Oxytropis stukovii Palib. - редкий вид остролодочника Восточного Забайкалья

Автор: Селютина Ирина Юрьевна, Санданов Денис Викторович
Журнал: Вестник Бурятского государственного университета. Философия @vestnik-bsu
Рубрика: Ботаника
Статья в выпуске: 4, 2015 года.
Бесплатный доступ
Описаны биоморфология и онтогенез редкого эндемичного вида Oxytropis stukovii, изучена онтогенетическая структура популяции. Онтогенез вида включает 4 периода и 8 возрастных состояний. Факторами, ограничивающими распространение данного вида, по нашему мнению, являются его стенотопность, а также резкие колебания численности и онтогенетической структуры популяций в зависимости от климатических условий разных лет.
Редкий вид, эндемик, морфология, онтогенез, онтогенетическая структура
Короткий адрес: https://sciup.org/148182936
IDR: 148182936 | УДК: 581.522.582.736
Oxytropis stukovii Palib. as the rare species of Oxytropis DC. of the Eastern Zabaikalye
The biomorphology and ontogenesis of the rare endemic species Oxytropis stukovii have been outlined and the onthogenetic structure of the population has been studied. The species ontogenesis includes 4 periods and 8 age stages. In our opinion, the factors limiting this species spreading are its steno- topic nature and also sharp fluctuations in quantity and ontogenetic structure of populations depending on climatic conditions within different years.
Текст научной статьи Oxytropis stukovii Palib. - редкий вид остролодочника Восточного Забайкалья
Oxytropis stukovii Palib. Остролодочник Стукова — эндемик, уязвимый вид узколокальных местообитаний со статусом 3 (R) — редкий вид, внесен в Красную книгу Читинской области и Агинского Бурятского автономного округа [5]. O. stukovii встречается на территории Байкальской Сибири и на прилегающей части Монголии (Хангайский и Восточно-Монгольский флористические районы) [2]. Этот вид приурочен к засоленным пустынно-степным ландшафтам и, по мнению ряда авторов, связан с древними пустынными или пустынно-степными ландшафтами [7; 11].
O. stukovii в Читинской области встречается только по берегам нескольких соленых озер и на солонцеватых степях: в окрестностях озер Ножий, Зун-Соктуй и Булун-Цаган, близ сел Дылыгыр, Агинское и Нижний Цасучей [15]. Типичные местообитания вида — окраины солончаков, где он в отдельные благоприятные годы в период цветения образует красочные солончаковые луга [5]. В прибрежной зоне западного берега оз. Ножий описаны сообщества с доминированием данного вида [4]. При этом в северо-восточной части озера отмечены единичные особи вида во второй прибрежной полосе [6]. Основными лимитирующими факторами служат резкое понижение уровня водоемов в засушливые годы, когда вид почти полностью переходит в состояние покоя и может исчезнуть в связи с интенсивным выпасом овец [5].
Общеизвестно, что для оценки состояния и сохранения редких видов необходимо изучение их биологических особенностей. С этой целью нами были изучены биоморфология, онтогенез Oxytropis stukovii , а также динамика онтогенетической структуры ценопопуляции данного вида в окрестностях оз. Ножий .
Материал и методы
При выделении возрастных состояний придерживались общепринятой методики [13; 14]. Поскольку остролодочник Стукова является редким и эндемичным видом, изучение его биоморфологии и структуры ценопопуляции проводили с минимальным изъятием особей [3]. Диагнозы возрастных состояний составляли на основе морфологических описаний 10–20 особей каждого возрастного состояния. Растения всех изученных онтогенетических состояний были исследованы в природной популяции. Особи прегенеративного периода были также изучены в интродукционном эксперименте в условиях Центрального сибирского ботанического сада СО РАН.
Результаты и их обсуждение
В период с 2007 по 2014 г. нами была проведена ревизия известных местонахождений изучаемого вида. Были осуществлены поисковые работы в окрестностях села Агинское, озер Зун-Соктуй, Кункур и Ножий. В результате была обнаружена лишь небольшая популяция данного вида в прибрежной полосе оз. Ножий. В окрестностях оз. Кункур были отмечены единичные особи, а полное высыхание оз. Зун-Соктуй, по-видимому, привело к исчезновению вида в данном местообитании.
Популяция вида на оз. Ножий входит в состав злаково-даурскокопеечниковой ассоциации на дресвяно-песчаной засоленной почве. Травостой в изученном сообществе разреженный, общее проективное покрытие сильно варьирует в пределах изученной площади (100 м2) и составляет на различных участках от 5 до 50 %. В травостое доминирует Hedysarum dahuricum Turcz. ex B. Fedtsch. (20 %), в меньшем обилии встречаются Saussurea salsa (Pall. ex M. Bieb.) Spreng. (5 %), Oxytropis stukovii Palib. (3 %), Hordeum brevisubulatum (Trin.) Link (3 %) Argusia rosmarinifolia (Willd.) Steven (2 %). В травостое также отмечены Calamagrostis epigeios (L.) Roth., Elymus sibiricus L., Artemisia scoparia Waldst. & Kit., A. mongolica (Besser) Fisch. ex Nakai, Carex reptabunda (Trautv.) V. Krecz., Elytrigia repens (L.) Gould.; Astragalus adsurgens Pall., Crepis tectorum L., Senecio jacobaea L., Sonchus arvensis L., Taraxacum sinicum Kitag., Lepidium densiflorum Schrad., Lappula anisacantha (Turcz. ex Bunge) Guerke.
Изучение особенностей биоморфологии вида показало, что Oxytropis stukovii является стержнекорневым поликарпическим травянистым гемикриптофитом с многоглавым погруженным каудексом, вегетативными розеточными полициклическими и удлиненными пазушными монокарпическими генеративными побегами. Корневая система представлена тонким, маловетвящимся, стержневым корнем, в результате его контрактильной деятельности каудекс оказывается постоянно погруженным в почву. Размножение осуществляется только семенным путем. В старом генеративном и постгенеративных состояниях наблюдается сенильная партикуляция без омоложения.
Онтогенез O. stukovii включает в себя 4 периода: латентный, прегенеративный, генеративный и постгенеративный и 8 возрастных состояний.
Латентный период. Семена мелкие, округло-почковидные, зеленовато-коричневые или оливковые, длиной 1–1,3 мм и 0,7–0,9 мм шириной; созревают в конце июля — в начале августа. Лабораторная всхожесть семян после 6 мес. хранения составила 96,7 %.
Прегенеративный период. Проростки — растения до 1,2 см высотой с гипокотилем длиной 1,2– 1,5 см. Семядольные листья продолговатые (2х4 мм), 1–2 первых настоящих листа растут вертикально или косо вверх. Корень стержневой достигает 5 см. Ювенильные растения 1–3 года жизни с розеточным побегом, имеющим от 3 до 6 листьев. Листья могут быть двупарные, пятипарные, шестипарные, а также с 7–8 мутовками по 3–4 мелких листочка в каждой. Длина листьев от 1,5 до 8 см. Имма-турные растения — растения 2–6 года жизни с одним розеточным побегом первого порядка. Листьев на побеге 5–7, лист с 7–16 мутовками по 4 листочка в каждой. Длина листа достигает 10 см. Формируется и становится заметным каудекс (диаметр его 0,5 см). Виргинильные растения имеют хорошо развитый каудекс диаметром до 3,5 см. Число розеточных побегов увеличивается до 3–13, на каждом побеге 6–7 листьев взрослого типа с 18–26 мутовками. Длина листа достигает 14–16,5 см.
Генеративный период. У молодых генеративных растений каудекс крепкий и плотный, диаметром до 6 см. Формируются генеративные побеги, их число достигает 13. Вегетативный розеточный побег один, на нем располагается 4–7 листьев взрослого типа с 18–26 мутовками. Многоглавый каудекс средневозрастных генеративных растений достигает в диаметре 16 см, на нем моноподиально нарастают боковые розеточные побеги, в розетке 5–9 листьев взрослого типа. Число генеративных побегов составляет 34–50. У старых генеративных растений каудекс многоглавый, диаметром до 12 см с обособленными, часто отмершими, главами. На растениях формируется 7–8 генеративных побегов.
Постгенеративный период. Характерными для субсенильных особей являются следующие признаки: побеги только вегетативные розеточные в числе 3–5 с листьями взрослого и переходного к сенильному типов (число мутовок сокращается до 15). Каудекс частично разрушенный, партикулирует без интеграции. Сенильное возрастное состояние выражено неясно, в изученной популяции нами не были обнаружены особи с сенильными признаками.
Необходимо отметить, что у 30 % генеративных средневозрастных особей в изученной популяции встречалась пролиферация соцветий, т. е. прорастание соцветий побегом. Есть данные о том, что крайне неблагоприятные погодные условия могут быть причиной пролиферации соцветий [1; 12]. В данном случае причиной пролиферации соцветий у особей генеративного состояния, вероятно, послужили экстремальные климатические условия последнего десятилетия. Так, для Восточного Забайкалья в последние десятилетия отмечаются положительные тренды температур [8; 9]. Кроме этого, с 1999 г. началась фаза пониженной увлажненности, которая характеризовалась более высокой температурой воздуха по сравнению с аналогичной фазой предыдущего цикла [10]. В результате повышение температуры в сочетании с сокращением осадков привело к аридизации климата. Проявление таких процессов, как почвенная засуха на фоне общей аридизации, также создало неблагоприятные условия для растений степной зоны Восточного Забайкалья.
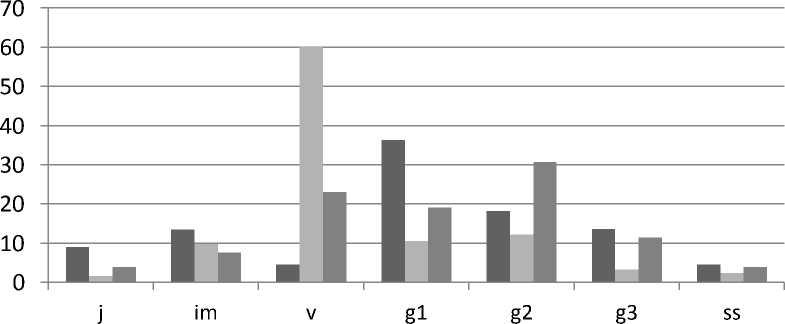
■ 2007 год ■ 2012 год ■ 2014 год
Рис. 1. Онтогенетическая структура ценопопуляции Oxytropis stukovii в разные годы исследований
Исследованная ценопопуляция Oxytropis stukovii на учетных площадках отличалась достаточно высокими показателями плотности 7,3–8,1 особей/м2 и неполночленностью онтогенетического спектра (отсутствуют сенильные особи) (рис. 1). В 2007 г. онтогенетический спектр характеризовался значительным преобладанием генеративной группы особей (72,8 % от общего числа особей). Большая жизнеспособность особей генеративного состояния позволяет им успешно существовать даже в условиях многолетней засухи. Однако за пятилетний период прошла довольно большая элиминация генеративных растений и в значительной степени увеличилась численность виргинильных особей. По-видимому, несколько лучшие условия увлажнения в данный период способствовали лучшему прорастанию семян и развитию растений прегенеративного периода. Этому процессу также благоприятствовало наличие запаса семян в почве, который сформировался в предыдущие годы, когда наблюдалось преобладание генеративных растений в популяции. Небольшое количество виргиниль-ных особей в 2007 г. свидетельствовало о значительной разнице в пополнении популяции молодыми особями в разные годы и невысокой выживаемости прегенеративных растений в неблагоприятных условиях среды. Необходимо отметить, что период с 2004 по 2007 г. характеризовался самыми минимальными годовыми суммами осадков за весь период наблюдений [10]. По-видимому, эти условия и оказали влияние на онтогенетическую структуру ценопопуляции O. stukovii. В последующие годы условия увлажнения были заметно лучше, что благоприятно сказалось на численности особей преге- неративного периода. При этом необходимо отметить, что состояние популяций O. stukovii также тесно связано с динамикой уровня воды в озере Ножий, поскольку в периоды наибольшего наполнения озера наблюдаются самые лучшие условия увлажнения для прибрежных растительных сообществ и ценопопуляций некоторых галофитных видов, таких как Hedysarum dahuricum, Saussurea salsa, Argusia rosmarinifolia, Oxytropis stukovii и O. varlakovii.
Онтогенетические спектры 2012 и 2014 гг. показывают определенные перспективы для дальнейшего развития изученной ценопопуляции. Наличие небольшого числа генеративных растений (26,1 %) в 2012 г. сменилось большой численностью растений генеративного периода в 2014 г. (61,4 %). В разные годы исследований доля особей субсенильного состояния в ценопопуляции обычно невелика — 2,4–4,6 %. Сенильных особей не обнаружено, что может свидетельствовать о невысокой жизненности растений в этом возрастном состоянии и может быть связано с относительно недолгим пребыванием растений в постгенеративном периоде.
Заключение
Oxytropis stukovii относится к типу моноцентрических каудексообразующих биоморф, онтогенез вида включает 4 периода и 8 возрастных состояний. Динамика онтогенетической структуры изученной популяции носит волнообразный характер. Для сохранения вида в данном местообитании необходим мониторинг численности популяции, особенно в засушливые годы, а также продолжение исследований особенностей биологии и экологии вида. Факторами, ограничивающими распространение данного вида, по нашему мнению, являются его стенотопность, а также резкие колебания численности и онтогенетической структуры популяций в зависимости от климатических условий разных лет.
Список литературы Oxytropis stukovii Palib. - редкий вид остролодочника Восточного Забайкалья
- Большой энциклопедический словарь. Биология. -М., 2001. -863 с.
- Губанов И. А. Конспект флоры Внешней Монголии (сосудистые растения) -М., 1996. -136 с.
- Заугольнова Л. Б. Методика изучения ценопопуляций редких видов растений с целью оценки их состояния//Охрана растительных сообществ редких и находящихся под угрозой исчезновения экосистем: материалы I Всесоюзн. конф. по охране редких растит. сообществ. -М., 1982. -С. 74-76.
- Игнатьев А. П. Рекомендуемые к охране уникальные растительные сообщества Читинской области//Исследования молодых ботаников Сибири: тез. докл. молод. конф. -Новосибирск, 2001. -С. 36-37.
- Красная книга Читинской области и Агинского Бурятского автономного округа. Растения. -Чита, 2002. -277 с.
- Макаров В.П. Береговые растительные сообщества озера Ножий (Восточное Забайкалье)//Социально-эколого-экономические проблемы развития приграничных регионов России -Китая -Монголии: материалы науч.-практ. конф. -Чита, 2010. -С. 62-67.
- Малышев Л. И., Пешкова Г. А. Особенности и генезис Флоры Сибири. Предбайкалье и Забайкалье. -Новосибирск, 1984. -264 с.
- Новороцкий П. В. Климатические изменения в бассейне Амура за последние 115 лет//Метеорология и гидрология. -2007. -№ 2. -С. 43-53.
- Обязов А. А. Изменения климата в Забайкалье//Материалы XIII науч. совещ. географов Сибири и Дальнего Востока. -Иркутск, 2007. -Т. 2. -С. 97-98.
- Обязов А. А. Адаптация к изменениям климата: региональный подход//География и природные ресурсы. -2010. -№. 2. -С. 34-39.
- Попов М. Г. Эндемизм во флоре побережий Байкала и его происхождение//Академику В. Н. Сукачеву к 75-летию со дня рождения. -М.; Л., 1956. -С. 442-462.
- Словарь ботанических терминов. -Киев, 1984. -306 с.
- Ценопопуляции растений (основные понятия и структура). -М., 1976. -215 с.
- Ценопопуляции растений (очерки популяционной биологии). -М., 1988. -184 с.
- Флора Центральной Сибири, 1979. -Т. 2. -1046 с.
