П-аминобензойная кислота стимулирует всхожесть семян, рост растений, фотосинтез и ассимиляцию азота у амаранта (Amaranthus L.)
")
Автор: Кириллова Л.Л., Назарова Г.Н., Иванова Е.П.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Фотосинтез и продуктивность
Статья в выпуске: 5 т.51, 2016 года.
Бесплатный доступ
В настоящее время накоплен огромный фактический материал относительно полезного действия п-аминобензойной кислоты (пАБК) на все элементы структуры урожая растений разных видов и хозяйственного назначения, описаны ее протекторные, репарационные, антимутагенные, антиоксидантные свойства. Однако в мировой литературе практически отсутствует информация о влиянии пАБК на растения амаранта. Овощной амарант - важная пищевая культура, перспективная для средней полосы России, в связи с чем вызывают интерес способы облегчения его интродукции, повышения продуктивности и пищевой ценности. С этой целью представляется возможным использовать пАБК - биологически активное экологически чистое природное соединение с широким спектром действия. В представленной работе впервые исследовано влияние пАБК на всхожесть семян, рост, развитие и продуктивность растений у овощной формы амаранта Amaranthus caudatus L. (сортообразец К173) и A. cruentus L. (сортообразец К185), а также на некоторые показатели фотосинтеза и ассимиляции азота у растений К173. Для оценки активности пАБК сравнивали действие этого соединения и синтетического цитокинина 6-бензиламино-пурина (6-БАП). Установлена зависимость всхожести семян амаранта от концентрации используемых для обработки растворов пАБК (от 10-9 М до 10-4 М). У К173 в варианте с 10-6 М пАБК всхожесть повышалась на 23±5 %, с 10-4 М пАБК - снижалась на 22±4 %. При прочих концентрациях достоверный эффект отсутствовал. У К185 эффекты оказались аналогичными. По характеру действия 6-БАП и пАБК были сходны. На стадии скрытого роста у К173 высота 15-суточных проростков превышала контроль на 10±5 %, масса - на 76±6 %, длина корня - на 133±17 %, у К185 эти показатели были выше, но незначительно. 6-БАП по эффекту оказался практически вдвое сильнее пАБК. В результате развитие растений ускорялось, что способствует большей выживаемости и адаптации. Стимулирующее действие пАБК на рост растений в высоту сохранялось в течение жизни, за исключением периода закладки генеративных органов (60-80-е сут) у обоих сортообразцов. На всех этапах онтогенеза обнаружили значительное (от +30±11 % до +85±10 %) увеличение массы растений в сравнении с контролем. Продуктивность биомассы листьев с растения под действием пАБК и 6-БАП у обоих сортообразцов на 115-е сут в опыте была в среднем на 50±11 % больше контрольной. Чистая продуктивность фотосинтеза листьев у обоих сортообразцов за 45-55 сут под действием пАБК и 6-БАП повышалась одинаково (в среднем на 30 %). В листьях 45-суточных растений К173 в результате предпосевной обработки семян пАБК отмечали увеличение активности нитратредуктазы на 37±4 % (в варианте с 6-БАП - на 85±1 %) и содержания суммарного белка на 10±3 % (для 6-БАП - на 38±14 %), за счет которого повышается пищевая ценность культуры. В результате воздействия пАБК скорость фотофосфорилирования в хлоропластах возрастала на 27±6 % (для 6-БАП - на 86±13 %), скорость транспорта электронов - на 32±6 % (для 6-БАП - на 35±14 %) в сравнении с контролем. Обсуждаются фитогормональные свойства пАБК. Ее применение может быть рекомендовано при возделывании овощного амаранта.
Амарант, п-аминобензойная кислота, фитогормоны, всхожесть семян, рост и развитие растений, скрытый рост, адаптация, продуктивность, хлоропласты, фотофосфорилирование, скорость транспорта электронов, содержание белка, усвоение азота, нитратредуктаза
Короткий адрес: https://sciup.org/142213972
IDR: 142213972 | УДК: 635:582.663.2:631.811.982:581.13/.14 | DOI: 10.15389/agrobiology.2016.5.688rus
Para-aminobenzoic acid stimulats seed germination, plant growth, development, photosynthesis and nitrogen assimilation in the amaranth (Amaranthus L.)
Currently protective, reparative, anti-mutagenic, antioxidant properties of para-aminobenzoic acid (pABA) are described. The huge number of the facts is saved up concerning its efficiency on all elements of yield structure of different plants species and household purposes. However, there is almost no information about the effect of pABA on amaranth plants in the world literature. Vegetable amaranth is a valuable food culture, promising to Central Russia. Therefore, we are especially interested in ways to facilitate its introduction, increase productivity, and nutritional value. To this end, it is possible to use pABA as environmentally friendly biologically active natural compound with a broad spectrum of action. In this work, we first investigated the effect of pABA on seed germination, and growth, development and productivity of vegetable amaranth Amaranthus caudatus L. variety K173 (K173) and A. cruentus L. variety K185 (K185) of grown plants as well as on light-dependent processes of photosynthesis and nitrogen assimilation in the leaves. To assess the characteristics and level of pABA activity the experimental results concerning influence of a synthetic citokinin 6-benzilaminopurin (6-BAP) on the same parameters are given. A dependence of amaranth seed germination on pABK 10-9 М to 10-4 М concentration used for treatment was found. The seed germination in both varieties was 23±5 % higher at 10-6 M pABK, whereas 10-4 M pABK decreased it by 22±4 %, and other concentrations had no significant effect. In K173 the 15-day seedlings exceeded the control by 10±5 % in height, by 76±6 % in weight, and by 133±17 % in root length at the latent growth stage. Parameters of K185 plants were slightly higher. The acceleration of plant development may contribute to survival and adaptation. Stimulating effect of pABA on К173 and К185 plant growth in height is maintained for the life of plants, except initiation of generative organs (days 60-80). A significant increase (30-85 %) in plant weight compared to control was identified at all stages of ontogenesis. Productivity of leaf biomass per plant in both varieties on day 115 was on average 50±11 % more than in the control. In the leaves of 45-day plants of К173 grown from treated seed, the activity of nitrate reductase increased by 37±4 % and the total protein level was 10±3 % higher when compared to control, thus improving nutritional value of the culture. The rate of photophosphorylation in chloroplasts was shown to be 27±6 % higher, and the rate of electron transport was 32±6 % higher in comparison with the control that led to an increase in leaf net photosynthesis by 22±7 %. These were no different from 6-BAP effects. The pABA influence on the seed germination and plant biometric parameters in K173 and K185 was comparable to the action of 6-BAP. pABK influenced nitrate reductase activity, protein level, chlorophyll content, and photophosphorylation rate in leaves of K173 plants much weaker, than 6-BAP, and differed from it. The pABA properties as a plant hormone are discussed, and the use is recommended for the vegetable amaranth cultivation.
Текст научной статьи П-аминобензойная кислота стимулирует всхожесть семян, рост растений, фотосинтез и ассимиляцию азота у амаранта (Amaranthus L.)
Первые сообщения о стимулирующем действии п-аминобензойной кислоты (пАБК) на растения были сделаны много десятилетий назад (1). В настоящее время установлены ее репарационные (2), протекторные (3), ан-тимутагенные (4), антиоксидантные (5), фитогормональные (6) свойства. Такое многообразие эффектов объясняют следствием того, что пАБК — компонент фолатов (7), важнейшего кофактора реакций переноса одноуглеродных единиц (8). Таким образом, остаток пАБК участвует во многих ключевых процессах: в биосинтезе пуринов и пиримидинов, пантотената, в синтезе S-аденозил-1-метионина (SAM), формил-метионина-тРНК, дезоксирибонуклеотидов, в превращениях аминокислот (9). Показана важная роль пАБК как компонента фолата в регуляции активности ДНК-полимеразы, ДНКазы и РНКазы (10, 11), транскрипции (12), при активировании фотофосфорилирования, окислительного фосфорилирования, фотодыхания (8) и 688
пр. Однако появляются факты, которые не вписываются в такую схему. Это касается, например, антиоксидантных (13) и некоторых протекторных свойств (14), обнаруженных у самой пАБК. Недавно установлено непосредственно участие пАБК в регуляции активности фермента окислительного стресса — пероксидазы (15). Показана способность пАБК вызывать у растений системную приобретенную устойчивость к заболеваниям (16). Возможно, и гормоноподобные свойства пАБК не исчерпываются ее участием в регуляции синтеза пуринов фолатами (8), но могут быть обусловлены особенностями ее молекулы (17).
Накоплен огромный фактический материал относительно полезного действия пАБК на все элементы структуры урожая растений разных видов и хозяйственного назначения: злаковые (ячмень, овес, кукуруза, пшеница), бобовые (горох, люпин), овощные (огурцы, томаты), хлопчатник, виноград и пр. (1, 2, 5, 6, 18-20). При этом в основном использовали предпосевную обработку семян. Среди обилия подобной информации нами обнаружено лишь единичное сообщение о применении пАБК на амаранте (21), где ее использовали «для фенотипической коррекции морфологических признаков». В результате опрыскивания соцветий амаранта повышалась масса семян на растении. Наиболее эффективной была концентрация пАБК 0,02 % (1,5½10-3 М). Мы предположили, что пАБК также будет оказывать регуляторное действие на растения амаранта при обработке семян.
Наш интерес к овощному амаранту обусловлен тем, что эта важная пищевая культура с высоким содержанием сбалансированного по составу белка весьма перспективна для средней полосы России (22). Для облегчения его интродукции, повышения продуктивности и пищевой ценности представляется возможным использовать пАБК — биологически активное соединение с широким спектром действия, экологически чистое вследствие его природного происхождения. Результаты такого использования пАБК могли бы пополнить недостаток сведений об ее влиянии на фотосинтетические и ассимиляционные процессы.
Целью работы было изучение действия п-аминобензойной кислоты (пАБК) на семена и выращенные из них растения овощного амаранта двух видов и сортообразцов, что позволило оценить возможность применения этого соединения при культивировании амаранта в средней полосе России. Задачи заключались в определении ростовых параметров растений разных сортообразцов в течение всего онтогенеза, а также в оценке активности ассимиляции азота и фотосинтетического аппарата по параметрам световой стадии фотосинтеза у одного из сортообразцов. В проведенных экспериментах для оценки величины и характера активности пАБК ее сравнивали с действием синтетического цитокинина 6-бензиламинопурина (6-БАП).
Методика . В работе использовали семена овощных форм амаранта Amaranthus caudatus L . сортообразца К173 (К173) и A. cruentus L . сортооб-разца К185 (К185), полученные из Всероссийского НИИ селекции и семеноводства овощных культур (Московская обл.) и кристаллические пАБК и 6-БАП («Sigma-Aldrich Corp.», США).
Для определения влияния пАБК и 6-БАП на всхожесть семена амаранта замачивали в водных растворах препаратов (от 10-9 до 10-4 М) на 24 ч. Затем семена высушивали в слабом токе воздуха при комнатной температуре. В качестве контроля использовали семена, замоченные дистиллированной воде. Всхожесть определяли по проценту проросших семян от их общего числа после инкубации на влажной фильтровальной бумаге в чашках Петри в течение 72 ч при 24 ° С.
Для изучения ростовых и биохимических показателей растения вы- ращивали из семян, обработанных 1 мкМ растворами пАБК или 6-БАП (опыт) или дистиллированной водой (контроль), как описано выше. Пророщенные семена высаживали в сосуды с песком (10 откалиброванных проростков на сосуд, по 3 сосуда на каждый вариант эксперимента) и выращивали растения при температуре 24 °С, 14-часовом фотопериоде и освещенности 150 Вт•м-2; для полива использовали питательную среду Кноппа. Биометрические показатели учитывали каждые 15 сут до даты сбора урожая (115-е сут), о продуктивности судили по приросту биомассы. В листьях 45-суточных растений определяли содержание хлорофилла (23); фотохимическую активность хлоропластов, изолированных (24) из таких листьев, оценивали по скорости электронного транспорта (25) и фотофосфорилирования (26). В листьях растений с 15-х по 45-е сут измеряли активность нитратредук-тазы (НР) и количество нитрита (27), содержание элементарного азота (28) и общего белка (29). Чистую продуктивность фотосинтеза (ЧПФ, г^м-2^сут-1) за период с 45-х по 55-е сут рассчитывали по методу Ничипоровича (30).
В статье представлены результаты одного типичного эксперимента из пяти. Биометрические параметры определяли у 30 растений. Повторность биохимических анализов 3-кратная. В таблицах и на рисунках приведены средние арифметические значения со стандартной ошибкой. Достоверность различий оценивали по t -критерию Стьюдента при Р = 0,95.
Результаты . Всхожесть семян амаранта зависела от концентрации растворов пАБК и 6-БАП, используемых для обработки (рис. 1). У необработанных семян она составляла 70 %. Воздействие пАБК приводило к ее значительному повышению (на 23±5 %) при концентрации 10-6 М и снижению на 22±6 % — при 10-4 М. Действие 6-БАП было аналогичным. Поскольку оптимальной для всхожести семян оказалась концентрация растворов 10-6 М (1 мкМ), ее далее использовали в экспериментах с растениями. Данные по всхожести семян у К185 и К173 почти не различались.
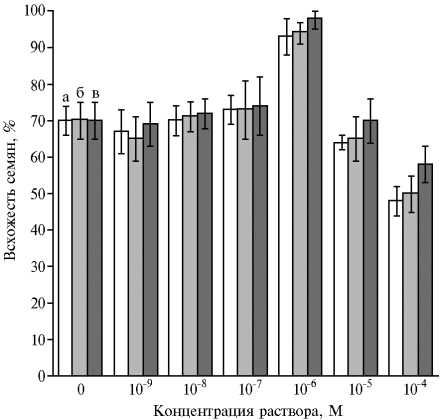
Рис. 1. Всхожесть семян у сортообразцов овощного амаранта Amaranthus caudatus L. К173 (а) и A. cruentus L . К185 (б) при обработке растворами п-аминобензойной кислоты и A. caudatus L . К173 при обработке 6-бензиламинопурином (в) ( Х±х , лабораторный опыт).
Мы обнаружили, что после предпосевной обработки семян 1 мкМ пАБК 15-суточные проростки К173 и К185 превосходили контроль по высоте на 10±5 % (с 6-БАП на 22±4 %), по массе на 76±6 % (с 6-БАП на 129±8 %) (рис. 2). Обычно в этот период развития у растений ама- ранта наступает остановка роста надземной части, называемая фазой скрытого роста. Из приведенных данных видно, что проростки миновали указанную фазу при воздействии обоих стимуляторов, но эффект 6-БАП был сильнее. Необходимо отметить, что при этом наблюдали значительное ускорение роста корней. Длина главного корня у контрольных растений составляла в среднем 1,2±0,1 см, после воздействия пАБК — 2,8±0,3 см, 6-БАП — 3,3±0,3 см, или соответственно 233±10 % и 275±9 % к контролю. Влияние 6-БАП на все показатели у 15-суточных проростков оказалось заметно сильнее, чем у пАБК. Результаты 690
воздействия стимуляторов на параметры проростков К185 были аналогичны таковым для К173.
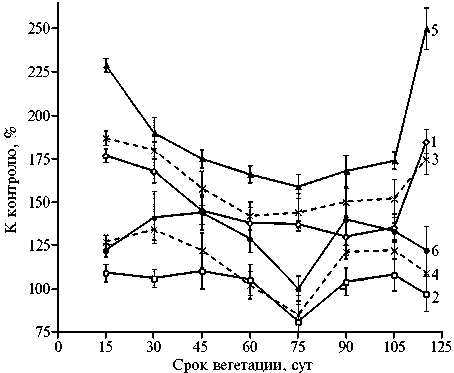
Рис. 2. Динамика ростовых показателей у двух сортообразцов овощного амаранта после предпосевной обработки семян раствором п-аминобен-зойной кислоты (пАБК, 1 мкМ) и у одного сортообразца при воздействии 6-бензиламинопурина (6-БАП, 1 мкМ) 1, 2 — Amaranthus caudatus L. К173, 3, 4, 5, 6 — A. cruentus L . К185; 1, 3, 5 — накопление биомассы, 2, 4, 6 — высота растений; 1, 2, 3, 4 — пАБК, 5, 6 — 6-БАП ( X±x , лабораторный опыт).
Стимулирующее действие пАБК на рост в высоту сохранялось на протяжении жизни растений, за ис- ключением периода закладки генеративных органов (60-80-е сут) (рис. 2). Было обнаружено достоверное значительное увеличение массы целых растений на всех этапах онтогенеза в сравнении с контролем. Максимальный эффект наблюдали к сбору урожая (115-е сут), когда масса надземной части растений, выращенных из обработанных пАБК семян, превышала массу контрольных на 85±10 %. Характер воздействия пАБК и 6-БАП на высоту и массу растений в онтогенезе совпадал (см. рис. 2), но степень влияния последнего была выше. Тенденции в изменении ростовых параметров у второго сортообразца К185 при применении регуляторов роста полностью воспроизводили их динамику для К173, но при этом абсолютные значения оказались несколько выше.
Основной показатель продуктивности овощных форм амаранта — выход биомассы листьев с растения. Его значение на 115-е сут при воздействии пАБК на К173 составляло в среднем в контроле 130±10 г, в опыте — 195±14 г (на 50±11 % больше, чем в контроле). Этот параметр для К185 превышал контроль на 68±9 % (95±6 г в контроле и 160±14 г в опыте). Масса 1 м2 листьев у растений К173 при предпосевной обработке семян превышала контроль на 48±5 % (520±26 г в контроле и 770±36 г в опыте), у К185 — на 40±8 %. 6-БАП увеличивал выход биомассы листьев с растения на 62±9 %, а массу 1 м2 листьев — на 61±8 %. Эти данные свидетельствуют, что степень воздействия пАБК и 6-БАП на продуктивность листовой массы у двух сортообразцов достоверно не различалась или различалась мало.
Поскольку продуктивность растений во многом определяется активностью фотосинтетического аппарата, мы изучили фотохимическую активность изолированных хлоропластов. В хлоропластах из листьев 45-суточных растений сортообразца К173 при предпосевной обработке семян пАБК и 6-БАП отмечалось увеличение как скорости транспорта электронов, так и скорости образования АТФ в сравнении с контролем (табл. 1). При этом степень воздействия у 6-БАП на скорость образования АТФ была значительно выше, чем у пАБК при практически равном влиянии на скорость транспорта электронов. Различие в эффекте двух биоактивных агентов заключалось и в том, что при применении пАБК содержание хлорофилла в листьях не изменялось, а под воздействием 6-БАП сильно увеличивалось. Несмотря на это, значения ЧПФ листьев с 45-х по 55-е сут роста в вариантах с пАБК и 6-БАП достоверно не различались: с пАБК ЧПФ увеличилась на 22±7 % относительно контроля (с 2,3±0,15 до 2,8±0,19 г•м-2•сут-1), с 6-БАП — на 30±5 % (с 2,3±0,15 до 3,0±0,15 г•м-2•сут-1).
1. Показатели фотосинтетической активности в хлоропластах из листьев 45-суточных растений амаранта ( Amaranthus caudatus L., сортообразец К173) после предпосевной обработки семян пАБК и 6-БАП ( X±x , лабораторный опыт)
|
Вариант |
Содержание хлорофилла |
Скорость |
|
|
транспорта электронов |
фотофосфорилирования |
||
|
мг/г сухой массы к контролю, % |
мкМ K 3 [Fe(CN 6 )]1к контролю, % |
мкМ АТФ к контролю, % |
|
|
Контроль |
9,9±0,2 |
100 |
110,4±8,0 |
100 |
112,0±2,3 |
100 |
|
пАБК |
9,9±0,6 |
100±6 |
145,9±7,0 |
132±6 |
142,0±2,0 |
127±6 |
|
6-БАП |
14,4±2,2 |
146±15 |
148,5±15,0 |
135±14 |
209,0±15,0 |
186±13 |
П р и м еч а ни е. пАБК — п-аминобензойная кислота, 6-БАП — 6-бензиламинопурин. Скорость указана в расчете на 1 мг хлорофилла за 1 ч. Концентрация растворов пАБК и 6-БАП — 1 мкМ.
Применение как пАБК, так и 6-БАП (1 мкМ) приводило к увеличению активности НР в сравнении с контролем в течение первых 45 сут вегетации, обеспечивая повышенный уровень синтеза азотсодержащих веществ в период активного роста и их расходования на пластические процессы (табл. 2). Однако характер действия двух стимуляторов различался: в варианте с пАБК активность НР стабильно превышала контроль в среднем на 34 %, с 6-БАП — увеличивалась постепенно по мере роста растений, при этом у 6-БАП эффект был значительно сильнее, чем у пАБК.
Обработка как пАБК, так и 6-БАП вызывала повышение содержания тотального белка в листьях растений амаранта К173 на протяжении вегетации с 15 по 45 сут. В процессе роста наблюдали тенденцию к снижению содержания общего белка при воздействии обоих биоактивных агентов, но в случае 6-БАП к 45-м сут этот показатель оставался заметно выше. При действии пАБК (в отличие от 6-БАП) накопления нитрита листьями не наблюдали. Не всегда активность нитратредуктазы, процентное содержание элементарного азота и количество общего белка в растениях прямо взаимосвязаны (31). Однако факт достоверного стимулирующего воздействия пАБК на ассимиляцию и превращение азота в растениях амаранта нами установлен (см. табл. 2).
2. Изменение показателей азотного обмена (к контролю, %) в листьях растений амаранта ( Amaranthus caudatus L., сортообразец К173) после предпосевной обработки семян пАБК и БАП ( X±x , лабораторный опыт)
|
Вариант |
Возраст растений, сут |
Нитрит |
Активность нитратредуктазы |
Общий |
|
|
азот |
белок |
||||
|
пАБК |
15 |
143±5 |
130±2 |
104,0±0,5 |
128±9 |
|
30 |
101±2 |
134±2 |
102,0±1,1 |
121±4 |
|
|
45 |
108±3 |
137±4 |
102,0±0,7 |
110±6 |
|
|
6-БАП |
15 |
124±4 |
147±9 |
108,0±0,4 |
142±9 |
|
30 |
109±2 |
167±12 |
105,0±0,5 |
137±9 |
|
|
45 |
122±5 |
185±18 |
106,0±1,2 |
138±14 |
|
П р и м еч а ни е. пАБК — п-аминобензойная кислота, 6-БАП — 6-бензиламинопурин. Концентрация растворов пАБК и 6-БАП — 1 мкМ.
Выявленное влияние предпосевной обработки семян пАБК на хозяйственно значимые характеристики амаранта обусловлено воздействием на фундаментальные физиолого-биохимические процессы. Впервые показано стимулирование световых реакций фотосинтеза и скорости усвоения азота у сортообразца К173 при применении пАБК. Необходимо отметить, что характер и степень проявления некоторых изученных эффектов у пАБК и 6-БАП различался, следовательно, пАБК действует не как цитокинин. Вероятно, повышенное содержание пАБК как предшественника фолатов стимулирует их синтез и накопление уже на начальных этапах развития растений из семени, ускоряя физиолого-биохимические процессы в течение всего онтогенеза и приводя к наблюдаемым эффектам.
Однако возможен и иной механизм: имеются, в частности, скудные сведения о цитокиновых (1, 6) и ауксиновых (17, 32) эффектах пАБК. Хотя эта область практически не исследована, нет оснований отрицать наличие у пАБК свойств собственно растительных гормонов — способности через систему мессенджеров экспрессировать ядерный ответ, как это описано, например, для цитокининов (33). Также не исключено взаимодействие с рецепторами фитогормонов (18). Обнаруженные нами эффекты пАБК указывают в пользу такого предположения, если исходить из общих признаков гормонов растений согласно International Plant Growth Substance Association (IPGSA, Международная ассоциация растительных ростовых веществ). По IPGSA, фитогормоны — природные органические соединения, влияющие на физиологические процессы (рост, дифференциацию, развитие растения и др.) в значительно меньших концентрациях, чем питательные вещества или витамины (34). Выявленные нами эффекты нельзя определить как типичные для какого-либо известного класса, а для утверждения о новом классе фитогормонов имеющегося фактического материала недостаточно. Мы согласны с предположением о гормональной активности некоторых малых молекул (35). Более того, мы описали цитокинин-подобные свойства 4-гидроксифенэтилового спирта и сделали предположение о его гормональной природе (36). Основанием послужил прецедент о-гидрокси-бензойной (салициловой) кислоты. В настоящее время эта малая молекула и несколько ее производных признана особым классом фитогормонов. Возможно, при накоплении данных о малых молекулах проявятся такие общие характеристики, которые позволят объединить их в новую группу соединений со специфическими фитогормональными свойствами.
Итак, предпосевная обработка семян овощного амаранта 1 мкМ водным раствором п-аминобензойной кислоты (пАБК) повышает всхожесть и улучшает биометрические и физиолого-биохимические показатели растений: ускоряет развитие проростков в фазу скрытого роста, усиливает рост, повышает продуктивность и пищевую ценность. По эффекту пАБК и цитокинин 6-бензиламнопурин не аналогичны. пАБК можно рекомендовать при выращивании овощного амаранта в Нечерноземной зоне России.
Список литературы П-аминобензойная кислота стимулирует всхожесть семян, рост растений, фотосинтез и ассимиляцию азота у амаранта (Amaranthus L.)
- Macht D.I. Influence of sulfonamides and para-aminobenzoic acid on the growth of Lupinus albus. Exp. Biol. Med., 1945, 60: 217-220 ( ) DOI: 10.3181/00379727-60-15140
- Рапопорт И.А., Васильева С.В., Давниченко Л.С. Роль пара-аминобензойной кислоты в репарации повреждений, индуцированных УФ-и γ-излучениями. ДАН, 1979, 247(1): 231-234.
- Tashenova A.A., Kulumbetova S.K., Sinel'shchikova T.A., Akhmatullina N.B., Zasukhina G.D., Rapoport I.A. Treatment of cells with para-aminobenzoic acid reduces the number of DNA breaks induced by chemical mutagens. Dokl. Akad. Nauk SSSR, 1990, 311(4): 977-979.
- Gichner T., Badaev S.A., Pospísil F., Velemínský J. Effects of humic acids, para-aminobenzoic acid and ascorbic acid on the N-nitrosation of the carbamate insecticide propoxur and on the mutagenicity of nitrosopropoxur. Mutat. Res., 1990, 229(1): 37-41 ( ) DOI: 10.1016/0027-5107(90)90006-P
- Эйгес Н.С., Волченко Г.А., Волченко С.Г., Вайсфельд Л.И., Козлов В.С., Донец Н.В. Некоторые аспекты ненаследственной изменчивости, индуцированной на сельскохозяйственных культурах с помощью антиоксиданта пара-аминобензойной кислоты. Автохтоннi та iнтродукованi рослини, 2012, 8: 71-78.
- Cronenberg L., Abelle T., Pacheco H. New synthetic substances with phytohormonal properties. Bull. Soc. Chim. Biol., 1964, 46: 703-715.
- Basset G.J., Quinlivan E.P., Ravanel S., Rebeille F., Nichols B.P., Shinozaki K., Seki M., Adams-Phillips L.C., Giovannoni J.J., Gregory J.F. III, Hanson A.D. Folate synthesis in plants: the p-aminobenzoate branch is initiated by a bifunctional PabA-PabB protein that is targeted to plastids. PNAS USA, 2004, 101: 1496-1501 ( ) DOI: 10.1073/pnas.0308331100
- Rebeille F., Ravanel S., Jabrin S., Douce R., Storozhenko S., van der Straeten D. Folates in plants: biosynthesis, distribution, and enhancement. Physiologia Рlаntarum, 2006, 126: 330-342 ( ) DOI: 10.1111/j.1399-3054.2006.00587.x
- Hanson A.D., Roje S. One-carbon metabolism in higher plants. Annu. Rev. Plant Biol., 2001, 52: 119-137 ( ) DOI: 10.1146/annurev.arplant.52.1.119
- Kozhevnikova N.A., Rapoport I.A. Ability of para-aminobenzoic acid to restore the activity of alkaline ribonuclease. Izv. Akad. Nauk SSSR Biol., 1987, 5: 787-791.
- Кожевникова Н.А., Рапопорт И.А., Иваницкая Е.А., Пудрина И.Д. Влияние пара-аминобензойной кислоты на активность дезоксирибонуклеазы интактного и облученного препарата. ДАН, 1983, 273(2): 476-479.
- Trausch J.J., Batey R.T. A disconnect between high-affinity binding and efficient regulation by antifolates and purines in the tetrahydrofolate riboswitch. Chem. Biol., 2014, 21(2): 205-216 ( ) DOI: 10.1016/j.chembiol.2013.11.012
- Sochor J., Zitka O., Skutkova H., Pavlik D., Babula P., Krska B., Horna A., Adam V., Provaznik I., Kizek R. Content of phenolic compounds and antioxidant capacity in fruits of apricot genotypes. Molecules, 2010, 15(9): 6285-6305 ( ) DOI: 10.3390/molecules15096285
- Мостовщикова С.М., Белозерова А.А. Оценка влияния пара-аминобензойной кислоты на морфометрические параметры растений пшеницы (Titicum aestivum L.) в условиях хлоридного засоления. Успехи современного естествознания, 2013, 8: 20-21.
- Pozdnyakova N., Makarov O., Chernyshova M., Turkovskaya O., Jarosz-Wilkolazka A. Versatile peroxidase of Bjerkandera fumosa: substrate and inhibitor specificity. Enzyme Microb. Technol., 2013, 52(1): 44-53 ( ) DOI: 10.1016/j.enzmictec.2012.10.005
- Song G.C., Choi H.K., Ryu C.M. The folate precursor para-aminobenzoic acid elicits induced resistance against Cucumber mosaic virus and Xanthomonas axonopodis. Ann. Bot., 2013, 111(5): 925-934 ( ) DOI: 10.1093/aob/mct049
- Crisan M.E., Bourosh P., Maffei M.E., Forni A., Pieraccini S., Sironi M. Chumakov Y.M. Synthesis, crystal structure and biological activity of 2-hydroxyethylammonium salt of p-aminobenzoic acid. PLoS One, 2014, 9(7) ( ) DOI: 10.1371/journal.pone.0101892
- Солоненко Ю.А., Алексейчук Г.Н. Влияние пара-аминобензойной кислоты на прорастание семян и развитие проростков Glycine max L. и Lupinus angustifolius L. Весцi Нацыянальнай акадэмii навук Беларусi, 2005, 5(2): 63-66.
- Эйгес Н.С. Активация фенотипа с помощью пАБК. В сб.: Химические мутагены и пара-аминобензойная кислота в повышении урожайности сельскохозяйственных растений. М., 1989.
- Шерер В.А., Гадиев Р.Ш. Применение регуляторов роста в виноградарстве и питомниководстве. Киев, 1991.
- Боме Н.А., Говорухина А.А. Эффективность влияния пара-аминобензойной кислоты на онтогенез растений в условиях стресса. Вестник Тюменского государственного университета, 1998, 2: 176-182.
- Кононков П.Ф., Пивоваров В.Ф., Гинс В.К., Гинс М.С. Интродукция и селекция овощных культур для создания нового поколения продуктов функционального действия. М., 2008.
- Wintermans J.F.G.M., De Mots A. Spectrophotometric characteristics of chlorophyll a and b and their pheophytins in ethanol. Biochеm. Biophys. Acta, 1965, 109(2): 448-453.
- West K.R., Wiskich J.T. Photosynthetic control by isolated pea chloroplasts. Biochem. J., 1968, 109(4): 527-532 ( ) DOI: 10.1042/bj1090527
- Izava S., Good N.E. Hill reaction rates and chloroplasts fragment size. Biochem. Biophys. Acta, 1965, 109: 373-381.
- Bjorkmann O.М., Boardman N.K., Anderson J.M. Chloroplast components and Structure. Carnegie Inst. Year Book, 1972, 71: 115-135.
- Hageman R.H., Reed A.J. Nitrate reductase from higher plants. Methods in Enzymology, 1980, 69: 270-280 ( ) DOI: 10.1016/S0076-6879(80)69026-0
- Smart M.M., Rada R.G., Donnermeyer G.N. Determination of total nitrogen in sediments using persulfate digestion an evaluation and comparison with Kjeldahl procedure. Water Res., 1983, 17(9): 1207-1211 ( ) DOI: 10.1016/0043-1354(83)90063-5
- Bredford A. Rapid and sensitive gram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, 1976, 72: 248-254.
- Ничипорович А.А. Фотосинтетическая деятельность растений как основа их продуктивности в биосфере и земледелии. М., 1988.
- Cheeseman J.M., Tankou S.K. Nitrate reductase and growth of Arabidopsis thaliana in solution culture. Plant and Soil, 2005, 266: 143-154 ( ) DOI: 10.1007/s11104-005-0947-1
- Crisan S.F., Garay A.S. Influence of p-aminobenzoic acid on the activity of indoleacetic acid oxidase. Experientia, 1968, 15(24-10): 1009-1010.
- Hutchison C.E., Kieber J.J. Signaling via histidine-containing phosphotransfer proteins in Arabidopsis. Plant Signal Behav., 2007, 2(4): 287-289.
- Plant hormones: biosynthesis, signal transduction, action!/P.J. Davies (ed.). London, 2010.
- Bektas Y., Eulgem T. Synthetic plant defense elicitors. Front. Plant Sci., 2015, 5: 1-17 ( ) DOI: 10.3389/fpls.2014.00804
- Иванова Е.П., Кириллова Л.Л., Смолыгина Л.Д., Сердюк О.П. Применение перспективного природного стимулятора роста 4-гидроксифенэтилового спирта для улучшения качества посевного материала и продуктивности амаранта. Сельскохозяйственная биология, 2011, 5: 118-122.
