Овуляторная функция крыс в условиях блокады мелатониновых рецепторов

Автор: Жукова Оксана Валентиновна, Обухова Елена Сергеевна, Хижкин Евгений Александрович, Тышдык Маргарита Леонидовна, Виноградова Ирина Анатольевна
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Физико-химическая биология
Статья в выпуске: 8 (153), 2015 года.
Бесплатный доступ
Исследована возрастная динамика эстральной функции самок крыс, находящихся с рождения в условиях стандартного освещения и получающих с пятимесячного возраста ежедневно 5 дней в неделю вместе с питьевой водой в ночное время блокатор мелатониновых рецепторов - лузиндол (N-Acetyl-2-benzyl-tryptamine) в концентрации 10 мг/л. В возрасте три, шесть, девять, двенадцать и пятнадцать месяцев у животных ежедневно в течение двух недель брали влагалищные мазки и проводили цитологическое исследование с оценкой длительности эстрального цикла и соотношения фаз овуляторного цикла. Рассчитывали относительное число коротких, средних и длинных циклов; процентное соотношение фаз эстрального цикла; относительное число животных с иррегулярными циклами. В результате зафиксировано уменьшение количества коротких и увеличение количества длинных эстральных циклов через один месяц применения лузиндола. Установлено, что в условиях блокады мелатониновых рецепторов наблюдалось появление преждевременных возрастных изменений эстральной функции у самок крыс: достоверное уменьшение количества регулярных циклов и появление иррегулярных эстральных циклов (с 6-месячного возраста); более раннее увеличение продолжительности овуляторного цикла (с 9-месячного возраста). По всей видимости, блокада мелатониновых рецепторов могла привести к усилению секреции гонадолиберинов и гонадотропинов, в результате чего активация роста и развития фолликулов в яичниках способствовала более длительному выделению эстрогенов и изменению фаз овуляторного цикла. Несмотря на различные механизмы развития недостаточности мелатонинергической системы - снижение выработки мелатонина (при воздействии постоянного освещения) или блокада мелатониновых рецепторов (при применении лузиндола) - эффекты действия на овуляторную функцию схожи. При фармакологической блокаде мелатониновых рецепторов не было зафиксировано ни одного животного с синдромом персистирующего эструса, что косвенно указывает на наличие безрецепторной гормональной регуляции мелатонина через систему половых гормонов.
Крысы, лузиндол, эстральный цикл
Короткий адрес: https://sciup.org/14751008
IDR: 14751008 | УДК: 612.4.018+577.345
Rats’ ovulatory function under melatonin receptors’ blockade
The paper investigates the problem of age dynamics of the estrous function in female rats under standard lighting from the time of birth. On the daily basis, 5 days a week, beginning from the age of five months the rats received along with drinking water a blocker melatonin receptor - luzindol (N-Acetyl-2-benzyl-tryptamine) at the concentration level of 10 mg / l. At the age of three, six, nine, twelve, and fifteen months daily, for a period of two weeks, vaginal smears and cytological examination was performed to estimate the length of the estrous cycle and the phase relationship in ovulatory cycle. The number of short, medium and long cycles; the percentage of the phases of the estrous cycle; relative number of animals with irregular cycles were calcultated. As a result, it recorded a decrease in the amount of short cycles and an increase in the amount of long estrous cycles after one month of luzindol application. It was found out that melatonin receptor blockade influence the appearance of premature aging changes in female rats: a significant decrease in the number of regular cycles and stable occurrence of irregular estrous cycles (6 months of age); earlier prolongation of ovulatory cycle (9 months of age). Apparently, melatonin receptor blockade could lead to an increased secretion of gonadotropins and gonadoliberines. This resulted in the activation of the follicles’ growth and ovaries’ development and contributed to a sustained release of estrogen and the phase change in the ovulatory cycle. Despite different mechanisms affecting the failure in melatoninergic system: loss of melatonin (under the influence of constant light) or melatonin receptor blockade (usage of luzindol) - the effects of actions on the ovulatory function is similar. No rodents with the syndrome of persistent estrus were registered under the pharmacological blockade of melatonin receptor. The obtained results point to the presence of hormonal regulation through sexual hormones.
Текст научной статьи Овуляторная функция крыс в условиях блокады мелатониновых рецепторов
Репродуктивная функция самок млекопитающих находится под контролем гипоталамуса, регуляторная деятельность которого осуществляется посредством гормона мелатонина [1]. Основной эффект мелатонина заключается в торможении секреции тропных гормонов передней доли гипофиза и в модифицирующем влиянии на нейросекреторную активность гипоталамуса. Независимо от того, активно животное днем или ночью, максимальный уровень мелатонина наблюдается всегда во время темновой фазы естественного или искусственно созданного цикла чередования дня и ночи [11].
Известно, что введение лузиндола – антагониста мелатониновых рецепторов МТ1 и МТ2 значительно ослабляет эффекты экзогенного и эндогенного мелатонина [14], [16], [17]. Исходя из этого представляется интересным изучение функционирования репродуктивной системы самок млекопитающих в условиях угнетения мелатониновых сигнальных механизмов.
Целью данного исследования явилось изучение эстрального цикла самок крыс в условиях блокады мелатониновых рецепторов лузиндолом.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования были проведены на 40 самках крыс Вистар собственной разводки, первоначально полученных из питомника лабораторных животных «Пущино» (научно-производственное подразделение Филиала Института биооргани-ческой химии им. академиков М. М. Шемякина и Ю. А. Овчинникова, Московская область). Животных содержали при температуре воздуха в помещении 22 ± 2 °С в клетках размером 40 × 60 × 20 см по 10 особей в каждой с обеспыленной подстилкой из древесной стружки. В помещении были соблюдены условия стандартного чередующегося режима освещения (12 ч. свет, 12 ч. темнота LD): люминесцентные лампы (освещенность 750 лк на уровне клеток) освещали помещение в течение 12 дневных часов, на ночь (12 ч.) свет отключали; окна в помещении отсутствовали.
Все крысы получали гранулированный корм (ГОСТ Р 50258-92 «Комбикорма полнорационные для лабораторных животных. Технические условия»: содержание протеина более 25 %, жиров 6–12 %, кальция и фосфора 1–1,4 %, лизина и метионина 0,7–1,5 %, микроэлементов и витаминов 3–4 %) и отстоянную водопроводную воду без ограничения.
Работа была выполнена с соблюдением международных принципов Хельсинкской декларации о гуманном отношении к животным, прин- ципов гуманности, изложенных в директиве Европейского Сообщества (2010/63/ЕU), «Био-этических правил проведения исследований на человеке и животных», «Правил проведения работ с использованием экспериментальных животных», «Санитарно-эпидемиологических требований к устройству, оборудованию и содержанию экспериментально-биологических клиник (вивариев)», «Руководства по лабораторным животным и альтернативным моделям в биомедицинских технологиях» [12], [13].
В пятимесячном возрасте самки крыс были рандомизированно (по весу) разделены на две группы. Животные первой группы (n = 20; LD + luz) получали ежедневно 5 дней в неделю с питьевой водой в ночное время с 20:00 до 08:00 лузиндол (N-Acetyl-2-benzyl-tryptamine) фирмы «Bachem AG» (Швейцария) в концентрации 10 мг/л. Поилки, в которых находился лузиндол, были изготовлены из темного непрозрачного полимерного материала. Свежий раствор препарата готовили ежедневно. Вторая группа (n = 20; LD) в ночное время получала питьевую воду в том же объеме и явилась контрольной.
В возрасте три, шесть, девять, двенадцать и пятнадцать месяцев у животных ежедневно в течение двух недель брали влагалищные мазки для изучения эстральной функции. У грызунов содержимое влагалища в зависимости от фазы овуляторного цикла имело различный состав, который можно определить при микроскопировании. Весь овуляторный цикл крысы делился на следующие стадии: 1) диэструс, или стадия покоя; 2) проэструс, или стадия подготовки к течке; 3) эструс, или стадия течки; 4) метаэструс, или стадия после течки. В соответствии с данными фазами изменялся состав содержимого влагалища. Мазки готовили по общепринятой методике из влагалищного содержимого, полученного утром в одно и то же время посредством введения во влагалище крыс ватного тампона. Свежеприготовленные мазки без фиксации и окраски изучали под бинокулярным микроскопом при увеличении в 70 раз (об. 10, ок. 7) с опущенным конденсором микроскопа. Результаты исследования вагинального содержимого фиксировали в журнале. Проводили оценку следующих параметров: длительность эстрального цикла; соотношение фаз овуляторного цикла; относительное число коротких, средних и длинных циклов; процентное соотношение фаз эстрального цикла; относительное число животных с иррегулярными циклами [9].
Статистическую обработку цифрового материала проводили с использованием общеприня- тых методов вариационной статистики в среде программы EXCEL. Достоверность результатов оценивали по непараметрическому U-критерию Уилкоксона – Манна – Уитни и методом χ2. Достоверно различающимися признавали значения при р < 0,01 и р < 0,05; при 0,05 < р < 0,1 различия между средними имели тенденцию к достоверности [8].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ОБСУЖДЕНИЕ
В физиологических условиях к возрасту 15–18 месяцев у крыс появляются признаки старения репродуктивной системы: короткие эстральные циклы исчезают, преобладают длинные овуляторные циклы (более 7 дней), снижается количество регулярных циклов, появляются иррегулярные циклы [3], [7].
В ходе наших исследований установлено, что в контрольной группе короткие циклы сохранялись и в зрелом возрасте, хотя их количество с возрастом достоверно снижалось с 54 % в 6 месяцев до 12 % к 15-месячному возрасту, причем уже в возрасте одного года количество коротких эстральных циклов было достоверно меньше (p < 0,05). На втором году жизни у самок контроля преобладали циклы средней продолжительности (достоверное увеличение наблюдали также с 12-месячного возраста). В опытной группе установлена аналогичная закономерность, но при этом через месяц приема лузиндола доля коротких циклов достоверно уменьшилась по сравнению с группой контроля и в возрасте 6 месяцев составила всего 14 %, а в 15-месячном возрасте короткие эстральные циклы у крыс дан- ной группы отсутствовали. Начиная с 6-месячного возраста в группе животных, получавших лузиндол, достоверно преобладали средние эстральные циклы. Различия исследуемых показателей контрольной и опытной групп начиная с возраста 6 месяцев являются достоверными (p < 0,05) (таблица). В возрасте одного года у самок, получавших лузиндол, преобладали эстральные циклы с длительностью более 7 дней, в дальнейшем происходило перераспределение эстральных циклов разной длительности за счет появления иррегулярных овуляторных циклов. Доля длинных циклов с 6-месячного возраста в группе LD + luz была достоверно выше по сравнению с опытной группой. К 15-месячному возрасту данный показатель возрос до 29 % в группе контроля и до 42 % в группе, получавшей лузин-дол. Установленные различия также являются достоверными (p < 0,05). Полученные результаты свидетельствуют о более быстром старении репродуктивной системы крыс, подвергавшихся блокаде мелатониновых рецепторов при действии лузиндола.
В трехмесячном возрасте длительность эстрального цикла у самок крыс составляла 3,7 ± 1,1 дня. В контрольной группе зафиксировано достоверное увеличение средней продолжительности овуляторного цикла к 12-месячному возрасту – 5,2 ± 1,2 дня (p < 0,05). В опытной группе длительность эстрального цикла уже в возрасте 6 месяцев была достоверно больше по сравнению с параметром, полученным в возрасте 3 месяцев, и составляла 5,9 ± 1,5 дня (p < 0,05). В последующие возрастные периоды данный
Возрастная динамика показателей эстральной функции у крыс
|
Возраст, мес. |
Средняя продолжительность эстрального цикла, дни |
Количество эстральных циклов разной длительности, % |
Соотношение фаз овуляторного цикла, % |
Число исследованных эстральных циклов, % |
||||
|
Короткие |
Средние |
Длинные |
Эструсы |
Диэструсы |
Нормальные циклы |
Иррегулярные циклы |
||
Стандартное освещение (LD)
|
6 |
4,8±1,4 |
54 в |
43 в |
3 в |
37 |
63 |
100 |
0 |
|
9 |
4,9±1,9 |
47 в |
43 в |
10 б, в |
58 б |
42 б |
93 в |
3 в |
|
12 |
5,2±1,2 а |
28 б, в |
64 б, в |
8 в |
55 б |
45 б |
89 б, в |
11 б, в |
|
15 |
6,1±1,5 а |
12 б, в |
59 б |
29 б, в |
45 |
55 |
81 б, в |
19 б, в |
Стандартное освещение + лузиндол (LD + luz)
|
6 |
5,9 ± 1,5 а |
14 в |
68 в |
18 в |
32 |
68 |
97 |
3 |
|
9 |
6,6 ± 2,0 а |
9 в |
64 в |
27 в |
64 б |
36 б |
88 б, в |
12 б, в |
|
12 |
7,2 ± 1,8 а |
7 в |
39 б, в |
54 б, в |
56 б |
44 б |
69 б, в |
31 б, в |
|
15 |
7,3 ± 1,8 а |
0 б, в |
58 |
42 б, в |
41 |
59 |
67 б, в |
33 б, в |
Примечание . а р < 0,05 – различия в группе с показателем в трехмесячном возрасте достоверны (критерий Уилкоксо-на – Манна – Уитни); б р < 0,05 – различия в группе с показателем в шестимесячном возрасте достоверны (метод χ2); в р < 0,05 – различия с показателем в группе LD в соответствующем месяце достоверны (метод χ2).
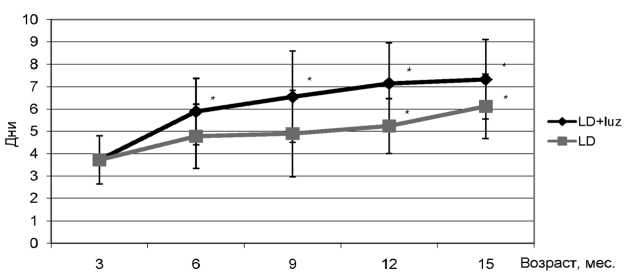
Рис. 1. Продолжительность эстрального цикла у самок крыс. р < 0,05 – различия в группе с показателем в трехмесячном возрасте достоверны (критерий Уилкоксона – Манна – Уитни); LD – стандартное освещение; LD + luz – лузиндол
показатель достоверно увеличивался до 7,3 ± 1,8 дня в возрасте 15 месяцев (рис. 1). В предыдущих исследованиях показано достоверное удлинение эстрального цикла с 8-месячного возраста у крыс-самок, находящихся в период прогрессивного роста в условиях естественной повышенной освещенности (белые ночи) или в условиях постоянного искусственного освещения [5], [6], [7]. Содержание животных в условиях постоянного или естественного освещения в период белых ночей приводило к нарушению работы шишковидной железы и угнетению продукции и секреции мелатонина – основного гормона эпифиза [1], что являлось причиной ранних возрастных изменений в репродуктивной системе крыс. В нашей работе применение лузиндола, блокирующего мелатониновые рецепторы, соответствовало эффектам, возникающим при физиологической эпифизэктомии, приводящей к угнетению выработки мелатонина. В других исследованиях показано, что искусственное увеличение продолжительности светового периода в течение дня даже на 2–4 часа, при котором не происходило достаточной секреции мелатонина, приводило у грызунов к увеличению продолжительности эстрального цикла, а в некоторых случаях – к его нарушению. Если воздействие света на крыс увеличить до 24 часов в сутки, у большинства из них в короткие сроки развивался синдром персистирующего (постоянного) эструса [2], [3]. В наших исследованиях применение лузиндола приводило у крыс-самок к более раннему увеличению средней продолжительности овуляторного цикла с возраста 9 месяцев, таким образом, показатели, полученные при помощи фармакологической блокады мелатониновых рецепторов, соответствовали параметрам при так называемой физиологической пинеалэктомии, которая достигается постоянным освещением животных [5], [6], [7].
При изучении соотношения фаз овуляторных циклов установлена тенденция к уменьшению доли диэструсов и увеличению доли эструсов с возрастом в обеих группах (см. табл.). У крыс группы LD доля диэструсов в 6 месяцев составила 63 %, в 15 месяцев – 55 %, у крыс группы LD + luz аналогичный показатель равнялся 68 % в возрасте 6 месяцев и 59 % в возрасте 15 месяцев. Соответственно, доля эструсов у крыс-самок контрольной группы в 6-месячном возрасте составляла 37 %, а у крыс-самок опытной группы – 32 %. С возрастом в контрольной группе количество эструсов изменялось незначительно, в то время как в опытной группе количество эструсов увеличилось в два раза в возрасте 9 месяцев, затем уменьшилось к 15 месяцам и составило 41 % от общего количества фаз овуляторного цикла. Достоверных различий данных показателей между группами не установлено.
С возрастом у животных было зафиксировано появление иррегулярных циклов. В группе контроля данный показатель впервые был отмечен в 9 месяцев и составил 3 % от общего количества. Достоверное увеличение доли иррегулярных циклов наблюдалось с 12-месячного возраста. В опытной группе данный показатель зафиксирован раньше – в возрасте 6 месяцев. Достоверное увеличение доли иррегулярных циклов отмечено с 9 месяцев. В 15 месяцев у животных опытной группы зафиксировано максимальное количество иррегулярных циклов – 33 %, у животных контрольной группы – 19 %. Достоверные различия между группами наблюдались с возраста 9 месяцев (рис. 2). В исследованиях Д. А. Батурина, И. Н. Алимовой и др. у мышей HER-2/neu, содержавшихся с 2 месяцев при постоянном освещении, в возрасте 9 месяцев иррегулярные циклы обнаруживали у 76 % самок, в то время как у животных, содержавшихся в стандартном режиме, у 50 % самок [4]. В других исследованиях при содержании крыс в условиях постоянного и характерного для Карелии естественного освещения с периодом белых ночей отмечено появление иррегулярных циклов в молодом (5-месячном) возрасте. К 23-месячному возрасту количество иррегу-
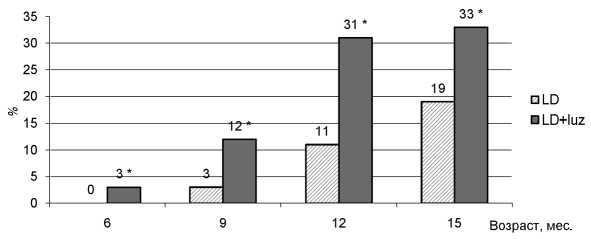
Рис. 2. Количество иррегулярных циклов (%) у самок крыс. * р < 0,05 – различия с показателем в группе LD в соответствующем месяце достоверны (метод χ 2 ). Обозначения, как на рис. 1
лярных циклов у животных этих групп возросло до 80 и 64 % соответственно [5]. По данным Д. В. Клочкова и Д. К. Беляева, продолжительное освещение с 2-месячного возраста крыс Вистар в течение 1, 2 или 6 месяцев приводило к появлению персистирующего эструса у 24, 61 и 100 % животных соответственно [10]. В другом исследовании у 100 % крыс Вистар отмечали постоянный эструс при содержании с 4-месячного возраста в течение 3 месяцев в условиях постоянного освещения [15]. В отличие от данных экспериментов в наших исследованиях не установлено наличие персистирующего эструса в опытной группе, что косвенно указывает на различие в ответах репродуктивной системы на снижение выработки мелатонина или блокаду мелатониновых рецепторов.
ЗАКЛЮЧЕНИЕ
Таким образом, в ходе десятимесячного исследования эстральной функции крыс-самок, получающих с возраста 5 месяцев лузиндол, выявлены ранние возрастные изменения репродуктивной системы по сравнению с группой контроля. У крыс опытной группы зафиксировано достоверное более раннее увеличение продолжительности овуляторного цикла с 9-месячного возраста, снижение количества регулярных циклов, появление иррегулярных циклов, уменьшение количества коротких и увеличение количества длинных эстральных циклов через 1 месяц приема лузиндола. Вероятно, применение лузиндола, результатом действия которого является блокада мелатониновых рецепторов, приводило к более выраженным возрастным изменениям репродуктивной системы. По всей видимости, блокада мелатониновых рецепторов, находящихся в гипофизе и гипоталамусе, могла привести к усилению секреции гонадолиберинов и гонадотропинов, в результате чего активация роста и развития фолликулов в яичниках способствовала более длительному выделению эстрогенов и изменению фаз овуляторного цикла. Несмотря на различные механизмы развития недостаточности мелатонинергической системы (снижение выработки мелатонина при воздействии постоянного освещения [5], [6], [7] и блокада мелатониновых рецепторов при применении лузиндола), эффекты действия на овуляторную функцию схожи: нарушения, возникающие в работе репродуктивной системы, характеризуются более быстрыми возрастными изменениями эстрального цикла. При фармакологической блокаде мелатониновых рецепторов не зафиксировано ни одного животного с синдромом персистирующего эструса, что косвенно указывает на наличие безрецепторной гормональной регуляции мелатонина через систему половых гормонов.
* Работа выполнена в рамках Программы стратегического развития ПетрГУ на 2012–2016 гг. «Университетский комплекс ПетрГУ в научно-образовательном пространстве Европейского Севера: стратегия инновационного развития», «УМНИК – 2013» и гранта РФФИ № 15-34-50841.
RATS’ OVULATORY FUNCTION UNDER MELATONIN RECEPTORS’ BLOCKADE
Список литературы Овуляторная функция крыс в условиях блокады мелатониновых рецепторов
- Анисимов В. Н. Молекулярные и физиологические механизмы старения: В 2 т. 2-е изд., перераб. и доп. СПб.: Наука, 2008. Т. 1. 468 с.
- Анисимов В. Н., Айламазян Э. К., Батурин Д. А. Световой режим, ановуляция и риск злокачественных новообразований женской репродуктивной системы: механизмы связи и профилактика//Журнал акушерства и женских болезней. 2003. Т. 52. № 2. С. 47-57.
- Анисимов В. Н., Батурин Д. А., Айламазян Э. К. Эпифиз, свет и рак молочной железы//Вопросы онкологии. 2002. Т. 48. № 4-5. С. 524-535.
- Батурин Д. А., Алимова И. Н., Попович И. Г, Анисимов В. Н., Забежинский М. А., Яшин А. И., Семенченко А. В. Влияние световой депривации на показатели гомеостаза, продолжительность жизни и развитие спонтанных опухолей у трансгенных мышей HER-2/neu//Вопросы онкологии. 2004. Т. 50. № 3. С. 332-338.
- Виноградова И. А., Чернова И. В. Влияние светового режима на возрастную динамику эстральной функции и уровня пролактина в сыворотке крови у крыс//Успехи геронтологии. 2006. № 19. С. 60-65.
- Виноградова И. А., Чернова И. В. Световые режимы и овуляторная функция у крыс в онтогенезе//Российский физиологический журнал. 2007. Т. 93. № 1. С. 90-98.
- Гайдин И. В., Баранова Ю. П., Виноградова И. А. Влияние продолжительности светового дня в условиях Карелии на показатели эстральной функции у самок крыс//Ученые записки Петрозаводского государственного университета. 2011. № 6 (119). С. 45-49.
- Зайцев В. М., Лифляндский В. Г., Маринкин В. И. Прикладная медицинская статистика. СПб., 2006. 432 с.
- Кабак Я. М. Практикум по эндокринологии. Основные методики экспериментально-эндокринологических исследований. М.: Изд-во МГУ, 1968. 153 с.
- Клочков Д.В., Беляев Д. К. Эффект постоянного освещения на репродуктивную функцию крыс//Онтогенез. 1977. Т. 8. № 5. С. 487-496.
- Мелатонин: теория и практика/А. Ю. Беспятых ; Под ред. С. И. Рапопорта, В. А. Голиченкова. М.: Медпрактика, 2009. 99 с.
- Руководство по лабораторным животным и альтернативным моделям в биомедицинских исследованиях/Под ред. Н. Н. Каркищенко, С. В. Грачева. М.: Профиль, 2010. 358 с.
- Этическая экспертиза биомедицинских исследований: Практические рекомендации/Под ред. Ю. Б. Белоусова. М.: Российское общество клинических исследований, 2005. 156 с.
- Drobnik J., Owczarek K., Piera L., Tosik D., Olczak S., Ciosek J., Hrabec E. Melatonin-induced augmentation of collagen deposition in cultures of fibroblasts and myofibroblasts is blocked by luzindole -A melatonin membrane receptors inhibitor//Pharmcological reports. 2013. Vol. 65. № 3. P. 642-649.
- Prata Lima M. F., Baracat E. C., Simones M. J. Effects of melatonin on the ovarian response to pinealectomy or continuous light in female rats: similarity with polycystic ovary syndrome//Brazil. J. Med. Biol. Res. 2004. Vol. 37. P. 987-995.
- Soares J. M., Masana M. I., Ersahin C., Dubocovich M. L. Functional melatonin receptors in rat ovaries at various stages of the estrous cycle//J. Pharmacol. Exp. Ther. 2003. № 306 (2). Р. 694-702.
- Wincz y k K., Fuss-Chmielewska J., Pawlikowski M., Karasek M., Lawnicka H. Luzindole but not 4-phenyl-2-propionamidotetralin (4P-PDOT) diminishes the inhibitory effect of melatonin on murine Colon 38 cancer growth in vitro//Neuroendocrinology letters. 2009. Vol. 30. P. 657-662.
