Отбор солеустойчивых растений разных видов люцерны ( Medicago L.) и анализ их морфобиологических и симбиотрофных показателей
 и анализ их морфобиологических и симбиотрофных показателей")
Автор: Румянцева М.Л., Степанова Г.В., Курчак О.Н., Онищук О.П., Мунтян В.С., Дзюбенко Е.А., Дзюбенко Н.И., Симаров Б.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Экологические основы безопасных агротехнологий
Статья в выпуске: 5 т.50, 2015 года.
Бесплатный доступ
Усиливающаяся деградация почв связана с истощением их плодородия в результате применения севооборота с внесением избыточного количества минеральных удобрений и химических средств защиты растений, а также в связи с повсеместно ухудшающимися природно-климатическими условиями и экологической обстановкой. Именно поэтому сельское хозяйство, основанное на экологически безопасных агротехнологиях, должно иметь абсолютный приоритет. Бобовые культуры фиксируют азот атмосферы в симбиозе с клубеньковыми бактериями и накапливают его в биомассе растений. Они служат уникальными предшественниками для выращивания зерновых, поскольку способствуют восстановлению плодородия почв за счет привнесения азота в биологически доступной форме. Создание пастбищ на основе бобовых трав благоприятствует восстановлению почв, выведенных из севооборота (например, опустыненных или засоленных). Поэтому разработка приемов создания продуктивных растительно-микробных систем, которые эффективны в неблагоприятных условиях, имеет огромное теоретическое и практическое значение. В задачи исследования входило выявление солеустойчивых форм люцерны ( Medicago L.), получение на их основе растений поколения I 1 методом принудительного самоопыления и анализ морфологических, биологических и симбиотических показателей в модельных опытах. Объектами изучения были 13 тетраплоидных и диплоидных сортов люцерны, в том числе сорта Солеустойчивая и Агния, которые испытывали при симбиотрофном питании (в симбиозе со штаммами Sinorhizobium meliloti ) или без инокуляции штаммами. Анализ симбиотической активности изученных сортов показал, что они высокоотзывчивы на инокуляцию тест-штаммом S. meliloti Rm2011 и могут формировать эффективные симбиозы в условиях засоления. Была оценена однородность географически различных сортов по накоплению сухого вещества (СВ) на фоне 75 мМ NaCl без инокуляции и 100 мМ NaCl с инокуляцией S. meliloti с привлечением коэффициента дисперсии (Д), что позволило установить, что показатель СВ у изучаемых сортов существенно изменялся только в случае симбиотрофного питания. По результатам анализа симбиотической активности в присутствии соли в микровегетационных опытах (Всероссийский НИИ сельскохозяйственной микробиологии - ВНИИСХМ) получены солеустойчивые формы у растений диплоидных видов М. caerulea и М. falcata, а также у тетраплоидного вида M. sativa L. сортов Солеустойчивая и Агния. Отобранные растения сортов Солеустойчивая и Агния выращивали далее в селекционно-тепличном комплексе (СТК) Всероссийского НИИ кормов им. В.Р. Вильямса. Проведенные 3-летние вегетационные испытания в СТК показали, что для солеустойчивых растений сортов Солеустойчивая и Агния характерна преимущественно сиреневая окраска цветов, более закрученная форма боба, сравнительно высокая ветвистость и кустистость, более поздний переход в период покоя. Из семян от испытуемых растений в микровегетационном опыте получили растения поколения I 1 (ВНИИСХМ). Растения I 1 анализировали по СВ, интенсивности роста надземной и подземной частей, по числу сформированных клубеньков в стандартных (бессолевые) условиях и при засолении. Установлено, что в поколения I 1 у обоих сортов растения однородны или достаточно однородны по СВ. В присутствии соли у сорта Солеустойчивая растения поколения I 1 успешно развивались без инокулянта (усредненная прибавка СВ составила 36,92 % относительно исходных растений), тогда как у сорта Агния прибавка СВ в I 1 была, наоборот, на 16,55 % ниже. У обоих сортов выявлена высокая специфичность взаимодействия растений I 1, контрастно различавшихся по солеустойчивости, оцениваемой по СВ, с симбиотически высокоэффективными штаммами CIAM1774 или Rm2011. Так, в условиях солевого стресса наибольшие значения СВ получены у сорта Солеустойчивая при инокуляции штаммом Rm2011 с солеустойчивостью, типовой для S. meliloti. Однако симбиотическая система на основе полученного нами солеустойчивого генотипа сорта Агния с сол
Клубеньковые бактерии, люцерна, эффективность симбиоза, солеустойчивость, длина корня и стебля, масса растений
Короткий адрес: https://sciup.org/142133631
IDR: 142133631 | УДК: 633.31:581.557:631.461.5:57.044 | DOI: 10.15389/agrobiology.2015.5.673rus
Selection of salt tolerant alfalfa ( Medicago L.) plants from different varieties and their morfo biological and symbiotic properties analysis
Soils degradation growing in our days is associated with the depletion of their fertility, a result of crop rotation with excessive amounts of mineral fertilizers and chemical plant protection products, as well as it links with widespread worsening climatic conditions and environmental conditions. For this reason, agriculture based on environmentally friendly technologies must be an absolute priority. Legumes can fix atmospheric nitrogen in symbiosis with nodule bacteria and accumulate it in plant biomass. Legumes are unique predecessors for grain crops, as they contribute to the effective restoration of soil fertility by introducing nitrogen into bioavailable form. Pastures based on legumes contribute to the restoration of soils destroyed and excluded from crop rotation, such as desert or saline. In this, the development of pathways to create new productive plant-microbe systems that can grow in adverse conditions, is of great theoretical and practical significance. The objectives of the study was to identify salt-tolerant plants of alfalfa ( Medicago L.), to obtain plants of the I 1 generation by self-pollination approach and to analyse their morphobiological and symbiotic properties in model experiments. The study was performed on 13 tetraploid and diploid varieties of alfalfa, including commercially valuable varieties Soleustoychivaya and Agniya, both of which were tested without rhizobia inoculation and in symbioses with Sinorhizobium meliloti strains. An analysis of the symbiotic activity of alfalfa varieties showed that they were highly responsible to S. meliloti Rm2011 strain inoculation and formed an effective symbiosis under saline conditions. Geographically different varieties were evaluated for the homogeneity according to dry matter (DM) accumulation at 75 mM NaCl without inoculation, and at 100 mM NaCl with inoculation by S. meliloti. Obtained DM data among the studied cultivars significantly changed only in case of symbiosis that was established with the assistance of the dispersion coefficient (D). Plants of salt tolerant phenotype was obtained for diploid M. caerulea and M. falcata species, as well as for tetraploid M. sativa L. varieties Soleustoychivaya and Agniya in microvegetative experiments done at the All-Russian Research Institute for agriculture microbiology (ARRIAM). Selected salt tolerant plants of both varieties were planted further in greenhouse complex (STC) of V.R. Villiams All-Russian Research Institute for Forages. It was found that salt-tolerant plants of Soleustoychivaya and Agniya are characterized by predominantly purple color of flowers, by a twisted form of bean, by relatively high branching and bushy, by later transition to a period of winter rest according to 3-year vegetation trials in the STC. From seeds obtained from tested plants the I 1 plants were grown, which were studied in microvegetative experiments in ARRIAM. Plants I 1 of both varieties were analyzed by DM, by growth rate of above-ground and underground parts, by number of nodules formed in typical (salt-free) conditions and under salinity. As a result, it was found that generation I 1 plants of both varieties were homogeneous or sufficiently homogeneous according to DM data. Not inoculated generation I 1 plants of Soleustoychivaya variety successfully developed in saline conditions (the average increase of DM was 36.92 % in comparison to plants of initial variety). DM of plants I 1 of Agniya variety was, on the contrary, lower than that of the plants of the initial variety in the saline conditions, being 16.55 % less. The high level of interaction specificity of both varieties of generation I 1 plants with strains CIAM1774 or Rm2011, differing in salt tolerance, was assessed by DM. Thus, under salt stress impact the highest values of DM was obtained for Soleustoychivaya variety plants in symbiosis with strain Rm2011 characterizing by S. meliloti s
Текст научной статьи Отбор солеустойчивых растений разных видов люцерны ( Medicago L.) и анализ их морфобиологических и симбиотрофных показателей
Бобовые травы обладают свойствами, способными сделать их перспективными для восстановления деградированных почв (1, 2). Они фиксируют атмосферный азот в симбиозе с клубеньковыми бактериями, который в дальнейшем закрепляется в плодородном слое почвы (3). Площадь почв, нуждающихся в восстановлении, в том числе засоленных, переувлажненных и закисленных, постоянно увеличивается в связи с ухудшающейся природно-климатической обстановкой как в мире, так и в России (4-6). Успешность освоения таких территорий подчас зависит от правильного подбора возделываемых культур (7, 8).
Источником получения новых сортов с повышенным адаптивным потенциалом могут стать экотипы и эндемики бобовых, произрастающие в различных эколого-географических регионах (9). Род Medicago представлен видами, значительно различающимися по адаптивности и продуктивности в различных почвенно-климатических условиях. Так, люцерна изменчивая ( Medicago varia ) широко распространена в средней полосе России, но может расти на слабо- и среднезасоленных почвах в пустынной зоне При-каспия, а люцерна серповидная, или желтая, ( M. falcatа ) — на слабокислых почвах Мурманской и Архангельской областей (10, 11). Выявление растений (фенотипов) люцерны, различающихся по адаптивности (стрес-соустойчивости), — один из подходов к созданию сортов и симбиотических систем с повышенным адаптивным потенциалом.
Показано, что инокуляция люцерны штаммами клубеньковых бактерий ( Sinorhizobium meliloti ), подобранными на основе генетической селекции, способствует повышению ее продуктивности в условиях засоления (12-15). Определение солеустойчивости ризобий, растений и сформированных ими симбиозов можно успешно проводить в модельных лабораторных условиях и в микровегетационных опытах (1, 12, 16).
Задачей исследования была оценка растений ряда тетраплоидных и диплоидных видов люцерны по солеустойчивости при различных формах азотного питания с целью выявления контрастных фенотипов, получения их потомства от самоопыления и сравнения по морфологическим, биологическим и симбиотическим показателям. Представлены новые данные о специфичности сортомикробных систем с высокой симбиотической эффективностью в обычных условиях и под воздействием солевого стресса.
Методика . Сорта и экотипы люцерны ( Medicago L.) предоставлены из коллекции ВИР (Всероссийский НИИ растениеводства им. Н.И. Вавилова) и получены из Всероссийского НИИ кормов им. В.Р. Вильямса. Для инокуляции использовали Sinorhizobium meliloti : тест-штамм Rm2011, устойчивый к 550 мМ NaCl (типовая устойчивость) и штамм CIAM1774 (AK23 или А1), резистентный к 700 мМ NaCl (солеустойчивость, характерная для 10 % штаммов этого вида) (17). Штаммы культивировали на среде TY при 28 ° С (16).
В стерильных микровегетационных опытах растения выращивали в стандартных (бессолевых) условиях либо в присутствии 75 или 100 мМ NaCl (засоление) в 10-кратной повторности на протяжении 28 или 56 сут (в зависимости от задачи эксперимента), при необходимости в почву вносили КNO3 (3 мM) как источник азота в соответствии с описанием (12, 16). В качестве субстрата использовали вермикулит или 0,6 % агар с минеральной средой Красильникова-Кореняко. Продуктивность растений оценивали по величине прибавки (%) сухого вещества (СВ) относительно 674
контрольного образца. Для определения энергии прорастания (ЭП) семена помещали в чашки Петри при 28 ° С и на 3-и сут рассчитывали процентное отношение числа проросших семян к их общему числу. В вегетационном опыте использовали сосуды с 6 кг плодородной почвы (рН 6,94, содержание гумуса — 4,41 %, общего азота — 0,28 %, подвижного фосфора и калия — соответственно 560,7 и 432,0 мг/кг почвы) и выращивали без подкормки при естественной освещенности и температуре. Фотопериодичность растений анализировали при естественном освещении и контролируемых температурных условиях.
Генотипы с разной солеустойчивостью отбирали в микровегетаци-онном опыте на 56-е сут на фоне 75 мМ NaCl согласно описанию (18). Отобранные растения подращивали в сосудах объемом 0,5 л со стерильным вермикулитом в бессолевых условиях, по истечении 2 мес пересаживали в сосуды с почвой и выращивали в фитотроне MLR-351H («Sanyo Electric Co.», Япония) до получения семян методом принудительного самоопыления. Принудительное самоопыление, анализ морфологических и биохимических показателей (2012-2014 годы) выполняли по описанной методикам (19) в условиях селекционно-тепличного комплекса (СТК, Всероссийский НИИ кормов им. В.Р. Вильямса). Растения поколения I1 (сорта Солеустойчивая и Агния), выращенные из семян (по 20 шт. в каждом из 8 вариантов), изучали по однородности, продуктивности и симбиотической эффективности в микровегетационных опытах (16).
Для статистического анализа использовали программу Statistica v. 6.0 и пакет программ MS Excel 2013. Проводили дисперсионный, корреляционный анализ, вычисляли коэффициент вариации ( Сv ) и t -критерий Стьюдента (20, 21).
Результаты . Сорта и экотипы люцерны были представлены образцами из географически удаленных регионов с засушливыми условиями и(или) засоленностью, с резкими перепадами суточных и сезонных температур (табл. 1). Для исследования отобрали возделываемые сорта люцерны посевной и изменчивой ( M. sativa , M. varia ), популяции дикорастущей люцерны M. falcatа из северного и южных регионов, один из последних подвержен экстремальному засолению (Приаралье) (см. табл. 1). В том же районе произрастает эндемичный вид многолетней люцерны М. trautvetteri . Кроме того, были проанализированы популяции М. caerulea — древнего синецветкового вида, распространенного на степных и затапливаемых прибрежных территориях Прикаспия. Также оценили солеустойчивость возделываемых сортов люцерны Солеустойчивая и Агния, из которых первый создан на основе сорта Хивинская местная методом клеточных технологий на фоне солевого стресса (22), второй — при сопряженной селекции из сложногибридной популяции П211, полученной на основе отечественных и канадского сортов (Пастбищная 88 ½ Северная гибридная 69 ½ Rizoma) (23).
1. Образцы люцерны ( Medicago L.) разного эколого-географического происхождения, протестированные на солеустойчивость
|
Вид (плоидность) |
Сорт, образец люцерны |
Район сбора семян |
¹ по каталогу ВИР |
|
М. falcata (2 n ) |
subsp. Borealis Grosshm., дикорастущая |
Псковская обл., Россия |
к-25557 |
|
subsp. Romanica рrod., дикорастущая, экотип |
Восточная часть Казахстана |
к-49669 |
|
|
М. caerulea Less. ex. Ledeb. (2 n ) |
Дикорастущая, экотип |
Дагестан, Россия |
к-12821 |
|
Дикорастущая, экотип |
Гурьевская обл., Казахстан |
к-28915 |
|
|
Дикорастущая, экотип |
Азербайджан |
к-49904 |
|
|
Дикорастущая, экотип |
Ставропольский край |
к-44044 |
Продолжение таблицы 1
М. trautvetteri Sumn . (2 n ) Дикорастущая, экотип Актюбинская обл., Казахстан к-36579
М. sativa L . (4 n ) Сорт Тибетская Актюбинская обл., Казахстан к-25782
Местный возделываемый сорт Киргизия к-6376
Узбекистан к-8913
Ливия к-39107
M. sativa L. subsp. sativa (4n) Сорт Солеустойчивая Московская обл., Россия -
M. sativa L. nothosubsp . varia
(Martyn) (4 n ) Сорт Агния Московская обл., Россия -
П р и м еч а ни е. Образцы получены из коллекции ВИР (Всероссийский НИИ растениеводства им. Н.И. Вавилова, г. Санкт-Петербург); прочерк означает, что образцы предоставлены Всероссийским НИИ кормов им. В.Р. Вильямса (Московская обл.). Тестирование проводили в микровегетационных опытах.
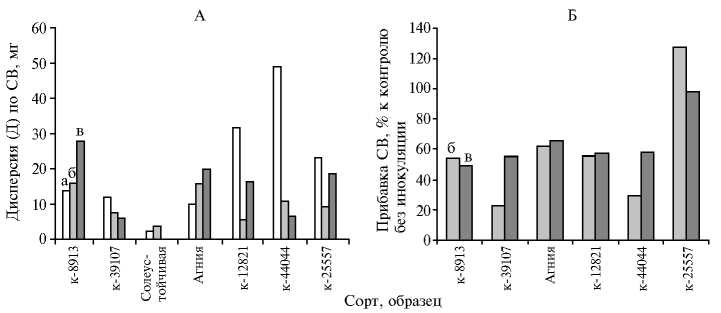
Рис. 1. Оценка дисперсии (A) и продуктивности (Б) растений люцерны ( Medicago L.) разных сортов и экотипов (по сухому веществу — СВ) при инокуляции штаммами Sinorhizobium meliloti в присутствии соли: А — 75 мМ NaCl, Б — 100 мM NaCl; а, б и в — соответственно контроль без инокуляции, инокуляция штаммами Rm2011 и CIAM1774.
Симбиотическая эффективность и солеустойчивость растений разных видов люцерны. Однородность сортов оценивали по количеству СВ у растений, которые выращивались при 75 мМ NaCl без инокуляции и при инокуляции штаммами S. meliloti , с привлечением коэффициента дисперсии (Д) в соответствии с методическими указаниями (16). Без инокуляции значения Д варьировали от 10,70 до 13,70 в случае сорта Агния и местных сортов M . sativa . Исключительно низкое (2,11) и, наоборот, высокое значение Д (17,50) выявили соответственно у сортов Солеустойчивая (рис. 1, А) и Тибетская. Зависимости между значением Д и географическим местом происхождения растений не обнаружили. Например, попарно сходные значения Д были получены для М. caerulea из районов Казахстана и Дагестана, из Азербайджана и Ставропольского края (усредненные величины Д соответственно 29,80 и 46,50). Контрастные значения Д отмечали также для популяций M. falcatа из Казахстана и Псковской области (соответственно 50,30 и 23,20). Самым высоким оказалось значение Д для растений эндемичного вида М . trautvetteri (62,90). По-видимому, сорта с низкими Д представлены генотипами со сходной устойчивостью, а при высоких Д она варьирует. Установлено, что значение Д существенно изменялось при инокуляции растений штаммами S. meliloti (см. рис. 1, А). В ряде случаев была выявлена обратная зависимость между величиной Д без инокуляции и при инокуляции (например, у М. caerulea к-12821 или сорта Агния) (см. рис. 1, А). Таким образом, изменение дисперсии отражает специфичность растительно-микробного взаимодействия.
Продуктивность растений у изучаемых видов и сортов люцерны оценивали по СВ в симбиозе со штаммами S. meliloti на фоне 100 мМ NaCl в микровегетационном опыте. Высокие прибавки по СВ отмечали у 676
местного сорта из Узбекистана (к - 8913) и сорта Агния при инокуляции соответственно Rm2011 или CIAM1774 (усредненная прибавка 57,60 %, см. рис. 1, Б). Эти же штаммы при засолении формировали эффективные симбиозы с образцами М . caerulea к - 12821 и М . falcata к - 25557. Установлено, что эффективность симбиотического взаимодействия M . sativa к - 39107 и М . caerulea к - 44044 со штаммом CIAM1774 была вдвое выше, чем при симбиозе этих же растений-хозяев со штаммом Rm2011 (см. рис. 1, Б). Сделано заключение, что местные и возделываемые сорта люцерны высокоотзывчивы на инокуляцию и могут формировать эффективные симбиозы в условиях засоления.
Исходя из полученных данных, представляло интерес выявить генотипы растений, различавшиеся по солеустойчивости. Формы, контрастные по габитусу (min — с низким стеблем, max — с развитым высоким стеблем), были отобраны из каждой изучаемой выборки. После подращивания в стерильных бессолевых условиях их переместили в фитотрон. При принудительном самоопылении семена от фенотипически различных растений были получены на 2-й год только у М . caerulea и М . falcata , а также у M . sativa L. сортов Агния и Солеустойчивая, однако только в последнем случае их количество оказалось достаточным для дальнейшего изучения.
Морфологические и биохимические показатели растений с min- и max-фенотипом. К морфологическим относят признаки, которые не зависят от условий выращивания (окраска венчика, число цветков в соцветии, форма боба), но используются для определения сортотипа люцерны. Растения сорта Солеустойчивая (Со) контрастных фенотипов (Со-max и Со-min) имели окраску венчиков цветков от сиреневой до темно-сиреневой, характерную для люцерны посевной ( M . sativa L.) синегибридного сортотипа. При этом у растений сорта Агния (Аг) контрастных фенотипов (Aг-max и Аг-min) окраска цветков различалась: сиреневая у max форм свидетельствовала о доминировании генов люцерны посевной, тогда как разнообразие оттенков от желтовато-сиреневатых до желтых у min-форм указывало на их генетическое родство с люцерной северной ( M . borealis L.), входящей в родословную одной из родительских форм исходного сорта (23).
2. Морфологические, биохимические показатели и продуктивность у фенотипически контрастных групп люцерны M. sativa L. сортов Агния и Солеустойчивая (вегетационные опыты, селекционно-тепличный комплекс, 2012-2014 годы)
|
Показатель |
Растения фенотипов |
t ф max/min |
|
|
max |
min |
||
|
Морфологические признаки: |
|||
|
оборотов в бобе, шт. |
2,7 |
2,5 |
- |
|
цветков в соцветии, шт. |
25 |
18 |
- |
|
стеблей, шт/раст. |
71 |
48 |
1,42 |
|
ветвей, шт/раст. |
253 |
124 |
2,24 |
|
стеблей с цветами, шт/раст. |
12 |
6 |
1,84 |
|
Содержание в сухом веществе, %: |
|||
|
сырой золы |
9,21 |
8,72 |
0,94 |
|
фосфора |
0,39 |
0,39 |
- |
|
калия |
1,41 |
1,39 |
0,06 |
|
Сухое вещество, г/растение |
151 |
134 |
0,88 |
|
Семена, г/растение |
12,70 |
11,50 |
0,21 |
П р и м еч а ни е. Растения max и min фенотипов — контрастные по габитусу (min — с низким стеблем, max — с развитым высоким стеблем); t ф — фактическое значение критерия Стьюдента, доверительный интервал 95 % (теоретическое значение критерия Стьюдента t 05 = 2,15). Прочерки означают, что нет достоверных различий между растениями max и min фенотипов.
У многолетних видов люцерны боб имеет форму, закрученную в спираль, число оборотов которой видоспецифично (например, 0,5-1,0 — у люцерны серповидной, 3,5-4,0 — у посевной). У сорта Агния этот показатель составил 2,1, что на один оборот меньше, чем у сорта Солеустойчивая (tф = 2,46 > t05). При этом по указанному признаку существенных различий между выборками растений с неодинаковым фенотипом мы не выявили (табл. 2), тем не менее, у имеющих солеустойчивый (max) фенотип наблюдалась тенденция к повышенной скрученности боба.
Выборки растений двух этих сортов существенно различались между собой по числу цветков в соцветии и кустистости ( t ф > t 01), однако при этом max и min выборки статистически не различались ( t ф < t 05). Тем не менее, у солеустойчивых растений прослеживалась тенденция к увеличению среднего числа цветков в соцветии и стеблей на растение (см. табл. 2).
Анализ среднего числа ветвей на растение показал, что при max фенотипе оно вдвое больше ( t ф = 2,24 > t 05, см. табл. 2). Сходные результаты получили при анализе числа стеблей с цветами на растение ( t ф = 1,84; t 05 = 2,15; см. табл. 2). Следовательно, растения солеустойчивого фенотипа имели тенденцию к скороспелости.
Анализ фотопериодичности при естественном освещении и контролируемых температурных условиях показал, что сокращение светового дня (10 ч и менее, III декада октября в Центральном районе РФ) приводит к увяданию и переходу растений к стадии покоя. Этот процесс количественно оценивают по содержанию СВ. Установлено, что у растений с max-фенотипом этот показатель в период увядания был на 11,30 % выше (по данным на 20 октября 2014 года, см. табл. 2). При выходе из зимнего покоя и увеличении продолжительности светового дня до 8 ч у max-форм отмечали раннее отрастание, а при увеличении светлого времени суток еще на 1 ч — отрастание у растений min-фенотипов ( t ф max = 2,36; t ф min = 1,97; t 05 = 2,15). Изученные выборки различались по содержанию сырой золы (см. табл. 2). Таким образом, приведенные данные свидетельствуют в пользу того, что солеустойчивые растения дольше сохраняли активность при укороченном световом периоде.
Поскольку растения контрастных фенотипов выращивали на плодородной почве с нейтральным значением кислотности (рН 6,94), то это не позволило выявить четкие различия между изучаемыми группами. Тем не менее, обобщая полученные данные, можно заключить, что для растений с солеустойчивым фенотипом, выявленным у изучаемых сортов, характерна преимущественно сиреневая окраска цветов, более закрученная форма боба, сравнительно высокая ветвистость и кустистость, фотопериодическая реакция короткого дня и повышенная зольность СВ.
Однородность и продуктивность в поколении I1. Растения I1, которые вырастили из семян, полученных от контрастных фенотипов методом принудительного самоопыления в условиях СТК, изучали по основным характеристикам в условиях микровегетационных опытов (в каждом из 8 вариантов по 20 шт.).
Оценка однородности семян I1 обоих сортов по ЭП относительно семян исходных форм (контроль) не выявила существенных различий (усредненная величина ЭП 95 %). Растения I1 без инокуляции оценивали по СВ с привлечением коэффициента варьирования ( Сv ). Для растений обоих изученных сортов люцерны — Солеустойчивая (Со-I1) и Агния (Аг-I1) значения Сv были существенно ниже таковых в соответствующих контро-лях (рис. 2, А). Однако для отдельных фенотипических групп значение Сv существенно изменялось в зависимости от условий выращивания. Например, для Аг-I1-max2 этот показатель был выше без засоления, а для Со-I1-678
min2 — при засолении (см. рис. 2, А). Согласно полученным нами данным, фенотипически несходные растения Со-I1 и Аг-I1 следует рассматривать как однородные ( Cv < 17 %) (21), а две вышеупомянутые группы — как достаточно однородные ( Cv в пределах 17-33 %) (21).
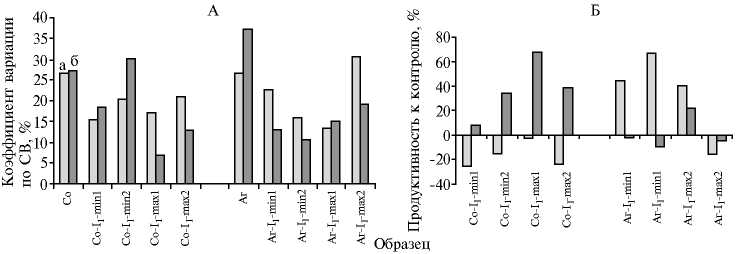
Рис. 2. Оценка однородности по Сv (А) и продуктивности (Б) растений I 1 из фенотипически контрастных групп люцерны M. sativa L. сортов Агния (Аг) и Солеустойчивая (Со) по сухому веществу (СВ) в норме и при засолении: а — контроль (без засоления), б — в присутствии 75 мМ NaCl; min и max — формы соответственно с низким и развитым высоким стеблем. При выращивании образцов не использовали инокуляцию Sinorhizobium meliloti и минеральный азот (микровегетационные опыты).
Продуктивность контрастных фенотипов растений Аг-I1 и Со-I1 по СВ (без инокуляции) в отсутствие засоления и при засолении сравнивали с показателями исходных растений соответствующих сортов (Аг и Со), полученными в аналогичных условиях (контроль, см. рис. 2, Б). Без добавления NaCl min- и max-растения Аг-I1 значительно различались по продуктивности. Так, генотипы Аг-I1 min- и max-растений либо существенно (в среднем на 50,50 %) превосходили по СВ контрольные растения, либо достоверно не отличались от них. В условиях засоления растения только одной (Аг-I1-maх1) из четырех фенотипических групп развивались лучше контрольных (превышение по СВ на 22,10 %) (см. рис. 2, Б). Растения Со-I1, наоборот, успешно развивались в условиях засоления (прибавка СВ от 33,85 до 67,69 %), за исключением группы Со-I1-min1, у которой значение прибавки СВ относительно контроля не превысило 7,69 %. Продуктивность всех фенотипических групп Со-I1 в отсутствие засоления была в среднем на 16,55 % ниже контроля. Внесение минерального азота в субстрат способствовало увеличению показателя СВ во всех фенотипических группах Со-I1 и Аг-I1 в среднем в 1,90 раза независимо от наличия или отсутствия засоления (данные не представлены).
Таким образом, фенотипически несходные группы растений Аг-I1 и Со-I1, охарактеризованные как однородные, тем не менее, существенно различались по продуктивности. Группы Со-I1 оказались более выраженными галофитами, чем растения исходного сорта. Наблюдаемый факт обусловлен тем, что сорт Солеустойчивая, полученный в результате культивирования каллуса на среде с 1,98 М NaCl (5), имеет расщепление по фенотипу. Растения Аг-I1 не отличались по солеустойчивости от контроля, однако показано, что и сорт Агния может быть использован для поиска стрессоустойчивых генотипов. Следовательно, дальнейший анализ будет проводиться на фенотипически различных группах Аг-I1 и Со-I1.
Симбиотические системы на основе растений I1. Эффективность симбиоза растений Аг-I1 со штаммом CIAM1774 или Rm2011 была сходной при засолении и на субстрате без добавления NaCl (усредненное значение по прибавкам СВ соответственно 60,05 и 135,05 %) (рис. 3, А). Они также мало различались в случае симбиоза растений Со-I1 с 679
теми же штаммами в варианте без засоления (в среднем 43,80 %), однако на фоне засоления прибавка в варианте с Rm2011 оказалась на 23,0 % больше, чем с CIAM1774 (см. рис. 3, Б). Таким образом, при инокуляции у растений Аг-I1 происходило увеличение сухого вещества в 3 раза в обычных условиях, у Со-I1 — в 2 раза при солевом стрессе.
Следовательно, в условиях солевого стресса высокоэффективен симбиоз солеустойчивого сорта люцерны со штаммом, имеющим типовую степень солеустойчивости (Rm2011). Вместе с тем, следует отметить перспективность симбиоза, сформированного на основе полученного солеустойчивого генотипа сорта Агния с солеустойчивым штаммом CIAM1774.
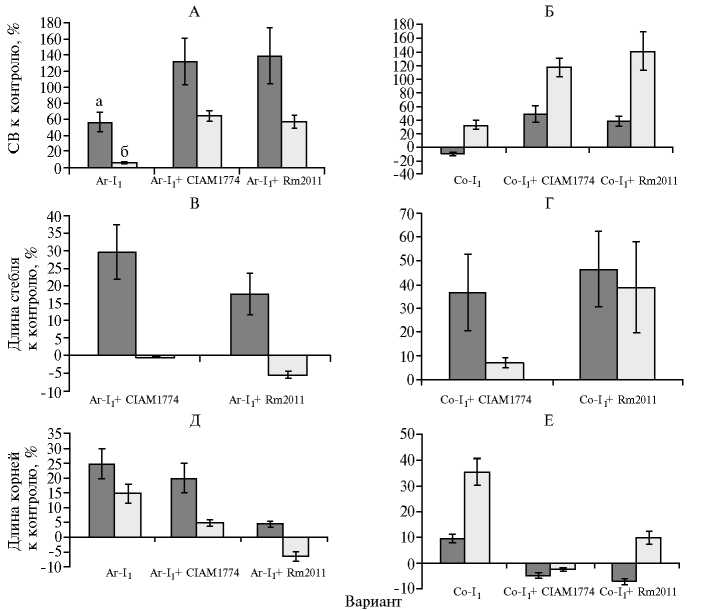
Рис. 3. Изменение содержания сухого вещества (СВ) (А, Б) , длины стеблей (В, Г) и корней (Д, Е) у растений I 1 люцерны M. sativa L. сортов Агния (А, В, Д) и Солеустойчивая (Б, Г, Е) при симбиотрофном питании в норме (а) и в условиях солевого стресса (б) : Аг и Со — соответственно сорта Агния и Солеустойчивая; CIAM1774 и Rm2011 — штаммы Sinorhizo-bium meliloti , использованные для инокуляции; А, Б, Д, Е — относительно соответствующих неинокулированных или инокулированных растений Аг или Co, В, Г — относительно не-инокулированных растений Ar-I 1 или Co-I 1 . Содержание NaCl в субстрате при солевом стрессе — 75 мМ (микровегетационные опыты).
Оказалось, что на субстрате без добавления NaCl в группе Аг-I1 стебель развивался лучше в случае инокуляции растений штаммом CIAM1774 (удлинение на 12,04 % по сравнению с вариантом, когда использовали штамм Rm1021, и на 29,62 % — по отношению к контролю без инокуляции) (см. рис. 3, В, Г). Наоборот, у растений Со-I1 длина стебля была на 9,78 % больше в варианте с Rm2011, чем с CIAM1774. Однако в условиях солевого стресса длина стебля растений Аг-I1 при симбиотрофном питании оказалась близкой или незначительно меньше (на 5,48 %) контроля. В то же время у растений Ср-I1 при солевом стрессе длина стебля увеличивалась в симбиозе с Rm2011 на 38,85 %, с CIAM1774 — только на 7,05 % (см. рис. 3, В, Г).
Таким образом, в отсутствие солевого стресса высота надземной части растений Аг-I1 и Со-I1 зависела от штамма-инокулянта, тогда как по СВ достоверных различий не выявили (см. рис. 3, В, Г). При этом растения Аг-I1 и Со-I1 при симбиозе соответственно с Rm2011 или CIAM1774 были низкорослыми. Аналогичные различия в условиях засоления отмечали для Со-I1: при инокуляции CIAM1774 растения были низкорослыми, Rm2011 — высокорослыми, при том что соответствующие значения СВ достоверно не различались.
Интенсивное развитие стебля и накопление биомассы растений зависит от процесса азотфиксации, обусловленного сформированными на корнях клубеньками (10). У Аг-I1 и Со-I1 число клубеньков в отсутствие солевого стресса при инокуляции CIAM1774 было в 2 раза больше, чем в варианте с Rm2011, относительно контрольных растений, инокулированных тем же штаммом (соответственно на 29,11 и 10,27 %). В условиях засоления этот показатель у растений Со-I1 и Аг-I1 был соответственно в 3,02 и в 1,34 раза выше при инокуляции Rm2011, чем в случае CIAM1774. Полученные данные указывают на то, что растительно-микробные взаимодействия могут существенно различаться в обычных условиях и при засолении.
Также оценивали развитие корневой системы, которая служит важной характеристикой успешности развития растений (24). Длина корня у неинокулированных растений I1 обоих сортов в обычных условиях была достоверно больше соответствующих значений у контрольных растений (см. рис. 3, Д, Е). Например, у растений Аг-I1 этот показатель превышал таковой у Со-I1 в 2,57 раза, тогда как при засолении у Со-I1 отмечали удлинение корня на 35,48 %, что более чем в 2 раза превысило соответствующее значение для Аг-I1 (см. рис. 3, Д, Е). Следовательно, у люцерны развитие корневой системы в условиях солевого стресса может служить характеристикой сорта.
При инокуляции растений I1 длина корня, как оказалось, существенно изменялась. Так, сходное уменьшение этого показателя у Аг-I1 и Со-I1 отмечали в симбиозе с Rm2011 (в среднем на 20,62 %) как в обычных условиях, так и на фоне засоления. При инокуляции растений Аг-I1 штаммом CIAM1774 уменьшение длины корня было недостоверным, если NaCl в субстрат не добавляли, и составило 10,43 % в условиях солевого стресса относительно соответствующих контрольных растений. Инокуляция растений Со-I1 тем же штаммом привела к сокращению длины корня (относительно контролей) на 14,44 и 37,88 % соответственно в обычных условиях и при засолении. Следовательно, солеустойчивый штамм CIAM1774 специфически влиял на длину корня у растений Со-I1, что выражалось в уменьшении этого показателя относительно такового у Аг-I1 в 2,87 и 3,76 раза соответственно в обычных условиях и при солевом стрессе. Таким образом, установлен факт влияния штамма-инокулянта на развитие корневой системы растения-хозяина, однако изменение длины корня не коррелировало с продуктивностью растений как в норме, так и при засолении (соответственно r = 0,3 и r = 0,5).
В заключение отметим, что в результате отбора и анализа контрастно различавшихся по солеустойчивости групп растений были получены принципиально новые данные об уровне специфичности сортомикробных систем с высокой симбиотической эффективностью в стандартных условиях и под воздействием солевого стресса. Для симбиотических систем, полученные на основе солеустойчивых растений поколения I1, характерна повышенная продуктивность, активное клубенькообразование, более длинный стебель растений. Впервые показано, что при симбиотрофном питании имеет место уменьшение длины корневой системы, что может указы- вать как на увеличение поглотительной способности корней (Г.В. Степанова, персональное сообщение), так и на активацию процессов передачи азота из корней в надземную часть (25). Установлено, что этот биометрический показатель зависит от специфичности растительно-микробного взаимодействия. Интересно, что штамм солеустойчивого фенотипа оказывал более существенное влияние на развитие подземной части растения именно у солеустойчивого фенотипа, что особенно проявилось в условиях солевого стресса. Подобный факт может быть обусловлен тем, что при взаимодействии микро- и макросимбионта, обладающих повышенной соле-устойчивостью, формируются оптимальные условия для метаболической активности бактероидов, внутренняя среда которых (даже в стандартных условиях) является гиперосмотической (26). Выявленные нами сочетания растения-хозяина и штамма-инокулянта в условиях солевого стресса указывают на то, что в описанных симбиосистемах создается и поддерживается оптимальный баланс азота и углерода. Очевидно, что изучение метаболических особенностей симбиосистем, сформированных на основе стрессоустойчивых микро- и макросимбионтов является существенным этапом на пути направленного конструирования симбиотических систем с заданным адаптивным потенциалом.
Итак, проведенное нами исследование показало перспективность анализа стрессоустойчивости как растительного, так и микробного компонентов, что обусловливает возможность направленного конструирования симбиотических систем с заданным адаптивным потенциалом.
Список литературы Отбор солеустойчивых растений разных видов люцерны ( Medicago L.) и анализ их морфобиологических и симбиотрофных показателей
- Zahran H.H. Rhizobium-legume symbiosis and nitrogen fixation under severe conditions and in an arid climate. Microbiology Molecular Biology Review, 1999, 63(4): 968-989 ( ) DOI: 10.1007/BF00369391
- Писковацкий Ю.М., Ломова М.Г., Соложенцева Л.Ф., Ломов М.В. Селекция люцерны для многовидовых кормовых агрофитоценозов. В сб.: Многофункциональное адаптивное кормопроизводство. Вып. 5(53). М., 2015: 159-168.
- Умаров М.М., Кураков А.В., Степанов А.Л. Микробиологическая трансформация азота в почве. М., 2007 (ISBN 5-89118-315-7).
- Добровольский Г.В. Почвенные ресурсы Pоссии за 150 лет. Россия в окружающем мире (aналитический ежегодник). М., 2002: 1-16.
- Reynolds J.F, Smith D.M.S., Lambin E.F., Turner B.L., Mortimore M., Batterbury S.P.J., Walker B. Global desertification: building a Science for dryland development. Science, 2007, 316: 847-851 ( ) DOI: 10.1126/science.1131634
- Rattan L. Restoring soil quality to mitigate soil degradation. Sustainability, 2015, 7: 5875-5895 ( ) DOI: 10.3390/su7055875
- Swaraj K., Bishnoi N.R. Effect of salt stress on nodulation and nitrogen fixation in legumes. Indian J. Exp. Biol., 1999, 37(9): 843-848.
- Косолапов В.М., Костенко С.И., Пилипко С.В. Адаптивные сорта кормовых трав для экстремальных условий России. Достижения науки и техники АПК, 2013, 7: 71-73.
- Дзюбенко Н.И., Чапурин В.Ф., Бухтеева А.В., Сосков Ю.Д. Мобилизация и изучение многолетних кормовых культур в свете идей Н.И. Вавилова. Труды по прикладной ботанике, генетике и селекции (ВИР, СПб), 2007, 164: 153-163.
- Гасанов Г.Н., Усманов Р.З., Мусаев М.Р., Абасов М.М. Экологическое состояние и возможности фитомелиорации засоленных почв Западного Прикаспия. Юг России: экология, развитие, 2007, 1: 79-85.
- Методика эффективного освоения многовариантных технологий улучшения сенокосов и пастбищ в Северном природно-экономическом районе. М., 2015.
- Ибрагимова М.В., Румянцева М.Л., Онищук О.П., Белова В.В., Курчак О.Н., Андронов Е.Е., Дзюбенко Н.И., Симаров Б.В. Симбиоз клубеньковых бактерий Sinorhizobium meliloti с люцерной Medicago sativa в условиях засоления. Микробиология, 2006, 75(1): 94-100 ( ) DOI: 10.1134/S0026261706010140
- Tikhonovich I.A., Provorov N.A. Cooperation of plants and microorganisms: Getting closer to the genetic construction of sustainable agro-systems. Biotechnology Journal, 2007, 2(7): 833-848 ( ) DOI: 10.1002/biot.200700014
- Степанова Г.В. Селекция люцерны на повышение эффективности симбиотических взаимодействий. Т. 6. Уфа, 2008: 32-38.
- Gubry-Rangin C., Garcia M., Bena G. Partner choice in Medicago truncatula-Sinorhizobium symbiosis. Proceedings of Biological Science, 2010, 277(1690): 1947-1951 ( ) DOI: 10.1098/rspb.2009.2072
- Румянцева М.Л., Симаров Б.В., Онищук О.П., Андронов Е.Е., Чижевская Е.П., Белова В.С., Курчак О.Н. Биологическое разнообразие клубеньковых бактерий в экосистемах и агроценозах: теоретические основы и методы. СПб-Пушкин, 2011.
- Румянцева М.Л., Мунтян В.С. Клубеньковые бактерии Sinorhizobium meliloti: солеустойчивость и ее генетическая детерминированность. Микробиология, 2015, 84(3): 263-280 ( ) DOI: 10.7868/S0026365615030179
- Курчак О.Н., Проворов Н.А., Онищук О.П., Воробьев Н.И., Румянцева М.Л., Симаров Б.В. Воздействие солевого стресса на генетически полиморфную систему Sinorhizobium meliloti-Medicago truncatula. Генетика, 2014, 50(7): 777-786 ( ) DOI: 10.7868/S001667581406006X
- Методические указания по селекции многолетних трав. М., 1985.
- Лакин Г.Ф. Биометрия. М., 1980.
- Трухачева Н.В. Математическая статистика в медико-биологических исследованиях с применением пакета Statistica. М., 2012.
- Каталог районированных и перспективных сортов кормовых культур селекции Всероссийского научно-исследовательского института кормов имени В.Р. Вильямса. М., 2007.
- Степанова Г.В., Золотарев В.Н., Мунтян В.С., Румянцева М.Л. Отзывчивость нового сорта люцерны Агния на инокуляцию клубеньковыми бактериями. Адаптивное кормопроизводство, 2013, 3(15): 43-48 (http://www.adaptagro.ru).
- Тищенко Е.Д., Тищенко А.В., Нижеголенко В.М. О корреляции признаков у люцерны. Кормопроизводство, 2012, Ноябрь: 21-22.
- Онищук О.П., Воробьев Н.И., Проворов Н.А., Симаров Б.В. Симбиотическая активность ризобий люцерны (Sinorhizobium meliloti) с генетическими модификациями системы транспорта дикарбоновых кислот. Экологическая генетика, 2009, 7(2): 3-10.
- Mandon K., Osteras M., Boncompagni E., Trinchant J.C., Spennato G., Poggi M.C., Le Rudulier D. The Sinorhizobium meliloti glycine betaine biosynthetic genes (betlCBA) are induced by choline and highly expressed in bacteroids. Mol. Plant Microbe Interact., 2003, 16(8): 709-719 ( ) DOI: 10.1094/MPMI.2003.16.8.709
