 4, 7, 9-12].
Цель исследования: изучение особенностей онтоморфогенеза полыни крымской ( Artemisia taurica Willd . ), которая является важным ценозооб-разователем в северо-западном Прикаспии [8].
Оценивалась пространственная структура разновозрастных ЦП данного вида. Работы проводились в районах добычи нефти в Кумо-Манычской впадине на территории Черноземель-ского района Республики Калмыкия.
Материалы и методы. Artemisia taurica относится к группе стержнекорневых каудексовых травянистых поликарпиков с моноциклическими косовертикальными удлиненными полуразеточ-ными монокарпическими побегами. Флоральной единицей полыни крымской служит одиночная корзинка. При описании структурных особенностей генеративных побегов выделяют фрагмент синфлорисценции – кисть из корзинок [1]. Вид характеризуется наличием двух форм соцветий с поникающими и прямостоячими корзинками [1, 5]. A. taurica является эдификатором или доминантом многих сообществ, так называемых «таврико-
полынников», развитых на светло-каштановых, каштановых и черноземных карбонатных почвах, как правило, солонцеватых. Эти сообщества вторичны: они развиты на залежах большой давности и сильно выпасаемых участках [6]. Работы проводились в полевые сезоны 2009-2012 гг. по классической методике изучения пространственной и демографической структуры популяций растений [4, 7, 10-12]. Для оценки горизонтальной проекции особей растений в ЦП закладывались площадки 3х3 м в различных участках катены.
Полученные результаты и их обсуждение. Жизненный цикл полыни крымской (A. taurica). Изученный жизненный цикл A. taurica включает 3 онтогенетические стадии (прегенеративная, генеративная, сенильная) и 8 возрастных состояний: проростки ( р ), ювенильные ( j ), имматурные ( im ), виргинильные ( v ), молодые генеративные ( g 1 ), средневозрастные-старые генеративные ( g 2 -g 3 ), сенильно-генеративные ( sg ), сенильные ( s ). Наблюдения в природе показывают, что длительность каждого возрастного состояния может колебаться в значительной степени в зависимости от климатических условий конкретного года. В 2010 г. фиксировались все возрастные состояния преге-неративной стадии ( p, j, im, v ), продолжительность каждой из которых можно оценить в один год; пре-генеративная стадия, таким образом, длилась 4 года. В 2011 г. в связи с аномально влажным летним периодом осенью наблюдалось зацветание и плодоношение виргинильных ( v ), имматурных ( im ) и даже ювенильных особей ( j ). Вероятно, этот эффект неотенизации и зацветания прегенеративных растений (т. е. переход их в g 1 ) может быть связан с возможностью дополнительного синтеза и накопления запаса пластических веществ, необходимых для закладки корзинок на моноциклических побегах, сформировавшихся в течение вегетационного сезона.
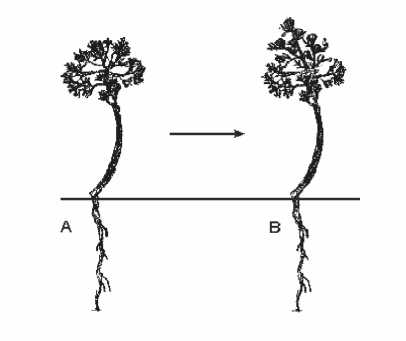
Рис. 1. Схема перехода имматурной особи ( im ) в генеративную стадию в течение вегетационного сезона 2011 г.: А – вегетирующая имматурная ( im ) особь весной; В – зацветающая имматурная ( im ) особь, перешедшая в стадию генерации ( g 1 ) осенью
Аналогично происходит переход из прегене-ративной ( j, v ) в стадию генерации. По-видимому, возможность перехода в генеративную стадию, минуя очередные возрастные состояния прегенера-тивной стадии, в определённых условиях позволяет виду увеличивать семенную продуктивность, пополнять банк семян в почве и повышает его конкурентную способность. Подобное явление неоте-низации наблюдается и у некоторых других видов полыней подрода Seriphidium Bess. Например, Б.А. Келлер наблюдал зацветание особей полыни черной ( Artemisia pauciflora Web.) в благоприятных условиях выращивания уже осенью первого года жизни [13]. Возможно, что неотенизация встречается и среди других полыней этого подрода. Наблюдаемый процесс «пропуска» отдельных возрастных состояний можно интерпретировать как поливариантность этапов онтогенеза [3].
Переход к генеративной стадии выражается в формировании на моноциклическом побеге многочисленных одиночных корзинок, образующих синфлоресценцию [1]. Средневозрастное генеративное ( g 2 ) и старое генеративное ( g 3 ) состояния на собранном материале различить не представляется возможным, поэтому они объединены в средневоз-растное-старое генеративное ( g 2 -g 3 ) состояние. Календарный возраст генеративной стадии может быть оценен в 3-6 и более лет. Нам удалось обнаружить переходное возрастное состояние между генеративной и сенильной стадиями. Это сенильно-генеративное возрастное состояние ( sg ) характеризуется наличием партикуляции куста, сопровождающейся отмиранием части побегов, но с сохранением возможности формирования у отдельных партикул генеративных монокарпических побегов. Длится такое возрастное состояние 2-3 года. Сенильное возрастное состояние ( s ) наблюдается у партикулирующих особей, неспособных формировать генеративные структуры. Сенильная стадия может продолжаться1-2 и более лет. Таким образом, общая продолжительность жизненного цикла
-
A. taurica может быть оценена в 7-17 лет. Широкий временной диапазон максимальной и минимальной продолжительности жизни полыни крымской свидетельствует о высокой пластичности вида и его значительных адаптационных возможностях.

Рис. 2. Горизонтальная проекция участка «молодой» ЦП А. taurica
Пространственно-демографическая структура ЦП A. taurica. Анализ результатов изучения пространственно-демографической структуры ЦП полыни крымской в районе исследования позволил выявить две их условные группы: «молодые» и «зрелые». В ходе исследований «стареющая» группа практически не отмечалась, т.к. в результате интенсивного выпаса скота большая часть особей находящихся в средневозрастном-старом генеративном ( g 2 -g 3 ) возрастном состоянии были съедены пасущимися овцами. Отдельные сенильные ( s ) и сенильно-генеративные особи ( sg ) встречались рассеяно по небольшим депрессиям и неровностям рельефа. Горизонтальная проекция участка «молодой» ЦП А. taurica представлена на рис. 2.
Онтогенетические состояния
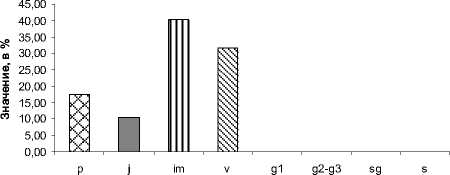
Рис. 3. Возрастной спектр «молодой» ЦП А. taurica
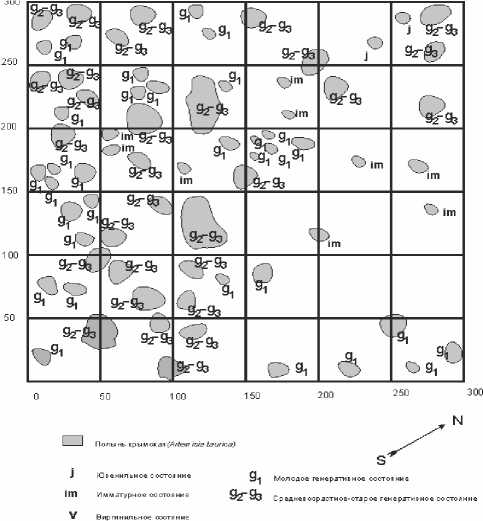
Рис. 4. Горизонтальная проекция участка «зрелой» ЦП А. taurica
В данной ЦП доминируют особи в вир-гинильном ( v ) и имматурном ( im ) состоянии с небольшим преобладанием последних. Возрастной спектр «молодой» ЦП показан на рис. 3. Он является левосторонним, с одним максимумом, приходящимся на имматурное возрастное состояние ( im ) .
Следующим этапом развития ЦП является ее «зрелое» состояние. В подобных ЦП растения находятся преимущественно в молодом генеративном ( g 1 ), средневозрастном-старом генеративном состоянии ( g 2 -g 3 ). Здесь также часто встречаются имматурные ( im ) особи. Горизонтальная проекция пространственной структуры такой ценопопуляции показана на рис. 45.
Возрастной спектр «зрелой» ЦП характеризуется наличием двух максимумов приходящихся на g 1 и g 2 -g 3 состояния (рис. 5).
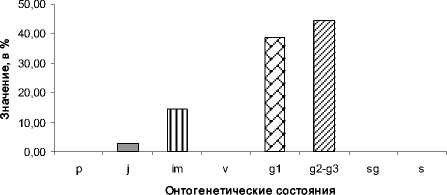
Рис. 5. Возрастной спектр «зрелой» ЦП А. taurica
Базовый возрастной спектр всех типов популяций полыни крымской показан на рис. 6. Он характеризуется сочетанием двух максимумов, приходящихся на виргинильное состояние (v) и средневозрастное-старое генеративное состояние (g2-g3).
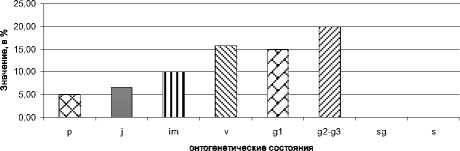
Рис. 6. Базовый онтогенетический спектр ЦП А. taurica
Небольшая доля проростков в базовом спектре полыни объясняется интенсивным выпасом скота в районе исследования и поздними сроками наблюдения (август-октябрь). В осенний период можно точно идентифицировать и наблюдать почти все возрастные состояния, но проростки в это время наблюдать затруднительно, т.к. опадают семядоли, и тогда отличить проросток от ювенильной особи очень сложно.
Выводы:
-
1. Жизненный цикл полыни А. taurica включает 3 онтогенетические стадии (прегенеративная, генеративная, сенильная) и 8 возрастных состояний.
-
2. В ходе онтогенеза полыни крымской наблюдается явление неотенизации, выражающееся в «пропуске» отдельных возрастных состояний пре-генеративной стадии; наблюдаемое явление может быть интерпретировано и как поливариантность этапов онтогенеза.
-
3. Пространственно-демографическая структура ЦП в районе исследований характеризуется почти полным отсутствием сенильной стадии, это вызвано высоким уровнем пастбищной нагрузки на це-нопопуляции.
-
4. Базовый спектр ЦП является неполночленным из-за почти полного отсутствия сенильной стадии и слабой представленности в спектре проростков. Это связано с высоким уровнем антропогенной нагрузки на ЦП полыни.
-
5. Оценивая в целом состояние изученных ЦП А. taurica , следует отметить высокий уровень их устойчивости и адаптированность к различным факторам экзогенной природы.
Список литературы Особенности жизненного цикла и пространственно-демографической структуры ценопопуляций полыни крымской (Artemisia taurica Willd.) в Республике Калмыкия
- Беэр, А.С. Сравнительное биоморфологическое исследование восточноевропейских представителей рода Artemisia L. (Asteraceae Dumort.)/А.С. Беэр: Дис. канд. биол. наук. -М., 2005. 424 с.
- Жукова, Л.А. Популяционная жизнь луговых растений/Л.А. Жукова. -Йошкар-Ола: «Ланар», 1995. 224 с.
- Жукова, Л.А. Поливариантность онтогенеза и динамика ценопопуляций растений/Л.А. Жукова, А.С. Комаров//Журнал общей биологии. 1990. Т. 51, № 4. С.450-461.
- Заугольнова, Л.Б. Структура популяций семенных растений и проблемы их мониторинга/Л.Б. Заугольнова: автореф. дис. докт. биол. наук. -СПб., 1994. 70 с.
- Куваев, В.Б. Об одной разновидности полыни таврической (Artemisia taurica Willd.)/В.Б. Куваев, Д.А. Пакалн//Новости систематики высших растений. Т. 8. -Л.: Наука, 1971. С. 240-243.
- Левина, Ф.Я. Новые данные к ареалу полыни Artemisia taurica Willd//Ботанический журнал. 1963. Т. 58, № 3. С. 49-52.
- Работнов, Т.А. Вопросы изучения состава популяций для целей фитоценологии/Т.А. Работнов//Проблемы ботаники. Вып. 1. -М.-Л., 1950. С. 465-483.
- Сафронова, И.Н. Пустыни как тип растительности. Северо-туранские (прикаспийские) пустыни/И.Н. Сафронова//Растительность европейской части СССР. -Л.: Наука, 1980. С. 285-291; 291-295.
- Смирнова, О.В. Структура травянистого покрова широколиственных лесов/О.В. Смирнова. -М.: Наука, 1987. 207 с.
- Уранов, А.А. Возрастной спектр фитоценопопуляций как функция времени и энергии волновых процессов/А.А. Уранов//Биол. науки. 1975. № 2. С.7-34.
- Ценопопуляция растений (основные понятия и структура). -М.: Наука, 1976. 217 с.
- Ценопопуляции растений (очерки популяционной биологии). -М.: Наука, 1988. 184 с.
- Keller, B.А. Die Methoden zur Erforschung der Ökologie der Steppen-und Wüstenpflanzen/B.A. Keller//Handbuch der biologischen Arbeitsmethoden (ed E. Abderhalden). Berlin: Urban und Schwarzenberg, 1930. 1-128 S.
