Палеоэкологические и палеобиогеографические особенности в эволюции табулят в позднем ордовике и раннем силуре на севере Урала

Автор: Лукин В.Ю.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 3 (291), 2019 года.
Бесплатный доступ
В статье сделан анализ палеоэкологических и палеобиогеографических закономерностей развития табулят в позднем ордовике и раннем силуре на севере Урала. К настоящему времени отсутствуют работы, в которых получили бы целостную характеристику таксономический состав, стратиграфическое и географическое распространение, а также эволюционное развитие табулят ордовика и силура севера Урала. В связи с этим представляется важным исследование временного и пространственного распространения табулят, необходимое для более полного использования этой группы бентоса в практических целях.
Табуляты, ордовик, силур, палеобиогеография, палеоэкология
Короткий адрес: https://sciup.org/149129363
IDR: 149129363 | УДК: 563.67: | DOI: 10.19110/2221-1381-2019-03-24-30
Paleoecological and paleobiogeographical regularities in the evolution of tabulates in late Ordovician and early Silurian in the north of the Urals
The paper analyzes paleoecological and paleobiogeographic regularities of tabulation evolution in the Late Ordovician and Early Silurian in the north of the Urals. By now there are no works completely describing the taxonomic composition, stratigraphic and geographical distribution and evolutionary development of Ordovician and Silurian tabulates of the north of the Urals. It is important to study the temporal and spatial distribution of tabulates, which is necessary for a more complete use of this group of benthos for practical purposes.
Текст научной статьи Палеоэкологические и палеобиогеографические особенности в эволюции табулят в позднем ордовике и раннем силуре на севере Урала
Коралловая фауна палеозоя севера Урала изучена недостаточно. До настоящего времени отсутствуют монографические работы по табулятам этой территории. Первые попытки создания биостратиграфических схем по ордовику и силуру были предприняты еще в прошлом веке. Однако терригенно-карбонатные отложения ордовика и силура, содержащие бентосную фауну, в том числе табуляты, широко распространенные на севере Урала и в сопредельных регионах, не могут быть расчленены и скоррелированы без изучения основных групп бентоса. В этом и заключается актуальность и практическая необходимость проведенных исследований.
Методы исследования
За основу были взяты стандартные методы исследования коралловой фауны — определение до вида или рода (зависит от сохранности ископаемой фауны) под бинокуляром, а также визуальные наблюдения при изучении и описании естественных обнажений ордовика и силура в бассейне р. Кожым (Приполярный Урал) и нар. Лек-Елец (Полярный Урал). Также были привлечены опубликованные в разные годы материалы.
История исследований
Первые сведения о фауне кораллов севера Урала содержатся в работах Р. И. Мурчисона и Ф. Н. Чернышева. Описание четырех новых видов из среднего ордовика Приполярного Урала приведены в монографии Б. С. Со колова. В работах многих геологов, занимавшихся стратиграфическими исследованиями на западном склоне севера Урала, — Г. А. Чернова, В. А. Варсанофьевой, А. И. Першиной, Б. Я. Дембовского, В. С. Цыганко и др. — приводятся лишь списки комплексов табулят. В трудах этих авторов использовались видовые определения, сделанные в свое время И. И. Чудиновой, В. Д. Чехович, В. Ф. Барской, М. А. Смирновой, Н. А. Боринцевой, Л. И. Лобурце-вой. В последнее время исследованием биостратиграфии и биоразнообразия табулят занимается автор данной статьи. В итоге по публикациям коллег и собственным материалам в ордовикских и силурийских отложениях Североуральского палеобассейна выявлен богатый и разнообразный комплекс табулят, имеющий большое биостратиг-рафическое, палеонтологическое и корреляционное значение [11]. Список наиболее распространенных североуральских табулят приведен в таблице.
Основные результаты
Территория Североуральского палеобассейна в раннем палеозое находилась в зоне тропического и субтропического климата, что благоприятствовало развитию и распространению морской теплолюбивой фауны. Существование и расселение табулятовых сообществ было связано непосредственно с историей развития бассейна и обусловлено наличием обширных мелководных областей.
Геологическое развитие территории Североуральского сегмента в конце ордовика и в силуре в значительной мере повторяет основные моменты глобальной истории этого периода [1]. Табуляты были обитателями преимущественно отмельной зоны, поэтому они чаще, чем население более глубоких частей моря, подвергались воздействию кратковременных изменений береговой линии.
В Североуральском морском палеобассейне одна из наиболее крупных вспышек биоразнообразия большинства бентосных организмов-фильтраторов — целентерат, мшанок — произошла в среднеордовикское время, когда были широко распространены табуляты семейств Lichenariidae, Billingsariidae, Lyoporidae, Tetradiidae. Массовое развитие получили представители рода Lichenaria. Кроме лихенарий здесь были распространены роды Lessnikovaea, Calapoecia, Nyctopora, Eofletcheria, Rhabdotetradium, Paratetradium и другие, мигрировавшие из Североамериканского палеобассейна через палеобассейны арктических островов [18].
В позднеордовикское кырьинское (катийское) время в Североуральском палеобассейне сложились благоприятные условия для обитания хализитид: Catenipora sp., Eocatenipora parallela (Shmidti), Eocatenipora sp.; фавози-тид (рис. 1): Favosites gotlandicus Sokolov, Palaeofavosites borealis Tchern, Mesofavosites aff. obliquus Sok. и гелиоли-тид (см. таблицу). Для табулят, ругоз, брахиопод в это время был характерен гигантизм [6]. Колониальные постройки у некоторых кораллов достигали более 50 см в диаметре. Доминантами табулятовых ассоциаций оставались представители рода Catenipora [17, 18].
Биогеографическое сходство североуральской ордовикской бентосной фауны, в том числе коралловой, с сибирской, казахстанской, алтайской и прибалтийской позволяет предположить существование в это время широких палеогеографических связей [3, 11, 12]. При этом следует отметить, что североуральская коралловая фауна обнаруживает наибольшее сходство с североамериканской.
Важная палеогеографическая особенность позднеордовикского времени — появление органогенных построек, образование рифовых массивов на северо- востоке палеобассейна [8, 9, 16]. Основными каркасостроителями рифов были водоросли, губки и колонии табулят родов Lichenaria, Saffordophyllum, Palaeofavosites, Mesofavosites, цепочечные кораллы рода Catenipora и гелиолитиды родов Plasmoporella, Acdalopora и Heliolites. (см. таблицу). Кораллы вместе с мшанками, криноидеями и водорослями формировали биогермы. Впадины между массивными колониями табулят заполняли крупные онколиты [9].
В конце кырьинского (катийского) времени ордовикские табуляты прекратили свое существование, как и другие бентосные ассоциации позднего ордовика, они были полностью замещены водорослевыми сообществами в юн-кошорское (хирнантское) время [6]. Обширная регрессия морского бассейна в это время привела к резкому сокращению площади эпиконтинентального шельфа. Это событие в конце ордовика многие исследователи связывают с крупным материковым оледенением африканской и южноамериканской частей Гондваны. Похолодание климата вызвало дисбаланс в шельфовых сообществах биоты, что проявилось в массовом вымирании фауны, в том числе и табулят, в большинстве морских бассейнов мира [20].
Обмеление моря в позднеордовикское время сменилось постепенно развивавшейся трансгрессией в начале лландовери. Морская лландоверийская трансгрессия в Североуральском палеобассейне достигла наибольшего распространения и вновь создала благоприятные условия для расселения бентосной фауны. Освободившиеся ранее
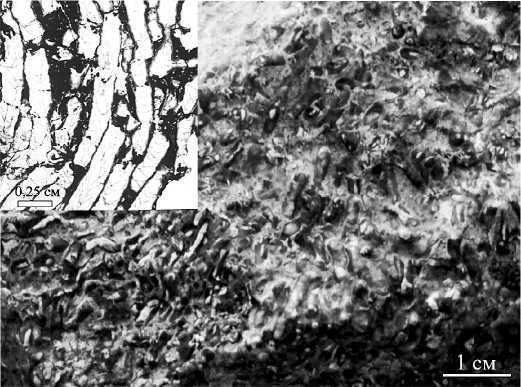
Рис. 1. Aulocystellaaseptata (Barskaja). Венлок. Руч. Безымянный, поднятие Чернова
Fig. 1. Aulocystella aseptata (Barskaja). The Wenlockian, Brook Bezymyannjy. Chernov Swell экологические ниши постепенно осваивали табуляты рода Catenipora, потомки ордовикских таксонов, которые пережили глобальный позднеордовикский кризис и широко расселились в палеобассейне.
Впервые в раннем лландовери зафиксированы представители сирингопорид (Syringopora), установленные в разрезе западного склона Приполярного Урала. В яренейс-кое и лолашорское время (руддан, аэрон) появились виды родов Favosites и Multisolenia. Хализитиды были представлены родами Catenipora и Halysites, существовавшими еще в ордовике [2, 6, 17]. Из гелиолитид сохранился только род Propora и появились виды рода Stelliporella. В среднем и позднем лландовери (в филиппъельское и маршрутнинс-кое время) наибольший расцвет получили фавозитиды родов Mesofavosites, Palaeofavosites [2, 6, 19]. В целом табуляты раннего силура характеризуются широким географическим распространением. Сходные комплексы выявлены в лландоверийских разрезах Сибири, Таймыра, островов Новая Земля, Долгий и Балтии, что подтверждает наличие широких связей между древними палеобассейнами в лландоверийское время, которые способствовали интенсивному обмену фауной.
В филиппъельское время систематический состав та-булятовой фауны Североуральского бассейна пополнился видами семейств фавозитид, альвеолитид и пахипарид (см. таблицу). В изученных автором разрезах Приполярного Урала табуляты представлены цепочечными Halysitida, первыми алвеолитидами (Subalveolites) и видами рода Multisоlenia (Favositida). Развитие табулят было связано с мелководными обстановками сублиторали восточной части палеобассейна, в которой обитали табуляты родов Mesofavosites, Multisolenia, Favosites, реликтовые формы ка-тенипор (Catenipora panga Klaаmаnn [2, 6, 19] и появившиеся впервые сирингопориды — Syringopora borealis Tchern. Сходные комплексы табулят известны в позднем лландовери и в бассейнах Балтии и Сибирской платформы [11].
В позднем лландовери (маршрутнинское время) трансгрессия морского бассейна вновь благоприятствовала широкому расселению бентосных организмов на территории бассейна. При этом родовой состав табулят не претерпел существенных изменений, а видовое разнооб-
Распространение табулят в позднем ордовике и раннем силуре в Североуральском палеобассейне
Distribution of tabulates in Late Ordovician and Early Silurian in Northern Ural paleobasin
|
Система / System Отдел / подотдел Division / subdivision |
Ордовикская / Ordovician Верхний / Upper |
Силурийская / Silurian Нижний / Lower |
||||||||
|
Лландоверийский Llandoverian |
Венлокский Wenlockian |
|||||||||
|
Ярус / Stage [21] |
Сандбийский Sandbian |
)S ед ед i^ |
ед X tf s fi X |
>s |
:S § 2 |
s ™ 5S |
Шейнвудский Гомерский Sheinwoodian Homerian |
|||
|
Региональные горизонты Regional horizons \ [5,6,15] Семейство/род/вид \ Family/Genus/Species |
a. |
)S 8 8 Я c |
IS s s § &^ Q |
ед s ^ |
a, S О Л a о Q ^ s § |
ед к с У cd ед я |
ед 5 S ч 'о |
s s ^ e |
ед S 5 |
Войвывский Voyvyvian |
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
|
|
Lichenariidae |
||||||||||
|
Lichenaria arctica Sokolov |
4 |
|||||||||
|
L. borealis Sokolov. |
+ |
|||||||||
|
L. expressa Sokolov |
4 |
|||||||||
|
L. markiitHr i Sokolov |
4 |
|||||||||
|
L. cf. carterensis (Saff.) |
4 |
|||||||||
|
Lessnikovaea elegans Sokolov |
4 |
|||||||||
|
L. spinosa Sokolov |
4 |
|||||||||
|
Billingsariidae |
||||||||||
|
Nyctopora billingsi Nicholson |
4 |
|||||||||
|
Saffordophyllum tulaensis (Sokolov) |
4 |
|||||||||
|
Lyoporidae |
||||||||||
|
Lyopora coxi (Bassler) |
||||||||||
|
Eotletcheria orvikui (Sokolov.) |
+ |
+ |
||||||||
|
Tetradiidae |
||||||||||
|
Rhabdotetradium sp. |
+ |
|||||||||
|
Halysitidac |
||||||||||
|
Catenipora rubra Sinclair et Bolton |
||||||||||
|
C. arctica Tchernyshev |
+ |
+ |
||||||||
|
C. copulataformis Barskaja |
4- |
|||||||||
|
C. escharoides Lam. |
4- |
|||||||||
|
C. gothlandica (Yabe) |
4- |
4 |
||||||||
|
C. maxima (F.-B.) |
4- |
|||||||||
|
C. panga Klaamann |
+ |
|||||||||
|
C. subgotlandicus Barskaja |
4 |
|||||||||
|
C. tarejaensis (Tchernyshev) |
4- |
|||||||||
|
C. sp. |
4 |
|||||||||
|
Eocatenipora parallels (Shmidti) |
4- |
|||||||||
|
Eocatenipora sp. |
4 |
|||||||||
|
Halysites borealis (Tchernyshev) |
+ |
|||||||||
|
H. pseudoorthopteroides Tchernyshev |
4 |
|||||||||
|
Favositidae |
||||||||||
|
Palaeofavosites borealis Tchem. |
4- |
|||||||||
|
Pf. alveolaris (Goldf.) |
4 |
4 |
||||||||
|
Pf. balticus (Rukh.) |
+ |
|||||||||
|
Pf. borealis Tchernyshev. |
4 |
|||||||||
|
Pf. felix Sokolov |
4 |
|||||||||
|
Pf. cf. nodousus Poul. |
4 |
|||||||||
|
Pf. maximus Tchernyshev. |
4 |
4 |
||||||||
|
Pf. paulus Sokolov |
+ |
|||||||||
|
Pf. schmidti borealis Sokolov |
4 |
|||||||||
|
Pf. schmidti schmidti Sokolov |
4 |
4- |
||||||||
|
Multisolcnia formosa Sokolov |
4- |
|||||||||
|
Ms. aff. misera Sokolov et Tesakov |
4 |
|||||||||
|
Ms. nikiforovae Sokolov et Tesakov |
4 |
|||||||||
|
Ms. tortuosa Fritz |
4 |
4 |
4 |
4 |
4 |
|||||
|
Ms. uralica Chekhovich |
4 |
4 |
||||||||
Окончание таблицы
Table ending
Перерыв в осадконакоплении на границе лландове-ри и венлока (маршрутнинского и войвывского горизонтов), установленный в опорном разрезе силура, позволяет предположить отсутствие ранневенлокских (шейнвуд) отложений на западном склоне Приполярного Урала [3, 7]. Трансгрессия в начале войвывского времени (гомер) способствовала широкому распространению бентосной фауны, в том числе брахиопод рода Spirinella, известного в венлоке и лудлове Алтая [7], о. Готланда, Великобритании, в лландовери и венлоке Сев. Америки и Арктической Канады, венлоке Австралии [18], лудлове Эстонии ( Quadrithyris sinuate ) [14], Богемии, Монголии и на юге Китая [17, 18], что свидетельствует о возможности свободной миграции в пределах обширных акваторий в венлоке [3]. Родовой состав венлокских табулят качественно не изменился, вместе с тем видовое разнообразие пополнили 11 видов (см. таблицу). Присутствие в венлокском разрезе таких космополитных видов, как Favosites gothlandicus Lam., Sapporipora favositoides (Tchern.), Parastriatopora commutabili Klaamаnn [10, 11], характерно для большинства палеобассейнов Урало- Новоземельско-го региона. Еще одной особенностью в развитии североуральских табулят венлока является формирование небольших по мощности слоев, переполненных ветвистыми колониями Parastriatopora commutabilis Klaam., P. admirabile Chekh., P. arctica (Tchern.), P. tebenjkovi (Tchern.). Эти слои приурочены к нижней половине разреза войвывского горизонта венлока и являются маркирующими в разрезах западного склона Урала [5, 11] (рис. 2). Исчезновение ветвистых табулят в верхней части разреза венлока, вероятно, связано с изменением среды обитания, сменой отмельных фаций тиховодных участков в морском бассейне на менее благоприятные условия существования.
На поднятии Чернова в разрезе венлока автором установлены тонковетвистые колонии табулят Aulocystella aseptata (Barskaja), захоронения которых, возможно, представляли собой коралловые луга, заселявшие мелководье ниже базиса волн (рис. 1), [4, 12].
К началу венлокского времени исчезли табуляты широко распространенного в раннем силуре рода Mesofavosites и появились роды Thecia, Laceripora, Riphaeolites, Coenites, которые принимали участие в формировании биогермов в лудловское время.
Выводы и обсуждение результатов
Таким образом, поздний ордовик был временем расцвета мелкоячеистых лихенариид, цепочечных хализитид, примитивных гелиолитид и ознаменован появлением первых фавозитид. В течение всего позднего ордовика происходило постепенное развитие целого ряда кратковременно существовавших стенофациальных видов. Заметное количественное и качественное обновление фауны табулят произошло в кырьинское (катийское) время.

Рис. 2. Ветвистые колонии табулят Parastriatopora commutabilis Klaam. Венлок. Обн. 212, сл. 8Г, р. Кожым, Приполярный Урал
Fig. 2. Bed containing branching colonies of Parastriatopora commutabilis Klaam. The Wenlockian, section 212, bed #8G, Kozhim River, Subpolar Urals
В раннесилурийское время доминирующее положение в сообществах табулят занимали фавозитиды. Крупноячеистые представители родов Palaeofavosites, Mesofavosites и Favosites составляли более половины от общего числа видов. Следующими по численности видов были хализитиды родов Catenipora и Halysites. В конце раннего силура преобладающими наряду с фавозитидами стали представители сирингопорид (Syringopora) и пахипорид (Parastriatopora).
Благоприятное влияние на разнообразие табулят оказывали отмельные зоны и стабилизация условий осадконакопления в лландовери (филиппьельское, маршрутнин-ское время) и в венлоке (войвывское время). На первый план здесь выступали эволюционные механизмы.
Динамика разнообразия табулятовой фауны в Североуральском регионе имела тесную связь с обстановками осадконакопления в бассейне, с региональными и глобальными событиями. Максимум их разнообразия приходился на периоды стабилизации морского палеобассейна. Анализ закономерностей распространения табулят в разрезе позволяет констатировать, что последовательная смена комплексов табулят в позднем ордовике и в раннем силуре на территории региона в большей степени определялась эвстатическими колебаниями уровня моря, миграцией фауны в палеобассейнах и в меньшей степени эволюционными процессами.
Полученные результаты изучения табулят имеют важное значение для биостратиграфических, биофациальных, палеогеографических построений и корреляции отдельных стратиграфических уровней в пределах изученной территории и смежных регионов.
Список литературы Палеоэкологические и палеобиогеографические особенности в эволюции табулят в позднем ордовике и раннем силуре на севере Урала
- Атлас литолого-палеогеографических карт палеозоя и мезозоя Северного Приуралья. Л.: Наука, 1972. 112 с.
- Барская В. Ф. О некоторых силурийских фавозитидах Приполярного Урала//Геология, палеонтология, палеогеография. М., 1973б. С. 44-61.
- Безносова Т. М. Сообщества брахиопод и биостратиграфия верхнего ордовика, силура и нижнего девона северо-восточной окраины палеоконтинента Балтия. Екатеринбург: УрО РАН, 2008, 217 с.
- Безносова Т. М., Лукин В. Ю. Матвеев В. А. Захоронения сообществ донных организмов в венлокских отложениях на поднятии Чернова//Труды Палеонтол. общества. Под ред. С.В. Рожнова. М., 2018. Том I. С. 23-28.
- Безносова Т. М., Лукин В. Ю., Мянник П. О возрасте горизонтов нижнего силура (Тимано-Североуральский регион)//Вестник Института геологии Коми НЦ УрО РАН. 2016. № 11. С. 12-15.
