Палеоэкологические и тафономические особенности строматопороидей

Бесплатный доступ
Короткий адрес: https://sciup.org/149128044
IDR: 149128044
Текст статьи Палеоэкологические и тафономические особенности строматопороидей
В силурийских отложениях Приполярного Урала широко распространены бентосные организмы. Среди них по численности доминируют стромато-пороидеи, которые составляют значительную часть бентосных и рифообразующих организмов. Силурийские строматопороидеи характеризуются большой экологической толерантностью. Различные по форме и размерам, они слагают многочисленные мелкие биогермы и биостромы или встречаются скоплениями или разрозненно в породах карбонатного ряда.
В ископаемом состоянии стромато-пороидеи хорошо сохраняют внешнюю форму и дают возможность судить о палеоэкологических условиях окружающей среды: характеризуют палеобиогеографические условия, гидродинамический режим, характер грунта, климатические факторы условий осадконакопления.
Влияние палеоэкологических обстановок на формирование скелетной постройки строматопороидей.
Верхнеордовикские и силурийские отложения Урала, в особенности Северного, Приполярного и Полярного, характеризуются широким распространением строматопороидей и широким участием их в породообразовании, а также в строении органогенных отложений, биогермов и биостромов [6].
Оптимальными условиями развития строматопороидей были небольшие глубины (до 25—30 м) со слабой волновой активностью, нормальной соленостью, аэрацией, газовым режимом, температурой воды около +20 °С, обилием света [5]. Подобные условия обеспечивались в зоне прибрежного мелководья и отмелей открытого моря, где формировались органогенно-обломочные известняки [6]. При таких условиях наиболее четко проявляются признаки рода и вида, а форма ценос-теума может служить признаком семейства.
Строматопороидеи реагировали на незначительные изменения среды и по- разному к ним приспосабливались. Они были весьма чувствительны к химизму среды, избегали селиться в зоне литорали и в зонах с особым физикохимическим режимом, где, например, отлагались доломиты. Наличие значительной примеси терригенного материала также не было благоприятным. Изучение внешнего вида и внутренней структуры строматопороидей позволяет восстановить влияние на них различных факторов.
Формы роста . Формирование це-ностеума строматопороидей начиналось после прикрепления личинки к субстрату. В ценостеуме любого вида невозможно проследить какие-либо стадии онтогенетического развития, он сразу приобретал признаки взрослой постройки [3]. Форма ценостеу-ма строматопороидей, их ориентировка, места прикрепления служат индикатором характера окружающей среды, в которой формировались их скелетные постройки, а также силы и направления течения.
В оптимальных условиях суще-
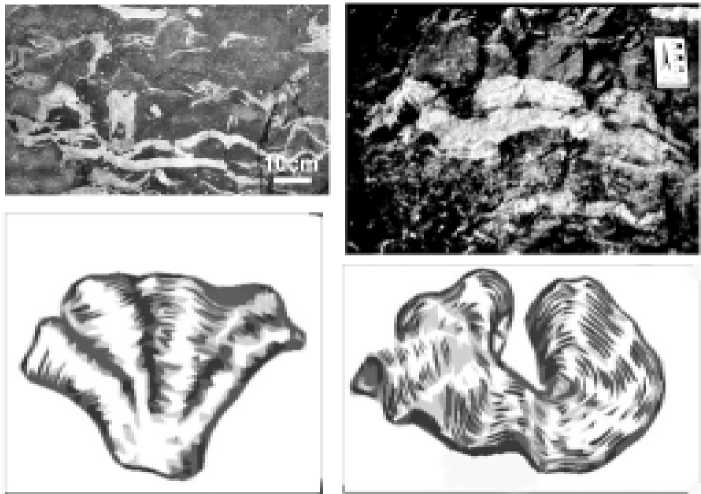
Рис. 1. Корковидная (а) и чашевидная (б) формы роста ценостеумов, развивающиеся при прогрессирующем обмелении. Приполярный Урал, р. Кожим, обн. 229, силур, ллан-довери, лолашорский ярус
ствования популяции ценостеумы приобретали формы, свойственные определенным семействам [3]. При незначительном изменении среды форма ценостеумов строматопорои-дей менялась.
В условиях прогрессирующего обмеления представители семейств, характеризующихся массивными ценос-теумами, не успевали сформировать завершенную постройку — таким образом, форма их близка к пластинчатой, так как у организмов не было возможности роста. Находясь близко к поверхности воды, ценостеумы принимали плоские корковидные или дисковидные формы (рис. 1). Чашевидная форма также свидетельствует об условиях прогрессирующего обмеления. При затрудненном водообмене, повышенной солености и недостатке освещения формируются цилиндрические и ко-люмнарные формы.
При слишком большом объеме поступающего материала скелетные постройки строматопороидей не достигают крупных размеров, ценостеумы ча-

сто имеют желваковидные, лепешковидные формы, нередко обширные по латерали. При высокой скорости осадконакопления строматопороидеи имеют прерывистый боковой край. По нему можно судить о смене обстановок осадконакопления (рис. 2).

Рис. 2. Формирование бокового края ценостеумов строматопороидей, обусловленное изменением количества поступающего осадочного материала
Часто наблюдаемая асимметричная форма строения ценостеумов — утол-щенность с одного конца и, соответственно, смещение центра роста — свидетельствует о поступлении питательного материала с определенной стороны, то есть о наличии течения (рис. 3).
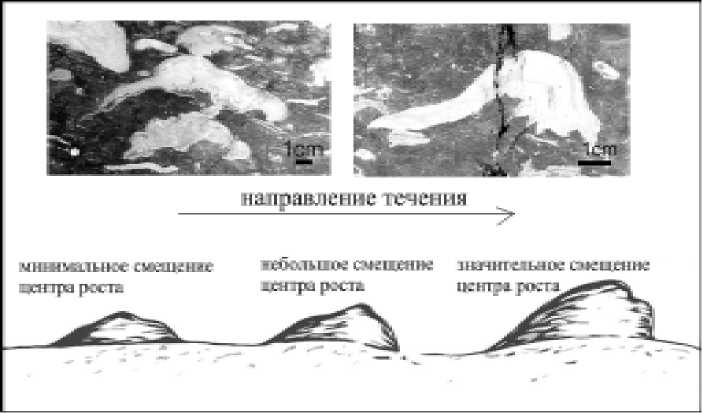
Рис. 3. Асимметричные формы скелетной постройки строматопороидей. Приполярный Урал, р. Кожим, обн. 229, силур, лландовери, маршрутинский горизонт
Формирование внешнего облика зависело не только от условий осадконакопления, но и от грунта. Информацию о грунте, на котором происходило формирование, несет нижняя поверхность скелетной постройки. Она, как правило, вогнута, но может варьироваться в зависимости от характера грунта. Формированию вогнутой нижней поверхности способствовали органогенно-обломочные (криноид-ные, раковинные и др.) грунты. В этом 8
случае часто сохраняется педункула, редко — эпитека.
Менее грубообломочные грунты характеризуются изогнутой, морщинистой нижней поверхностью — эпитека обволакивала неровности грунта. Особенно характерны изогнутость, по- вторение рельефа грунта для пластинчатых форм.
Выпуклая нижняя поверхность свидетельствует о том, что скелет формировался на мягком грунте — края загибались, препятствуя собственному погружению в осадок.
Формы роста строматопороидей, обитающих на биогермах и вне их, а также способы прикрепления и размеры значительно различаются. Для биогермов характерны ценостеумы массивные, полусферические, значительные по площади. В зоне повышенного волнения преобладающей являлась пластинчатая форма колоний, так как она позволяла более надежно закрепиться. Таксономический же состав на биогермах и вне их одинаков. Строма-топороидеи преимущественно сели- лись на колониях табулят. Быстро разрастаясь, они лишали кораллы питания, способствовали их гибели и использовали в качестве субстрата. К такому выводу приводят находки цено-стеумов с заключенными в них колониями табулят, размером до 3 см. У заключенных фрагментов колоний та-булят и гелиолитоидей кораллиты могут быть заполнены породой, что свидетельствует о поселении стромато-пороидей на полипняке после его гибели.
Сходное влияние внешней среды определяет и формирование колоний современных коралловых полипов, развивающихся в мелководных условиях при недостаточной освещенности, нагреве воды, опреснении, затенении колоний и других стрессовых воздействиях. Возможно, строматопороидеи и влияние палеоэкологических обстановок на них можно рассматривать по аналогии с современными кораллами при достаточной изученности этого вопроса.
Необходимо отметить, что под влиянием внешних воздействий ценосте-умы представителей различных видов могли приобретать одинаковую форму, поэтому, несмотря на необходимость принятия во внимание типа ценостеу-ма при определении строматопорои-дей, форма его не несет определяющей информации о таксономической принадлежности.
Внутреннее строение . Внешние условия оказывают влияние и на внутреннее строение строматопороидей. Довольно часто наблюдалось нами явление, когда скелетная постройка определенной особи содержит микропрослойки илистого, а иногда и обломочного материала мощностью до 2 мм, возможно, сносимого с биогермов. Это свидетельствует о перерывах роста и/или периодическом заиливании окружающей среды и связано с временными интервалами, характеризующимися усиленным осадконакоплением [5, 7]. В такие периоды рост ценостеу-ма прекращался и возобновлялся при сокращении поступления материала (рис. 4).
Латиламины также можно рассматривать как показатель периодического замедления роста, когда под влиянием внешних условий нарушается четкость внутренних элементов скелета. Можно сделать вывод, что участки с неправильным скелетом связаны с неблагоприятными для развития ценостеума


Рис. 4. Simplexodictyon kyssuniense c нормальным каркасом (1) и с зональным сгущением элементов (2).
1 — Приполярный Урал, р. Кожим, обн. 229, силур, лландовери, устьдурнаюский горизонт;
2 — Приполярный Урал, р. Кожим, обн. 112, силур, лландовери, устьдурнаюский горизонт
условиями, так как даже в пределах одной колонии могут чередоваться зоны с различным по густоте расположением элементов.
Тафономические особенности строматопороидей.
Степень физического разрушения, наблюдаемого у строматопороидей, характерна и для некоторых других палеозойских организмов. Так как строматопороидеи обладают ломким карбонатным скелетом, то с некоторыми исключениями их разрушение можно рассматривать по методу аналогий с массивными кораллами или хетети-дами [8]. Несмотря на существенные различия в образе жизни и внутреннем строении скелета строматопороидей и других рифостроящих беспозвоночных, можно выделить несколько факторов, влияющих на захоронение стро-матопороидей.
Формы роста. Строматопороидеи представлены богатейшим разнообра-
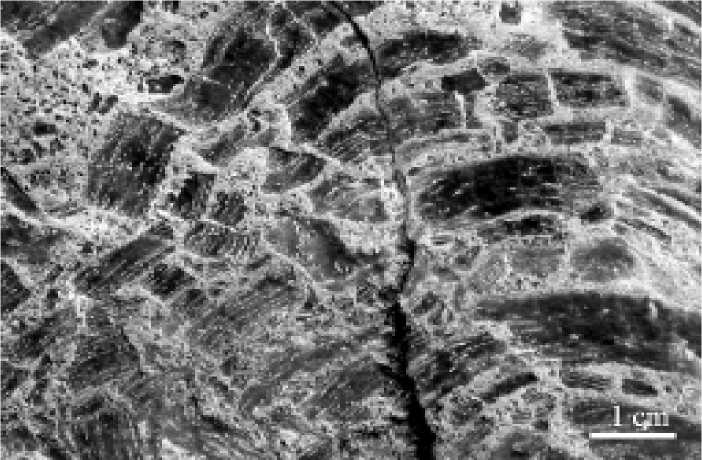
Рис. 5. Диагенетическое разрушение ценостеума: при захоронении латиламины были раздроблены. Приполярный Урал, р. Кожим, обн. 112, силур, венлок, устьдурнаюский горизонт
зием форм ценостеумов: от пластинчатых до куполовидных; от дендроидных (встречающихся, как правило, в виде фрагментов) до сложных, уклоняющихся. Гидродинамически стабильны ценостеумы строматопороидей, обладающие пластинчатой формой или близкой к массивной (но не излишне куполовидной). Стабильность этих форм может быть увеличена наличием соседних особей, которые ограничивают гидродинамическое воздействие — такое явление наиболее актуально в рифах и органогенных постройках [1].
Крупные массивные формы отличаются достаточной сопротивляемостью воздействию среды. Пластинчатые формы расценены наличием меньшей гидродинамической силы, чем массивные. Наиболее слабой устойчивостью подобным воздействиям характеризуются дендроидные формы, встреченные в верхнем силу- ре и, как правило, характерные для амфипорид [2].
Субстрат. Субстрат, на котором развиваются строматопороидеи, представляет собой, как правило, карбонатные мадстоуны и рудстоуны, редко песчаники. Наибольшая приуроченность выявлена к карбонатным мелководным фациям. Строматопороидеи, развивающиеся на грубом карбонатном субстрате, более устойчивы к гидродинамическим потокам [8]. Скелеты строматопо-роидей на карбонатно-терригенном грунте наиболее часто сохраняются в ненарушенном и неперемещенном виде. При большой примеси терригенного, в частности песчаного, материала формируются пластинчатые формы, которые могут транспортироваться по дну моря.
Разрушение строматопороидей диктуется строением ценостеума и местом обитания (формы роста ценостеума, плотность скелета, степень прикрепления ко дну моря, субстрат и обстановки осадконакопления). Физическая поломка и диагенетическое разрушение ценостеума в соответствии с оказанными давлениями очень влияют на степень изменчивости форм роста (рис. 5). Хотя перелом ценостеума может быть случаен, при повреждения при седиментации на ранних этапах роста, однако образование определенных элементов строения (например, ла-тиламин и параламин) создает в цено-стеуме слабые участки, такие, что разрушение происходит согласно этим элементам, образуя изогнутые или плитчатые фрагменты (рис. 6). Образцы из отложений раннего силура часто являются иллюстрацией тенденции строматопороидей раскалываться по латиламинарным поверхностям, либо по слабым участкам скелета (строение с разреженной плотностью).
Изучение палеоэкологических и тафономических особенностей, фациальной приуроченности строматопоро-идей, выявление палеобиоценозов с их участием имеют большое значение для палеогеографических и палеоэкологических исследований. Эврифациаль-ные виды строматопороидей одними из первых осваивали пригодные для жизни территории палеобассейна после глобальных событий. Сообщества строматопороидей играли одну из ведущих ролей в процессе развития трофической структуры позднеордовикских и силурийских биоценозов Приполярного Урала.


ПЛАСТИНЧАТЫЕ ФОРМЫ разрушаются по лЗгиламннам рвзрушаюгся поперек
МАССИВНЫЕ ФОРМЫ
ЦИЛИНДРИЧЕСКИЕ ФОРМЫ колюмнарныс дендроидные разрушаются иес[[прадични разрушаются по лвтипаыннйм разругаются по кем направлениям
Рис. 6. Схема разрушения строматопороидей по дифференцированным формам роста
Список литературы Палеоэкологические и тафономические особенности строматопороидей
- Антропова Е. В., Лукин В. Ю. Бентосные сообщества лландоверийского времени Североуральского палеобассейна // Современная палеонтология: классическая и нетрадиционная: Тезисы докл. LII сессии Палеонтолог. общества. СПб, 2006. С. 8-9.
- Богоявленская О. В. Опыт эколого-стратиграфического изучения органогенных построек (на примере среднего-верхнего девона западного борта магнитогорского синклинория) // Известия Уральской государственной горно-геологической академии. 2000. Вып. 10. С. 12-18.
- Богоявленская О. В. Строматопораты: морфология, систематическое положение, классификация и стратиграфическое значение: Автореф. дис.. доктора геол-минер. наук. М. 1976.
- Богоявленская О. В., Елькин Ю. А. Особенности зарождения и расселения строматопорат в раннепалеозойских бассейнах // Палеонтологическая летопись региональных и глобальных событий: Тезисы докл. LI сессии Палеонтолог. общества. СПб, 2005. С. 20-21.
- Большакова Л. Н. Строматопороидеи силура и нижнего девона Подолии. М.: Наука, 1973. 135 с.
- Опорные разрезы верхнего ордовика и нижнего силура Приполярного Урала. Сыктывкар: Коми фил. АН СССР, 1987. 94 с.
- Kershaw S. Stromatoporoid Grouth form and Taxonomy in a Silurian Biostrome, Gotland // Journal of Palleontology. 1981. V.55, № 6. P. 1284-1295.
- Kershaw.S., Brunton F.R. Palaeozoic stromatoporoid taphonomy: ecologic and environmental significance // Palaeogeography, palaeoklimatology, palaeoecology. 1999. № 149. P. 313-328.
