Отношение растений видов и сортов гречихи к основным абиотическим факторам

Автор: Наполова Г.В., Наполов В.В.
Журнал: Вестник аграрной науки @vestnikogau
Рубрика: Физиология растений
Статья в выпуске: 3 (6), 2007 года.
Бесплатный доступ
В статье отражены результаты исследований биологических особенностей растений гречихи различных экологических групп. Проанализированы способы и направленность продукционных процессов при стрессовых факторах. Изучена терморезистентность растений гречихи, определена репарационная способность тканей, органов и растения в целом.
Короткий адрес: https://sciup.org/147123240
IDR: 147123240 | УДК: 633.12:581.4
Текст научной статьи Отношение растений видов и сортов гречихи к основным абиотическим факторам
Для обеспечения применения адекватной схемы обработок фунгицидами в хозяйствах должны быть разработаны и запланированы различные схемы проведения опрыскиваний : стандартная для выпол нения технологических работ в соответствии с ка лендарным планом и интенсивная , с максимальной интенсификацией выполнения приемов для предот вращения развития массовых вспышек и снижения потерь урожая и его качества .
Для повышения устойчивости агроценоза ози мой пшеницы в интенсивных технологиях и дости жения высокой урожайности и качества зерна необ ходимо последовательно использовать при вегета ции фунгициды альто супер 0,5 л / га в фазу выхода в трубку и амистар экстра 0,7 л / га в фазу колошение .
При обработках против комплекса болезней , если их нарастание идет с различной интенсивностью , ори ентироваться следует на болезнь , развивающуюся наиболее быстро . Обработку против болезней надо связывать с динамикой их развития , а не со стадией развития . Если в посеве отмечены первые признаки болезни , а погодные условия не способствуют ее раз витию , то применение фунгицида нецелесообразно .
Литература
-
1. Алехин В . Т . Пути стабилизации фитосани - тарной обстановки .// Защита и карантин растений .-2004.- № 1.- С .8-12.
-
2. Буров В . Н ., Долженко В . И ., Сухорученко Г . И ., Тютерев С . Л . Состояние , проблемы и перспек тивы химического метода защиты растений на поро ге ХХ I века . Вестник защиты растений .- С . Пб .: ВИЗР , 2001.- С . 3-15.
-
3. Ж ученко А . А . Ресурсный потенциал производ ства зерна в России .- М .: Изд - во Агрорус , 2004.-1110 с .
-
4. Захаренко В . А ., Кузмичев А . А ., Плотников В . Ф . и др . Основные мероприятия по борьбе с бо лезнями растений .// Защита и карантин растений .-2003- № 12.- С .16-25.
-
5. Концепция научного обеспечения развития агропромышленного комплекса Российской Феде рации на период до 2010 г ./ Огарков А . П ., Соколов М . Н ..- М .: МСХ РФ , РАСХН , 2003.- 33 с .
-
6. Котова В . В ., Котикова Г . Ш ., Гришечкина Л . Д . Комплекс мероприятий по защите растений от болезней .- С .- Пб .: ВИЗР , 2004.-32 с .
-
7. Санин С . С . Основные составляющие звенья систем защиты растений от болезней .// Защита и ка рантин растений .- 2003.- № 10.- С . 16-21.
-
8. Санин С . С . Влияние вредных организмов на качество зерна .// Защита и карантин растений .-2004.- № 11.- С .14-18.
УДК : 633.12:581.4
Отношение растений видов и сортов гречихи К ОСНОВНЫМ АБИОТИЧЕСКИМ ФАКТОРАМ
Г.В. Наполова , к.б.н. (Орел ГАУ)
В.В. Наполов , к.с.- х .н. (Орел ГАУ)
Центром происхождения многих видов гречихи являются субтропики Китая или Индии . Эти районы характеризуются теплым климатом и большим коли чеством осадков , наибольшее количество которых приходится на вторую половину вегетации гречихи . Эти обстоятельства наложили отпечаток на жизнен ный цикл , биологию и морфологию различных видов гречихи . Причем , эти особенности остаются мало из менившимися в течение многих веков , особенно у ди корастущих видов . У видов , подвергшихся окультури ванию - гречиха культурная (F. esculentum Moench) и татарская (F. tataricum Gaertn), изменения происходили главным образом под влиянием двух факторов - про изводственной деятельности человека и искусствен ных отборов ( бессознательного и сознательного ).
Возделывание гречихи связано с началом изуче ния её агротехники и установление условий произра стания , а также отбор форм отвечающих хозяйствен ным требованиям . Повышение плодородия и рыхлости почвы , улучшение обеспеченности влагой и питатель ными веществами сказались на увеличение изменчи вости и усилении некоторых хозяйственно - ценных признаков . Сознательный отбор и системная селекци онная работа с применением специальных методов в значительной мере ускорили формирование этих из менений . Вместе с тем , на выбор направлений и мето дов селекции большое влияние оказывал биоклимати - ческий потенциал страны или региона , в которых про водилась селекционная работа . Учитывались , прежде всего , его лимитирующие факторы . За последние два десятилетия селекционерами создано много новых сортов гречихи . Они различаются морфологией , про должительностью вегетации , соотношением периодов вегетативного и генеративного развития , семенной продуктивностью и рядом других особенностей . Од новременно осуществляется работа по дальнейшему изменению отдельных морфологических признаков и биологических свойств , повышающих адаптивность и продуктивность сорта на фоне общих защитно приспособительных реакций , свойственных виду .
Из всего многообразия биологических особен ностей видов , подвидов и сортов гречихи наиболь ший интерес представляет их отношение к темпера туре , влаге , почве , параметрам вегетационного пе риода . Из всего спектра биологических особенно стей , выше перечисленные являются наиболее важ ными при обосновании на физиологическом уровне продукционного процесса , - базы для формирова ния биологической и семенной продуктивности .
Объекты и методы исследования
Объектами исследования служили виды и мор-фогенотипы гречихи, ретроспективно отражавшие становление вида гречихи культурной от диких форм до современных высокопродуктивных ее сортов: Fagopyrum cymosum ( 4n), Китай; Fagopyrum tataricum ssp.potanini, Китай; Fagopyrum tataricum ssp.tataricum, Китай; Fagopyrum homotropicum (2n), Китай; Fagopyrum homotropicum (4n) Китай; Fagopyrum esculentum ssp. ancestrale, Китай; Fagopyrum esculentum ssp. esculentum: Богатырь, Россия; Шатиловская 5, Россия; Краснострелецкая, Россия; Казанская 3, Россия; Молва, Россия; Минчанка (4n), Беларусь; М- 822, Северный Китай; Botansoba, Япония; Shinano-1, Япония; Shinanonatsusoba, Япония; Скороспелая 81, Россия; Киевская, Украина; Астра, Украина; Коричневая Лимадо, Ю жный Китай; Mankan Royal, Канада.
Физиолого - биохимические анализы проводи лись на оборудовании экспериментальной базы ла боратории физиологии растений с использованием тепличного комплекса .
В наших исследованиях терморезистентность рас тений гречихи изучалась методом проращивания се мян при прямом воздействии на них различными тем пературами с использованием климатических камер .
Изучение водопотребления растений проводилось в процессе их жизнедеятельности , как при оптимальной , так и недостаточной влажности почвы (70% и 30% от полной влагоемкости соответственно ), путем ежеднев ного взвешивания сосудов с ними за вычетом воды , испаряющейся с поверхности почвы . Так же различия по отношению к водному фактору устанавливалось в опытах с созданием условий физиологической сухости с помощью гипертонических растворов осмотиков ( саха роза , маннит , полиэтиленгликоль и др .).
Особенности продукционного процесса изучались по накоплению сырой и сухой биомассы отдельных органов ( стебли , ветви , листья , плоды , корни ) весовым методом по основным фазам развития . Абсолютно су хое вещество определяли по А . В . Петербургскому .
При этом определялась концентрация азота , фосфора , калия в тканях отдельных органов :
-
- изменение концентрации азота в тканях органов микрометодом Кьельдаля в модификации Сереньева ;
-
- концентрация фосфора в тканях по методу А . Молюгина и С . Хреновой с помощью ФЭК – КФК -2;
-
- концентрация калия в растворах на пламенном фотометре FLAPHO 40.
Результаты и их обсуждение :
Гречиха произрастает в различных регионах мира с широким диапазоном температурных условий . Так , в Западном Тибете ( Индия , Непал ) и Восточном ( Китай ), где отмечено большое разнообразие ее видов и подвидов , сумма среднесуточных температур воздуха выше +10 ° С составляет 5500.. .7000 ° С , в Японии - от 2000 до 4000 ° С и в Канаде - 1500^2000 ° С . В России температурный диа пазон зон ее возделывания также весьма широк - от 1500...2000 ° С на Северо - западе Европейской части и в Западной Сибири до 3000.. .3500 ° С на Северном Кавказе .
Как установлено в ходе исследований, изучаемые виды и сорта гречихи существенно различались по способности их семян прорастать при пониженной температуре воздуха (рис. 1, табл. 1). При этом у сортов возделываемых видов и подвидов (F. escul. ssp. esculentum, F. tat. ssp. tataricum) этот процесс начинался при более низкой (+5...+6°С) температуре, и в течение двух недель они прорастали на 70...80%. В то же время семена большинства диких видов и подвидов начинали прорастать при температурах выше + 10°С и прорастали только на 3...15%.
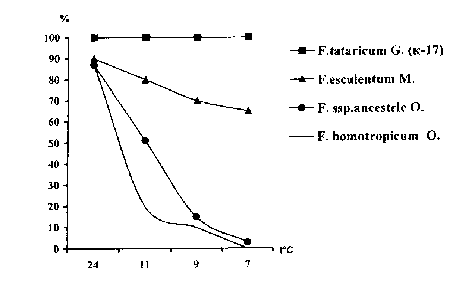
Рисунок 1 - Динамика прорастания семян видов и под видов гречихи при различной температуре воздуха
Наибольшей дивергенцией по способности се мян прорастать при пониженной температуре отли чались генотипы гречихи культурной . Абсолютное количество проросших семян в большинстве случа ев соответствовало эколого - географическому про исхождению и возделыванию сорта .
Таблица 1 - Способность семян видов , подвидов и сортов гречихи к прорастанию при пониженной температуре воздуха (+5...+6 ° С )
|
№ п / п |
Вид , подвид , сорт |
Проросло семян , % |
|
1 |
F.cymosum ( 4n) |
0,0 |
|
2 |
F.tat.ssp.potanini |
15,0 |
|
3 |
F.tat.ssp.tataricum |
96,3 |
|
4 |
F.homotropicum (2n) |
6,7 |
|
5 |
F.homotropicum (4n) |
3,5 |
|
6 |
F.escul.ssp. ancestrale |
12,1 |
|
7 |
F.escul.ssp. esculentum: |
71,0 |
|
8 |
Богатырь ( Россия ) |
73,4 |
|
9 |
Шатиловская 5 ( Россия ) |
75,1 |
|
10 |
Краснострелецкая ( Россия ) |
71,4 |
|
11 |
Казанская 3 ( Россия ) |
82,6 |
|
12 |
Молва ( Россия ) |
70,9 |
|
13 |
Скороспелая 81 ( Россия ) |
54,4 |
|
14 |
Минчанка ( Белорусь ) |
71,0 |
|
15 |
Киевская ( Украина ) |
32,2 |
|
16 |
Астра ( Украина ) |
18,6 |
|
17 |
М - 822 ( Северный Китай ) |
89,4 |
|
18 |
Коричневая Лимадо ( Ю жный Китай ) |
36,7 |
|
19 |
Botansoba ( Япония ) |
85,3 |
|
20 |
Shinano-1( Япония ) |
61,4 |
|
21 |
Shinanonatsusoba ( Япония ) |
65,9 |
|
22 |
Mankan Royal ( Канада ) |
46,8 |
Отношение растений к температурному фактору в период активной вегетации имеет значительно более сложный характер , чем в стадии прорастания семян .
Это связано с различным физиолого биохимическим состоянием растений .
По исследованиям Дроздова С . Н ., Курец В . К ., Титова А . Ф .,1984. Выделяют 5 температурных зон , влияющих на терморезистентность растений . Это фоновая , 2 закаливающие и 2 повреждающие в об ласти повышенных и пониженных температурных значений соответственно .
Терморезистентность растений гречихи - сложный в физиологическом отношении процесс , заключающий ся в устойчивость тканей различных органов к измене нию температуры за фоновым диапазоном и их репара ционной способности , а так же растения в целом .
В условиях Центральной России для большинства однолетних диких видов гречихи , происходящих из Китая , для прохождения этапа вегетативного развития требовалась сумма активных температур от 240 до 331 ° С , а генеративного - более 700 ° С . Так как не все из них успевали полностью закончить жизненный цикл , то общая потребность в сумме активных температур мо жет быть ориентировочно определена свыше 1000 ° С .
Наиболее наглядно проявляется взаимосвязь по требности в тепле с биоклиматическим потенциалом региона возделывания и направлений селекции у гено типов гречихи культурной . Селекция ее на скороспе лость , характерная для большинства регионов России в связи с коротким безморозным периодом вегетации предопределила и меньшую потребность сортов в сум ме активных температур в пределах 750.. .850 ° С .
Таблица 2 - Потребность видов , подвидов и сортов гречихи в сумме активных температур (> +10 ° С ) в отдельные этапы роста и развития растений , о C
|
Вид , подвид , происхождение |
Периоды вегетации |
||
|
посев - начало цветения |
цветение - созревание |
за всю вегетацию |
|
|
F.cymosum (4n), Китай |
518 |
670 |
1188 |
|
F.tat.ssp.potanini, Китай |
331 |
707 |
1038 |
|
F.tat.ssp.tataricum, Китай |
320 |
579 |
900 |
|
F.homotropicum (2n), Китай |
315 |
728 |
1043 |
|
F.homotropicum (4n), Китай |
241 |
753 |
994 |
|
F.escul.ssp.ancestrale, Китай |
262 |
737 |
999 |
|
F.escul.ssp.esculentum : |
205-313 |
505-757 |
760-1023 |
|
Богатырь (2n), Россия |
205 |
555 |
760 |
|
Молва , Россия |
215 |
604 |
819 |
|
Минчанка (4n), Беларусь |
243 |
648 |
889 |
|
М - 822, Северный Китай |
227 |
757 |
984 |
|
Botansoba, Япония |
275 |
678 |
953 |
|
Shinano-1, Япония |
313 |
710 |
1023 |
|
Shinanonatsusoba, Япония |
269 |
636 |
905 |
|
Mankan Royal, Канада |
262 |
695 |
957 |
Сорта , происходящие из Японии , Канады , Се верного Китая для прохождения этапа вегетативного развития требовали в условиях Центральной России суммы активных температур от 250 до 300 ° С , а ге неративного - от 636 до 757 ° С , что на 150 ° С больше , чем у сортов возделываемых в России .
Большинство исследователей относят гречиху к числу влаголюбивых культур , что в принципе соответ ствует представлениям о первичных центрах ее проис хождения и распространения ( субтропики Китая и Индии с теплым и влажным климатом ), о филогенезе рода и его видов [4, 6]. Утвердилось мнение , что на образование единицы сухого вещества надземной мас сы у сортов гречихи культурной расходуется от 500 до
600 единиц воды , что значительно больше , чем у дру гих зерновых культур [1]. Вегетационные опыты по выращиванию различных видов и сортов гречихи в условиях Центральной России в целом подтверждают эти данные , но и вносят существенные коррективы в абсолютные их величины .
Водопотребление растений и расход воды на обра зование единицы сухого вещества генетически детер минировано через продолжительность вегетационного периода , облиственность , масштабы продукционного процесса , развитие корневой системы и механизмов регулирования транспирации . С другой стороны , вы явлена высокая фенотипическая зависимость этих по казателей от условий их произрастания ( наличие влаги в почве , температура и относительная влажность воз духа , агротехника возделывания ). По нашим данным , в условиях Центральной России наибольшее количество воды за вегетационный период потребляли представи тели диких видов и подвидов гречихи , интродуциро ванные из Китая ( табл . 3).
Таблица 3 - Потребление и расход воды на образование единицы сухого вещества растениями различных видов и сортов гречихи
|
Вид , подвид , сорт |
Периоды роста и развития |
|||||
|
вегетативный |
генеративный |
за всю вегетацию |
||||
|
потреб треб - ление , л / раст . |
рас ход . г / г . |
потреб треб - ление , л / раст . |
расход г / г . |
потреб треб - ление , л / раст . |
расход г / г . |
|
|
F. с ymosum (4n) |
2,26 |
469 |
3,10 |
233 |
5,36 |
296 |
|
F.tat.ssp.potanini |
1,20 |
340 |
5,30 |
300 |
6,5 |
307 |
|
F.tat.ssp.tataricum |
0,83 |
300 |
4,10 |
410 |
4,93 |
386 |
|
F.homotropicum (2n) |
0,99 |
477 |
5,35 |
583 |
6,34 |
564 |
|
F.homotropicum (4n) |
1,49 |
923 |
7,88 |
534 |
9,37 |
583 |
|
F.esc.ssp.ancestrale |
0,63 |
393 |
5,99 |
428 |
6,62 |
425 |
|
F.esc.ssp.esculentum: |
0,520,85 |
393982 |
2,065,44 |
307590 |
2,586,02 |
305602 |
|
Богатырь (2n) |
0,52 |
837 |
2,06 |
400 |
2,58 |
447 |
|
Молва (2n) |
0,56 |
982 |
2,79 |
431 |
3,35 |
476 |
|
Минчанка (4n) |
0,58 |
749 |
5,44 |
590 |
6,02 |
602 |
|
М -822 |
0,63 |
591 |
4.9 |
510 |
5,53 |
518 |
|
Botansoba |
0,85 |
393 |
4,18 |
379 |
5,03 |
395 |
|
Shinanonatsusoba |
0,67 |
401 |
4,19 |
307 |
4.86 |
305 |
|
Mankan Royal |
0,82 |
437 |
4,43 |
385 |
5,25 |
392 |
Сорта гречихи культурной (F.escul.ssp. esculenum) потребляли воды значительно меньше своих диких сородичей , в качестве которых могут считаться вид F.homotropicum и подвид F.escul.ssp.ancestrale. Особенно низким водопотреб - лением в связи с меньшей продолжительностью ве гетационного периода отличались сорта Восточно европейского экотипа , отселектированные и возде лываемые в России . Такая же закономерность ха рактерна и для окультуренного подвида гречихи татарской (F. tat. ssp. tataricum).
Наибольшими потребностями в воде растения диких видов и сортов гречихи культурной характеризовались в период их генеративного развития. Это связано со значительной его продолжительностью (в 2…3 раза) по сравнению с вегетативным, а также с большей биомассой и облиственностью. Абсолютные величины расхода воды на создание единицы сухого вещества в наших опытах были меньше, чем приводимые в спра- вочниках для гречихи культурной в связи с тем, что учитывалась масса сухого вещества не только надземной части растений, но и корневой системы, что в методическом отношении является более правильным.
Влияние условий произрастания на водопотреб - ление растениями различных сортов можно проил люстрировать результатами опыта по их выращива нию при различной влажности почвы ( табл . 4).
Таблица 4 - Влияние влажности почвы на водопотреб - ление растениями сортов гречихи культурной , г / раст .
|
Сорт |
Влажность почвы , % от ПВ |
|||
|
30 |
30/70 |
70 |
70/30 |
|
|
Шатиловская 5 |
1241 |
2428 |
2390 |
1640 |
|
Дождик |
1296 |
2810 |
2525 |
1695 |
|
Казанская крупноплодная |
944 |
2477 |
2392 |
1020 |
|
Мелколистная |
920 |
2448 |
2212 |
1034 |
|
ОВ - 4 |
1243 |
2843 |
2173 |
1858 |
|
Дождик треугольный |
712 |
1676 |
1805 |
1361 |
При недостатке влаги в почве все изучаемые сорта снижали расход воды , при этом у относительно мелко листных генотипов ( Казанская крупноплодная , Мел колистная , Дождик треугольный ) это снижение было наибольшим . Меньшей была потребность в воде у них и при оптимальной влажности почвы (70% от ПВ ). Из этого следует , что при прочих равных условиях отно шение растений к водообеспеченности определяется не только генотипом , но и их морфотипом через раз мер и форму листьев , как основных органов испаре ния . При изменении влажности почвы после начала цветения с недостаточной на оптимальную (30/70%) растения восстанавливают размеры водопотребления до уровня контрольных или даже превосходят их . Если же после начала цветения оптимальное водоснабжение изменялось на недостаточное (70 /30%), то наблюда лась сильная депрессия в их росте и водопотреблении .
Различия по отношению к водному фактору у ви дов и сортов гречихи начинают проявляться уже в пе риод прорастания семян . Это выявлено в опытах с соз данием условий физиологической сухости с помощью гипертонических растворов осмотиков . Согласно по лученным экспериментальным данным , наибольшей способностью к прорастанию в условиях физиологи ческой сухости отличался культивируемый подвид гречихи татарской ( к -17) и агроэкотипы гречихи куль турной ( рис . 2). Они сохраняли способность к прорас танию на 50% даже при величине осмотического дав ления в 12 атм ., тогда как семена диких видов и под видов давали такой процент прорастания при величине гипертонического раствора в 3…4 раза меньшей .
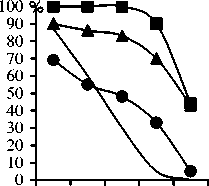
F.tataricum G. (к-17)
F. esculentum M.
—•— F. ssp.ancestrle O.
F. homotropicum О.
атм
0 3 6 9 12
Рисунок 2 - Динамика прорастания семян видов гречихи при различной величине осмотического давления раствора сахарозы , %
Все изученные виды гречихи отличались отно сительно слабой устойчивостью тканей различных органов к засухе и высокой температуре , в связи с отсутствием защитных свойств характерных , на пример , некоторым злаковым ( опушенность , воско вой налет и др .) и слабой выраженностью физиоло гических механизмов регулирования водного режи ма клеток и тканей . Одним из механизмов регулиро вания является способность тканей различных орга нов ( прежде всего листьев ) удерживать воду в клет ках , что обусловлено состоянием биоколлоидов их цитоплазмы . И как видно из данных , помещенных в таблице 5, наибольшей водоотдачей отличались ткани листьев тетраплоидной формы многолетнего вида гречихи (F. cymosum) и однолетнего диплоид ного вида F. homotropicum. Самая же высокая водо отдача была характерна для окультуренного подви да гречихи татарской (F. tat. ssp. tataricum)
Таблица 5 - Относительная засухоустойчивость видов и сортов гречихи
|
Вид , подвид , сорт , происхождение |
Вид устойчивости |
|
|
засухоустой чивость , % |
устойчивость к завяда нию (% водоотдачи ) |
|
|
F. cymosum (4n) |
5,3 |
48,5 |
|
F. tat. ssp . potanini |
0,0 |
37,6 |
|
F. tat. ssp. tataricum |
44,0 |
77,2 |
|
F. homotropicum (2n) |
3,0 |
45,3 |
|
F. homotropicum (4n) |
0,0 |
33,2 |
|
F. esc. ssp. ancestrale |
36,4 |
33,4 |
|
F. esc. ssp. esculentum: |
54,4 |
39,1 |
|
Богатырь ( Россия ) |
37,1 |
38,4 |
|
Молва ( Россия ) |
89,1 |
35,8 |
|
Минчанка ( Беларусь ) |
60,8 |
37,6 |
|
М - 822( Северный Китай ) |
53,7 |
40,5 |
|
Botansoba ( Япония ) |
51,9 |
36,7 |
|
Mankan Royal ( Канада ) |
34,1 |
45,8 |
Сорта различных экотипов гречихи культурной превосходили дикие виды по способности семян прорастать в условиях физиологической сухости , т . е . обладали высокой сосущей силой , и не имели су щественных преимуществ перед предковыми фор мами по водоудерживающей способности тканей листьев . С изменением условий произрастания ( в частности водообеспеченности растений ) меняется и водоудерживающая способность тканей . При недос татке влаги в почве уменьшается ее доступность для корневой системы в связи с тем , что сосущая сила почвенного раствора возрастает быстрее , чем сосу щая сила клеток корня . Уменьшение количества поступающей воды приводит к снижению водопо - требления , общей оводненности тканей растения , повышению их водоудерживающей способности и депрессии продукционного процесса .
Отношение к почве и потребность в питательных веществах. Предковые формы гречихи культурной и ряд диких видов произрастали на легких, хорошо аэрируемых, суглинистых и супесчаных почвах в долинах предгорных рек. Они характеризуются высоким содержанием питательных веществ, слабокислой или нейтральной реакцией почвенного раствора. Предпочтение к такому типу почв сохранили и современные сорта гречихи культурной и татарской [3]. Такое предпочтение по отношению к легким и хорошо аэрируемым поч- вам определяется также способностью растений образовывать на нижней половине подсемядольного колена стебля придаточные корни, которые хорошо развиваются при достаточном увлажнении, хотя в настоящее время эта культура возделывается на самых различных по физико-химическому составу почвах.
Корневая система гречихи отличается высокой усвояющей способностью элементов питания из почвы , в том числе из труднодоступных форм . Рас творяющая сила её корней по данным В . Дитриха была в 23 раза сильнее , чем у озимой ржи и в 12 раз больше , чем у пшеницы . По интенсивности погло щения гречиха превышала яровую пшеницу в 2,5 раза , а ячмень - более чем в 5 раз . Отмечается осо бенно высокая способность ее по сравнению с дру гими культурами , усваивать фосфор из труднодос тупных соединений [5].
Высокая потребность растений гречихи в азоте связана с формированием большой биомассы , а ка лий необходим для улучшения газообмена растений и синтеза углеводов , а также для упрочение тканей стебля . Фосфор требуется ей преимущественно во время формирования генеративных органов . Наи большая концентрация фосфорной кислоты в расте ниях обнаруживается в фазе бутонизации и начале цветения . Различия в количестве потребляемых пи тательных веществ видами и сортами гречихи опре деляются не столько концентрацией их в тканях различных органов , сколько их биомассой . Растения диких видов формировали в 1,5…2 раза большую биомассу , чем сорта гречихи культурной , а потому и потребность их в питательных веществах на ее соз дание была во столько же раз большей ( табл . 6).
Таблица 6 - Потребность растений различных видов и сортов гречихи в питательных веществах для формирования биологической продуктивности
|
Вид , подвид , сорт |
Элементы , мг / раст . |
||
|
N |
Р 2 О 5 |
К 2 О |
|
|
F. cymosum (4n) |
269 |
102 |
321 |
|
F. homotropicum (2n) |
256 |
96,9 |
317 |
|
F. esc. ssp. ancestrale |
262 |
92,4 |
287 |
|
F. esc. ssp. esculentum: |
122-165 |
42-53,7 |
140-198 |
|
Богатырь |
146 |
50,4 |
140 |
|
Баллада |
157 |
52,2 |
178 |
|
Дождик |
157 |
50,0 |
177 |
|
Дикуль |
165 |
53,7 |
198 |
|
Скороспелая 86 |
122 |
41,8 |
143 |
|
Botansoba |
153 |
47 |
189 |
|
Mankan Royal |
215 |
50 |
197 |
При этом у диких видов гречихи 80…90% посту пающих питательных веществ расходовалось на созда ние биомассы вегетативных органов и только 10…20% на формирование плодов . У сортов гречихи культурной на создание биомассы вегетативных органов идет около 50% потребляемого азота , 50…55% фосфора и около 80% калия . Поэтому различия в потреблении питатель ных веществ у сортов гречихи культурной в большей степени определяется уровнем урожайности плодов ( зерна ), чем массой вегетативных органов .
Выводы :
Таким образом, отношение растений гречихи к температурному фактору определяется их геномом с одной стороны, и климатическими условиями региона их произрастания или возделывания- с другой. Наиболее радикальным средством воздействия на этот признак является селекция, основанная на использовании мутаций из природного мутационного резерва культуры с последующей их селекционной доработкой. Однако вызванные изменения относятся к модификационным и не наследуются последующими поколениями.
Резюмируя отношение культуры гречихи к водно му фактору , можно сделать выводы о высоком ее вла - голюбии , слабой засухоустойчивости на всех этапах онтогенеза , при одновременно высокой репарацион ной способности , которая снижается по мере старения органов и растения в целом . Все это согласуется с про исхождением , филогенезом и формированием у видов защитно - приспособительных реакций и адаптивной стратегии в целом . Становление вида гречихи куль турной и экологическая специализация ее сортов в различных регионах земного шара привели ( при со хранении тенденции к влаголюбию ) к появлению в мутационном резерве её относительно менее влаголю бивых или более засухоустойчивых мутаций , из кото рых в процессе естественного или искусственного от боров сформировались сорта - популяции с изменен ным отношением к водному режиму .
Гречиха очень отзывчива на улучшение плодо родия почвы и применение минеральных удобре ний . В наибольшей степени на это отзываются ве гетативные органы растения , которые в этом случае могут израстать в ущерб формированию урожая плодов . Поэтому возделывание этой культуры на различных по плодородию почвах требует разработ ки оптимального уровня питания и сбалансирован ности питательных веществ .
Список литературы Отношение растений видов и сортов гречихи к основным абиотическим факторам
- Гродзинский, А.М. Краткий справочник по физиологии растений/А.М. Гродзинский, Д.М.Гродзинский. -Киев: Наукова думка, 1973. -592с.
- Дроздов, С.Н. Терморезистентность активно вегетирующих растений./С.Н. Дроздов, В.К. Курец, А.Ф. Титов -Л.,1984.-168с.
- Ефименко, Д.М. Агротехнические приемы получения высоких и устойчивых урожаев гречихи в условиях Полесья и Лесостепи УССР./Д.М. Ефименко. -Киев, 1964. -20с.
- Кротов, А.С. Гречиха -Fagopyrum Mill//Культурная флора СССР.Т.III.Крупяные культуры (гречиха, просо, рис)./А.С. Кротов-Л., Колос,1975.-С.7-118.
- Прянишников, Д.Н. Некоторые биологические и анатомические особенности корней гречихи/Д.Н. Прянишников//Докл. ТСХА. -1962. -т.77.-с.121-125
- Ohnishi, O. Discovery of new Fagopyrum shtcies and its implication for the studies of evolution of Fagopyrum and of the origin of cultivated buckwheat//Current Advances in Buckwheat Research. Proc. 6 th Intl. Symp./O. Ohnishi. -Buckmheat Japan, Shinshu University Press, 1995. -Р.175-190.
