Палиностратиграфия и условия формирования Майкопской свиты Шамаха-Гобустанской области Азербайджана

Автор: Ализаде аК.А., Тагиева Е.Н., Байрамова Ш.Ш.
Журнал: Вестник Пермского университета. Геология @geology-vestnik-psu
Рубрика: Палеонтология и стратиграфия
Статья в выпуске: 4 (25), 2014 года.
Бесплатный доступ
По разрезам Шамаха-Гобустанской области выделены палинокомплексы для отдельных ярусов майкопской свиты, определяющие условия их формирования. Все палинокомплексы характеризуются сочетанием «полтавской» паратропиче-ской эоценовой флоры с элементами вечнозеленой средиземноморской и умеренной листопадной флоры «тургайского» типа, существовавшими в условиях влажного субтропического типа климата со среднезимними температурами в предгорье не ниже 15 оС. Палинокомплексы среднего слоя хаттского яруса отличаются высоким содержанием пыльцы теплоумеренной и умеренной флоры, по сравнению с остальными олигоценовыми комплексами, что соответствует снижению среднегодовых температур. В начале раннего миоцена (кавказский век) климатические условия стабилизируются с сохранением в составе растительности листопадной умеренной флоры.
Майкопская серия, спорово-пыльцевые спектры, олигоцен, миоцен, кавказ, рюппель, хатт
Короткий адрес: https://sciup.org/147200924
IDR: 147200924 | УДК: 551.8
Palynostratigraphy and environments of formation of Maykop suite of Shemakha-Gobustan region of Azerbaijan
The palynocomplexes of individual Stages of the Maykop Suit, indicating their formation environments, were received from the geological sections of Shamakh-Gobustan region. All the palynocomplexes are characterized by combination of "Poltava" paratropical Eocene flora and elements of evergreen Mediterranean, and temperate deciduous "Turgay" type flora, existed in the humid subtropical climate with the average winter temperatures in the foothills not less than 15° C. Comparing to other Oligocene complexes, palynocomplexes of middle layer of the Hutt Stage are characterized by high content of warm-temperate and temperate flora pollen, which existence indicates the decrease of average annual temperatures. At the beginning of the Early Miocene (Caucasian century), climatic conditions were stabilized preserving a deciduous temperate flora in the vegetation structure.
Текст научной статьи Палиностратиграфия и условия формирования Майкопской свиты Шамаха-Гобустанской области Азербайджана
Предмет исследования. Майкопская серия представляет собой стратиграфическое подразделение на границе палеогена и неогена. Впервые майкопская толща (серия) была установлена И. М. Губкиным [7] и в дальнейшем изучалась целым рядом исследователей.
В Понто-Каспийской шкале майкопская свита соответствует олигоцену и нижнему миоцену, подразделяется на 3 подотдела и различное число свит и горизонтов [16]. Нижний подотдел соответствует нижнему олигоцену (стампийскому или рюппельскому ярусу – 36,5–31,5 млн лет назад), средний – верхнему олигоцену (хаттскому ярусу – 31,5–24,0 млн лет назад), верхний – нижнему миоцену (от 24 до 16,5 млн лет назад).
Раннему миоцену в Понто-Каспии соответствует верхний подотдел майкопской толщи (серии). Он имеет одну датировку (из Предкавказья) – 23,3 млн лет назад [18] и подразделяется на 3 яруса -(снизу-вверх) – кавказский, сакараульский и коцахурский. Они не датированы, но хорошо сопоставляются с эгериевским, эггенбургским и оттнангийским ярусами Западного Паратетиса, датируемыми 24– 22, 22–19 и 19–17,5 млн лет назад [5, 9, 12]. Их возраст нами принят за возраст соответствующих ярусов Понто-Каспия (за исключением верхней границы коца-хурского яруса)1. К раннему миоцену относится и тарханский ярус (17,2–16,5 млн лет назад)
История изучения. Растительные остатки в отложениях майкопской серии были обнаружены благодаря геологическим работам, проводимым по заданию Нобеля в 1885, 1889 гг. У селения Пере-кишкюль, в бассейне р. Сумгаитчай, в верхнемайкопских отложениях шведским геологом Х. Шегреном (H. Sjogren) обнаружена ископаемая древесина, изученная в дальнейшем немецким ботаником И. Феликсом [23]. Вторичные сборы в этом районе были произведены И.В. Палиби-ным [14], а затем изучались А.В. Ярмоленко [22].
Богатый комплекс раннеолигоценовой ископаемой флоры на горе Даррыдаг (На-хчыванская АР) исследовал И.В. Палибин [15], а позже Г.М. Касумова [10]. Отличительной особенностью этой флоры является присутствие пальм, папоротников и тропических видов растений. Позднее списки флоры пополнились исследованиями М.А. Ахметьева и Н.И. Запорожец [1], которые обнаружили в шести флороносных слоях значительное присутствие пальм (Sabal, Trachycarpus) и уточнили возраст этих отложений – не моложе раннего олигоцена. Ими же приводятся и данные спорово-пыльцевого анализа (более 100 зерен пыльцы и спор) из III флороносного слоя. Низкое содержание в пали-нокомплексе спор папоротников (2%) и пыльцы пальм (8%) по сравнению с массовой встречаемостью этих растений в макроостатках (90%) ученые объясняют приуроченностью пальм и папоротников к долинным азональным группировкам.
Олигоценовая флора северовосточных склонов Малого Кавказа и Та-лыша (Лерикский район) была подробно изучена Г.М. Касумовой [11].
Палинологическое исследование майкопских отложений впервые приводится Х.С. Джабаровой [8] по разрезам из Джа-лилабадского района (Талыш), что пополнило списки макрофлоры в этом районе. При изучении стратиграфии майкопских отложений Центрального Гобустана был проведен комплексный анализ, в том числе и палинологический, по нескольким разрезам майкопской серии, однако результаты этих исследований подробно в печати не освещены [2, 21].
Спорово-пыльцевые спектры стратотипических разрезов с учетом комплекса макрофоссилий являются надежной основой для восстановления изменений расти- тельности, климата, природной среды [3, 4, 13, 17] и полноценным критерием достижения стратиграфических целей.
Результаты исследований. На территории Азербайджана майкопские отложе-
ния наиболее полно представлены в юговосточном окончании Большого Кавказа в Шамах-Гобустанской области (рис.1).
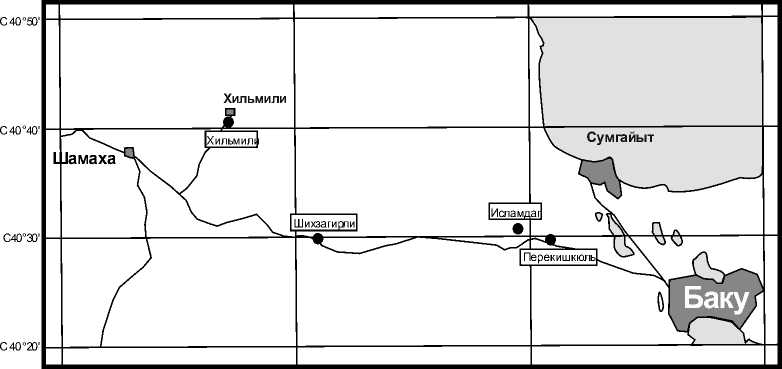
Рис. 1. Схематическая карта расположения разрезов Шамаха-Гобустанского района
Они датированы по фауне и в большом количестве содержат останки рыб, фораминифер, радиолярий, моллюсков, диатомей и остатки растений. Нами были изучены палинологические комплексы нефтеносной майкопской серии в этом районе по разрезам Перекишкюль, Ис-ламдаг, Шихзагирли. Корреляция и сопоставление спорово-пыльцевых спектров этих разрезов позволили выделить палинологические комплексы для каждого хронологического среза и проследить из- менения растительной биоты и условий их обитания на протяжении позднего палеогена.
Разрез Перекишкюль (рис. 2) охватывает стратиграфические единицы хадума, нижнего и верхнего майкопа. Аналогичные отложения представлены и в разрезе Исламдаг (рис. 3), но здесь отсутствует хадумский горизонт. Разрез Шихзагирли (рис. 4) характеризует верхний подотдел майкопской серии (кавказский, сакарауль-ский и коцахурский ярусы).
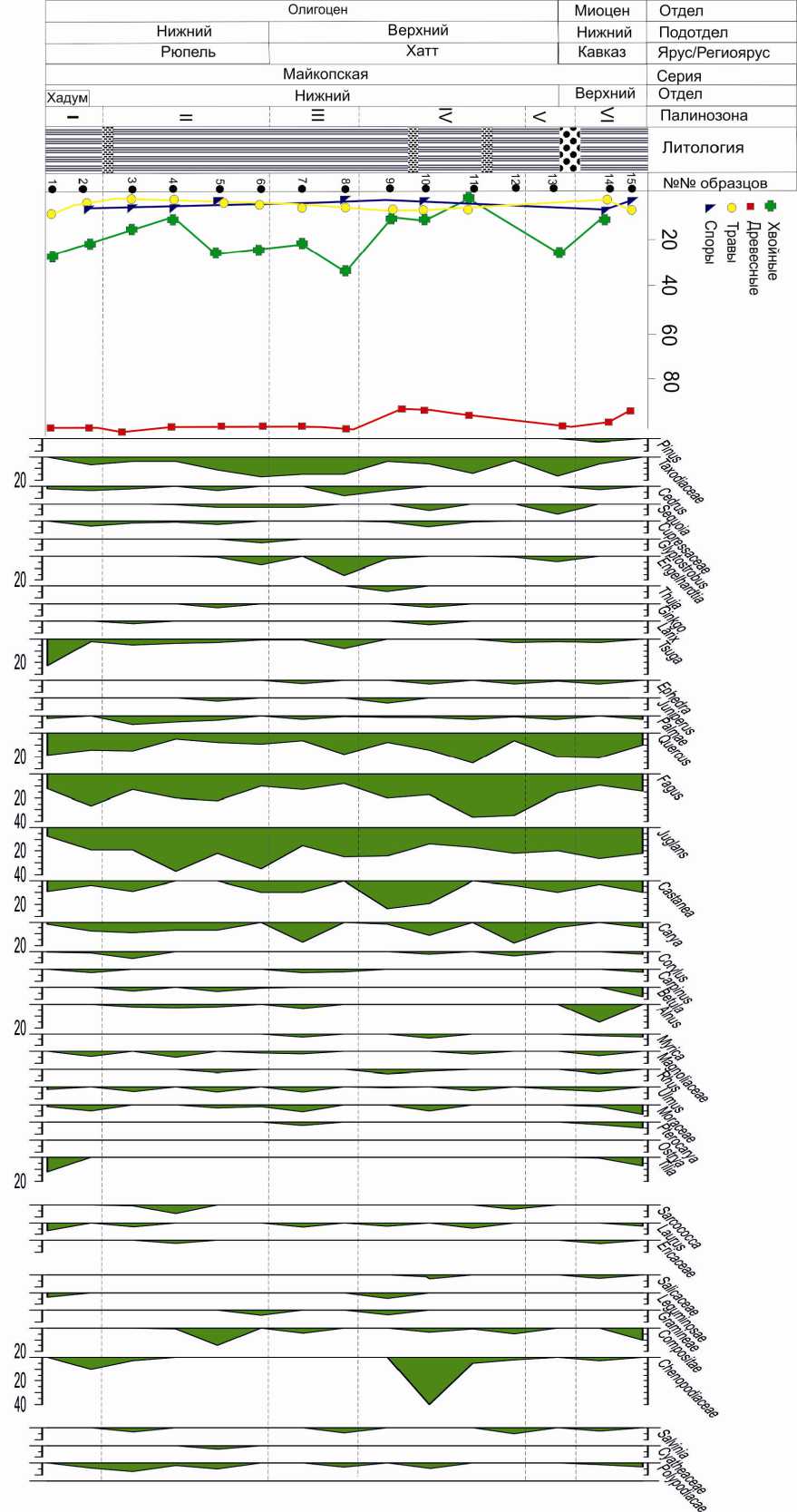
Рис. 2. Спорово-пыльцевая диаграмма разреза Перекишкюль
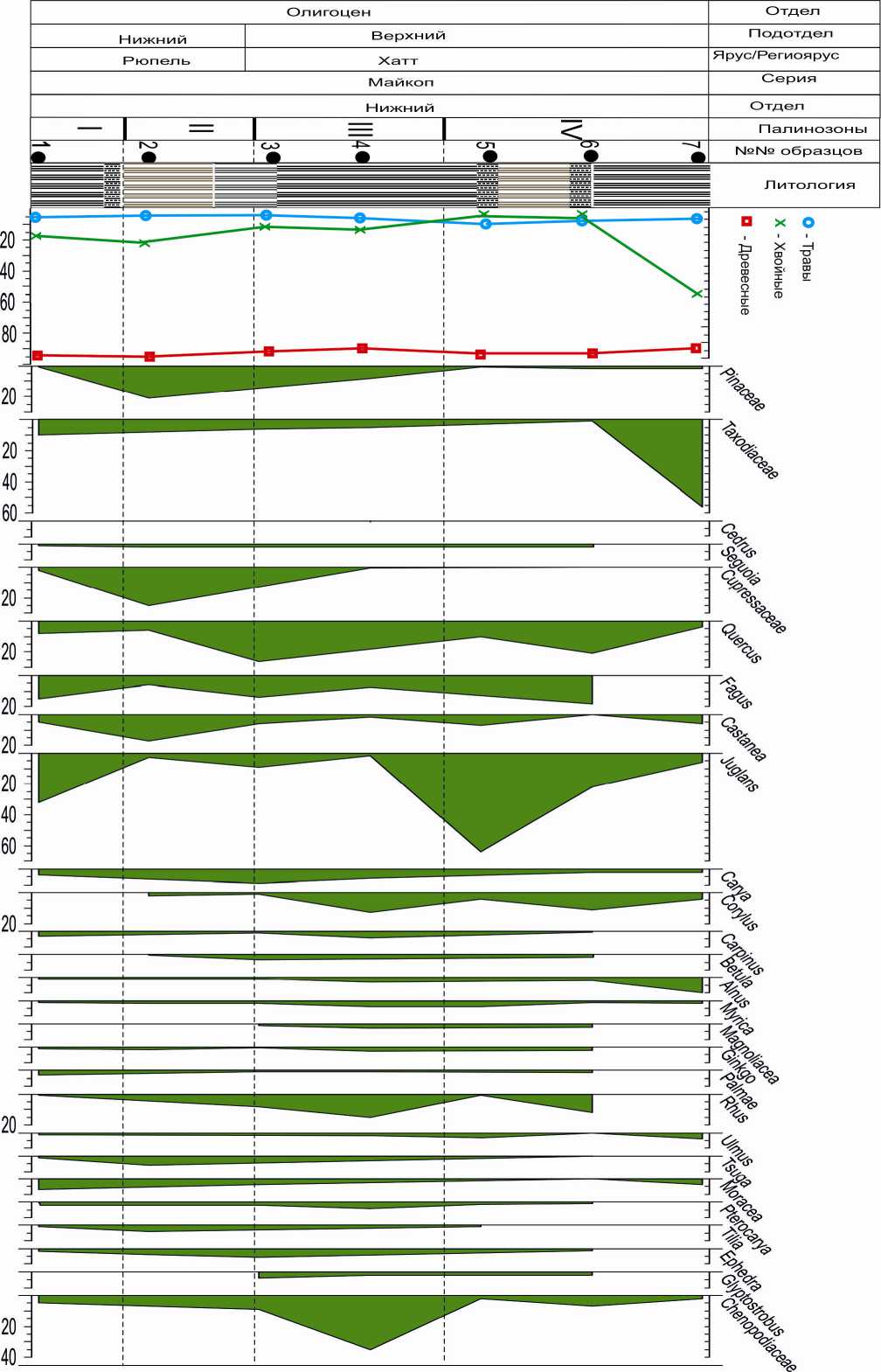
Рис. 3. Спорово-пыльцевая диаграмма разреза Исламдаг
|
Миоцен |
Отдел |
|||||||
|
Нижний |
Подотдел |
|||||||
|
Кавказ |
Коцахур |
Сакараул |
Ярус/Региоярус |
|||||
|
IV |
айкопская |
Серия |
||||||
|
Верхний |
Отдел |
|||||||
|
— |
— — |
< |
< |
< |
< |
Палинозоны |
||
|
• • • |
• • |
№№ образцов |
||||||
|
-•:• ■ ^^—в 1 *• • । 1 ™ - |
Литология |
|||||||
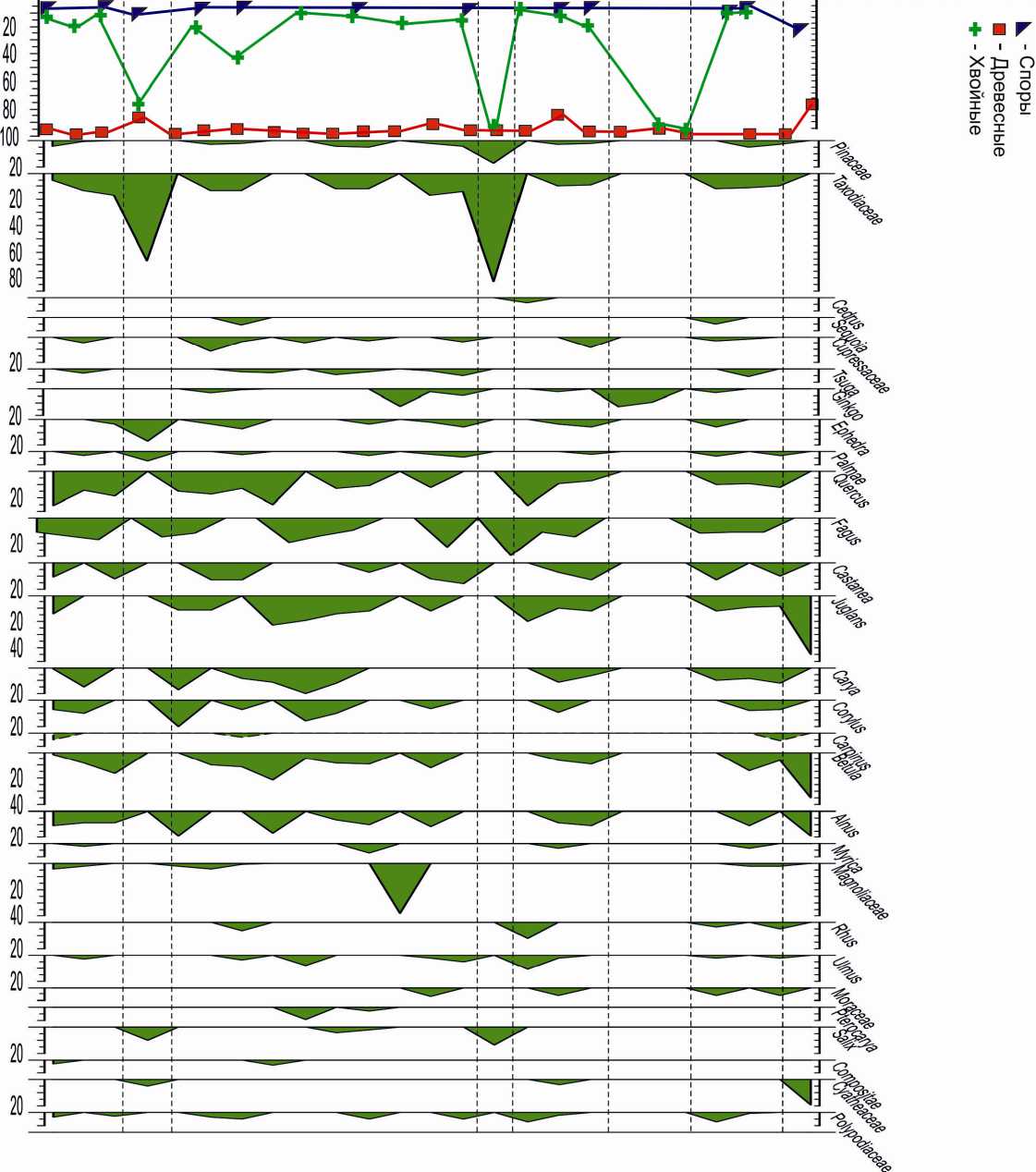
Рис. 4. Спорово-пыльцевая диаграмма разреза Шихзагирли
Во всех спорово-пыльцевых спектрах вие пыльцы травянистых растений и спор в группе пыльцы общего состава домини- отмечается единично.
рует пыльца древесных пород, присутст-
Основание майкопской свиты, хадум-ский горизонт
Общая мощность 36 м. Представлен в разрезе Перекишкюль – палинозона I (обр. 1, 2). В общем составе содержание пыльцы древесных пород равно 91% и трав – 9%. Среди пыльцы покрытосеменных преобладают представители семейства Fagасеае – 35% (Fagus – 12–27%, Quercus – 14–19%, Castanea – 4–9%) и Juglandасеаe – 22% (Juqlans sp. – 11%, Juqlans polyporata – 3%, Carya – 2–8%, Carya spackmania – 2%, Engelhardtia – 2%). Единично отмечена пыльца Alnus, Corylus, Carpinus, а также Tilia, Palmae, Magnoliaceae, Morаceaе, Myrica. В группе пыльцы древесных пород голосеменные растения составляют 26–18%, среди них выделяются Tsuga, Cedrus с участием Taxodium, Abies, Cupressaceae. Пыльца Pinus и Picea отсутствует. Среди пыльцы травянистых единично отмечается Chenopodiaceae.
Нижний олигоцен, нижние слои рюпеля
Представлены в разрезах Перекиш-кюль, палинозона II (обр. 3, 4, 5, 6), и Ис-ламдаг, палинозона I (обр. 1). Среди пыльцы широколиственных пород доминируют Juglans – 17–35%, Quercus – 5– 25%, Fagus – 10–22%, присутствуют также Castanea – 5–9%, Carya, Carpinus и единично Betula (только в разрезе Перекиш-кюль), Alnus, Corylus, Ulmus, Moraceae, Magnoliaceae. Отмечено до 1–2% пыльцы древесных ксерофитов (Rhus, Pistaсiae, Ephedra). Процентное содержание голосеменных в группе древесных сохраняется (14–25%), но разнообразие их увеличивается. Преобладает пыльца Taxodiaceae – 4–17%, присутствует также Tsuga – 4%, Cedrus – 4% (в разрезе Перекишкюль), Ephedra – 4% (в разрезе Исламдаг), содержание пыльцы остальных голосеменных (Sequoia, Cupressaceae, Ginkgo, Glyptostrobus, Pinus (только в Исламдаге), Palmae) составляет 1–2%.
Верхние слои рюппеля палинологиче-ски не охарактеризованы, данные отсутствуют.
Верхний олигоцен, хаттский ярус
Нижние слои хатта представлены па-линозоной III (обр. 7, 8) в разрезе Пере-кишкюль и палинозоной II (обр. 2) в разрезе Исламдаг. В спорово-пыльцевых спектрах пыльца древесных пород составляет 98%. Содержание голосеменных в группе древесных возрастает в разрезе Перекишкюль до 31%, в Исламдаге до 22% за счет пыльцы Taxodiaceae (6–15%) а также Cedrus, Sequoia, Tsuga (в разрезе Перекишкюль) и Gliptostrobus, Ephedra (в разрезе Исламдаг). Среди покрытосеменных доминирует пыльца Juglans – 15– 25%, Quercus – 18%, (в разрезе Исламдаг -25%), Fagus – 6–13%, Castanea – 6–10%, Carya – 9–17%, Engelhardtia – 2–16% (в разрезе Перекишкюль). Единично (1–2%) встречена пыльца Carpinus, Betula (в разрезе Перекишкюль отсутствует), Alnus, Corylus, Ulmus, Moraceae, Magnoliaceae, Laurus, Nissa (в Исламдаг). Cодержание пыльцы Palmae составляет 2–3%. Пыльца травянистых растений представлена водно-болотным родом Salvinia в разрезе Пе-рекишкюль.
Надо отметить, что палинозона II (обр. 2) в разрезе Исламдаг стратиграфически соответствует верхам рюппельского яруса (конец раннего олигоцена). Однако по соотношению и составу пыльцы и спор она коррелируется с палинозоной III (низы хатта) разреза Перекишкюль. Соответственно граница между рюппелем и хаттом в разрезе Исламдаг, возможно, несколько завышена и ее надо проводить ниже.
Средние слои хатта представлены па-линозоной IV в разрезе Перекишкюль (обр. 9, 10, 11, 12) и палинозоной III (обр. 3, 4, 5) в разрезе Исламдаг. Содержание хвойных в группе древесных пород сокращается (4–16%) в основном за счет пыльцы Taxodiaceae (3–6%), но появляется пыльца Cedrus, Pinus (9%), Larix, Thuja, Juniperus. Спорадически присутствует пыльца, Sequoia, Gliptostrobus, Cupressus, Ginkgo, Tsuga, Ephedra. В группе пыльцы широколиственных пород доминируют Fagus (17–36%), Juglans (15–24%), Quercus (8-25%), Castanea (4–24%). Увеличивается присутствие пыльцы Carya (6–18%), Myrica (до 4%), Rhus (4–15%), Corylus (до 13 %). Отмечается пыльца Magnoliaceae, Ulmus, Moraceae, Pterocarya, Tilia, Betula, Alnus, Ephedra, а также Palmae. Присутствие Engelhardtia, Laurus отмечено только в разрезе Перекишкюль. В группе пыльцы травянистых содержание пыльцы Chenopodiaceae несколько увеличивается.
Верхние слои хаттского яруса представлены палинозоной V в разрезе Пере-кишкюль (обр. 13) и палинозоной IV в Исламдаг (обр. 6, 7). Они характеризуются увеличением пыльцы хвойных пород в группе древесных за счет пыльцы Taxodiaceae (до 28%), Sequoia (9%) с участием пыльцы Pinus, Glyptostrobus, Cupressaceae, Ginkgo (Исламдаг), Tsuga, Ephedra. Среди пыльцы широколиственных пород преобладают Juglans (20–22%), Quercus (21%), Fagus (16–18%), Corylus (11%), Castanea (10%). Единично встречена пыльца Engelhardtia, Carya, Carpinus, Alnus, Ulmus, Pterocarya, Palmae, Rhus, Betula.
Нижний миоцен (верхний майкоп), кавказский ярус
Нижние слои кавказского яруса характеризуются палинозоной VI (обр. 14, 15) разреза Перекишкюль и палинозоной I (обр. 1, 2, 3) разреза Шихзагирли. В группе древесных пород преобладает пыльца Quercus (14–19%), Fagus (11–17%), Juglans (14%), Betula (2–16%), Carya (5–15%), Castanea (12%). Среди голосеменных (9– 22%) встречена пыльца Taxodiaceae (5– 17%), Cupressaceae (5%), Pinaceae (1–4%).
Средние слои кавказского яруса представлены палинозоной II (обр. 4) разреза Шихзагирли. Здесь встречена пыльца исключительно голосеменных растений: Taxodiaceae (67%), Palmae (7%), Ephedraceae (17%), пыльца покрытосе- менных отсутствует. Низкое содержание и аналогичное соотношение пыльцы наблюдается в образце 27 (30–35 зерен на весь объем осадка)2, а в образцах 35, 37 подсчитано 10 зерен, принадлежащих Ginkgo.
Зоны с низким содержанием пыльцы, в основном голосеменных растений, отражают растительность исключительно прибрежной зоны, как влажной, так и сухопесчаной. Учитывая летучесть пыльцы хвойных и перенос ее на дальние расстояния, а также отсутствие малолетучей пыльцы покрытосемянных в этих образцах, можно говорить о расширении прибрежной зоны и, возможно, о регрессивной фазе морского бассейна.
Нижний миоцен, сакараульский ярус
Верхние слои кавказского и нижние слои сакараульского ярусов характеризуются палинозоной III (обр.5-14) разреза Шихзагирли. Среди пыльцы покрытосемянных доминируют Quercus (11-25%), Fagus (10-23%), Betula (10-21%), Juglans (11-23%), Castanea (7-16%), Carya (1120%), единично Palmae, Magnolia. Среди голосеменных преобладает пыльца Taxodiaceae (12-17%) с участием Cupressus (1-11%), Sequoia (6%), Tsuga, Pinus (по 2-5%), Ginkgo (1-5%), Ephedraceae (2-7%).
Средние и верхние слои сакараульско-го яруса характеризуются палинозонами IV (обр. 15), V (обр. 16, 17, 18), VI (обр. 19, 20, 21) разреза Шихзагирли2. Палино-зоны IV и VI представлены пыльцой исключительно голосеменных. Соответственно в палинозоне IV отмечена пыльца Taxodiaceae (83%), Pinaceae (17%), в па-линозоне VI – только пыльца Ginkgo (10 зерен).
Палинозона V (обр. 16, 17, 18) характеризуется преобладанием пыльцы Fagus
(11-29%), Quercus (7-26%), Juglans (1020%), Castanea (7-13%), Betula (6-9%), Carya (6-11%) и единично Palmae. Среди голосеменных присутствует пыльца Taxodiaceae (10%), Cupressaceae (8%), Pinaceae (3%), Ephedra (4-6%) и единично Cedrus, Ginkgo.
Нижний миоцен, коцахурский ярус
Нижние и средние слои коцахурского яруса характеризуются палинозоной VII (обр. 22,23) разреза Шихзагирли. Среди покрытосеменных доминирует пыльца Quercus (9-12%), Fagus (11-12%), Juqlans, Carya (по 8-12%), Castanea (10-13%), Betula (6-14%), а также Magnolia, Palmae и другие. Хвойные представлены пыльцой Taxodiaceae (10-12%), Pinaceae (3-5%), Sequoia (5%), Tsuga (6%), Ephedra (6%) и единично Ginkgo, Cupressaceae.
Верхние слои коцахурского яруса характеризуются палинозоной VIII (обр. 24, 25) разреза Шихзагирли. Пыльца хвойных в образце не встречена, среди покрытосеменных присутствуют только Juglans (45%), Betula (5%), Alnus (20%). Среди спор отмечено максимальное на протяжении спектра присутствие папоротников семейства Cyatheaceae.
Обсуждение результатов
В позднем палеогене Кавказ был представлен архипелагом крупных и мелких островов в океане Тетис. В раннем олигоцене большие островные суши Большого, Малого Кавказа и восточная часть Курин-ской впадины в результате тектонических движений значительно расширились и на отдельных участках приобрели горный рельеф, высоты которого не превышали 500 м [18, 19].
Хадумский горизонт (основание майкопской свиты). На островных сушах юго-восточной оконечности Большого Кавказа господствовала лесная растительность, представленная тропическими вечнозелеными влаголюбивыми формами наряду с мезофильными теплоумеренными и умеренными. Лесные формации без ярко выраженного доминанта были представлены смешанными хвойно-широколиственными орехово-буково-дубовыми лесами с участием хвойных пород Cedrus, Abies, Tsuga, Larix; субтропическими лесами с участием вечнозеленых элементов (Laurus, Magnoliaceae, Cupressaceae) и влажными низинными и приречными лесами из Taxodium, Carya, Ulmus, Morus с участием пальм по морскому побережью. Небольшие абсолютные высоты, низкое процентное соотношение пыльцы голосеменных и отсутствие в их составе основных лесообразующих элементов свидетельствуют о том, что чистые хвойные леса в хадуме не формировались. Присутствие в спектрах элементов субтропической и тропической флоры говорит о высоком режиме температур и увлажнении в это время.
Ранний олигоцен (рюпельский ярус). Полидоминантные влажные хвойношироколиственные леса продолжают доминировать. В широколиственных формациях, по сравнению с хадумским веком, некоторое преимущество получает орех, а не бук. Увеличивается разнообразие представителей вечнозеленых восточноазиатских и тропических флор (Sarococca, Engelhardtia, Cyatheaceae). А среди голосеменных растений отмечаются роды, принадлежащие американо-средиземноморско-азиатской, восточноазиатской, североамериканской (пихта, кедр, секвойя, глиптостробус, кипарис, гинкго) географическим группам. Одновременно единично появляются панголарктические представители листопадной умеренной флоры – береза и ольха, отмечается сосна, отсутствующие в хадумском горизонте. Субтропические, средиземноморского типа леса были образованы как вечнозелеными, так и листопадными породами -магнолией, лавром, кипарисом, карией, мирикой. Заболоченные речные и морские террасы занимали формации из болотного кипариса, ольхи и речного кедра. Ксерофиты – сумах, фисташка, эфедра – осваивали сухие открытые пространства. По песчаным побережьям произрастали пальмы.
Наличие в составе флоры видов и родов, произрастающих в условиях с высокой и избыточной влажностью воздуха и почвы (секвойя, гинкго, таксодиум, глип-тостробус), свидетельствует о высоком режиме увлажнения. Присутствие панголарктических родов Betula, Pinus и ксерофитов американо-средиземноморскоазиатской флоры Pistacia, Rhus говорит о меняющихся в сторону похолодания и иссушения условиях климата и начале дифференциации растительных сообществ. Одной из причин таких изменений явилось увеличение общей площади суши и её абсолютной высоты.
Начало позднего олигоцена (хаттский ярус). Основные типы лесных формаций и их распределение сохраняются. На этом фоне отмечается расширение заболоченных таксодиевых ассоциаций с участием Glyptostrobus. В составе полидоминант-ных хвойно-широколиственных лесов уменьшается доля представителей панголарктической группы (Betula, Corylus) и возрастает роль восточно-азиатского рода Engelhardtia – обитателя вечнозеленых влажных лесов Индии, Китая, Малайского архипелага. Этот период характеризуется более благоприятными условиями для обитания гигро- и термофильных пород относительно предыдущего этапа олигоцена.
Середина позднего олигоцена (хатт-ский ярус) характеризуется перераспределением доминантов в хвойношироколиственных лесах с участием восточноазиатских, североамериканских и средиземноморских элементов. На смену мезофильному ореху приходят умеренные и более холодоустойчивые горные породы (бук, дуб, каштан), увеличивается роль сосны. Появляются мелколиственные представители панголарктической географической группы – лещина, береза, восковник. Среди хвойных пород присутствуют роды, устойчивые к низким зимним температурам, – сосна, кедр, лиственница. Это свидетельствует о пониже- нии среднегодовых температур. Сокращение заболоченных таксодиевых формаций и расширение ксерофитных редколесий (можжевельник, эфедра, сумах) с участием трав (маревые) говорит об иссушении. Эти изменения, происходившие в составе субтропической вечнозеленой гигрофильной и мезофильной с элементами тропической растительности флоры, явились следствием проявления сезонности в распределении как температур, так и осадков. Родовой и видовой состав флоры отличается присутствием наибольшего количества умеренных представителей за весь олигоцен.
Конец позднего олигоцена (хаттский ярус). Во влажных полидоминантных лесах вновь увеличивается доля ореха на фоне дуба, бука и хвойных пород – гинкго, секвойи, тсуги. Лещина продолжает оставаться основным элементом подлеска. Роль панголарктических элементов – березы и сосны, появившихся в период похолодания, заметно уменьшается. Состав влажных приречных и низинных лесов обогащается средиземноморскими и восточноазиатскими элементами (кария, пте-рокария, мирика, ольха, ильм). В прибрежно-морских формациях участвовали пальмы. Расширяются заболоченные формации с участием болотного кипариса, в составе которых также участвует восточно-азиатский род – глиптостробус – обитатель заболоченных лесов на залитых водой трясинах. Это говорит о высоком уровне как почвенного, так воздушного увлажнения. Общий состав и распределение родов в группе покрытосеменных и голосеменных растений сходны с ранне-олигоценовым этапом (низы рюппеля) и свидетельствуют о повышении среднегодовых температур и увлажнения относительно середины позднего олигоцена (средний хатт). Определенные нами методом ареалограмм для этого периода количественные показатели климата по разрезу Исламдаг составляли для января 9–130, июля 26–290 оС, среднегодовое количество осадков 1000–1500 мм [6].
Палинокомплексы позднего майкопа (ранний миоцен) свидетельствуют, что преобладающим типом растительности и в раннем и в среднем майкопе были леса, среди которых выделялись прибрежные морские на заболоченных почвах из болотного кипариса, мирики с участием пальм по песчаным побережьям; полидо-минантные многоярусные хвойношироколиственные из бука, дуба, ореха, каштана, березы с участием гинкго, кедра, тсуги, секвойи; средиземноморского типа из карии, магнолии; пойменные леса с различными видами ольхи, ивы, граба, мирики и сухие редколесья с участием сумаха, эфедры. Единичное присутствие пыльцы трав говорит о незначительной роли открытых пространств.
Эти леса существовали в достаточно влажных условиях с практически равномерным распределением осадков в течение года, о чем говорит, как наличие буковых лесов, требовательных к летней влаге и не переносящих длительных периодов засухи, так и присутствие гигрофитов (таксодиевые, мирика, гинкго, циа-тейные папоротники). Большая часть представителей семейства циатейных – это древовидные папоротники влажных горных лесов в тропических и субтропических (юг Японии и Новой Зеландии) областях.
Наличие индикаторных видов, имеющих узкий ареал распространения (гинкго, пальмы, циатейные), свидетельствует о довольно теплых условиях со среднезимними температурами в предгорье не ниже 15оС.
Выводы
Палинокомплексы майкопской серии по разрезам Перекишкюль, Исламдаг и Шихзагирли, расположенных в юговосточном окончании Большого Кавказа, характеризуются сочетанием «полтавской» паратропической эоценовой флоры (пальма, циатейные) с элементами вечнозеленой средиземноморской (магнолия, кипарис, каштан, орех) и умеренной лис- топадной флоры «тургайского» типа (бук, граб, вяз, липа, орешник). Эта растительность существовала в условиях влажного субтропического типа климата.
Во всех разрезах майкопской серии преобладает пыльца древесных пород. В группе голосеменных растений доминирует пыльца Taxodiaceae. Содержание пыльцы сосны в спектрах незначительное, ее единственный максимум (9% пыльцы древесных) приходится на средние слои хаттского яруса. В палинокомплексах раннего рюппеля впервые появляется пыльца березы, что может свидетельствовать как об активизации орогенных процессов, так и о начальном процессе похолодания и снижении среднегодовых температур.
Наибольшие изменения в составе па-линокомплексов фиксируются в середине хаттского яруса, когда возрастает количество умеренной панголарктической и ксе-рофильной средиземноморской флоры.
Палинокомплексы позднего майкопа (кавказ, сакараул, коцахур) не имеют больших различий в своем составе. От нижне- и среднемайкопских (олигоцено-вых) комплексов их отличает отсутствие пыльцы лавра, однако ископаемая древесина Laurinium maicopiae Jarm. (Lauraceae) встречена в верхнемайкопских отложениях у селения Перекишкюль [22] и в более ранних отложениях встречается лавр. От нижнего олигоцена к нижнему миоцену в спектре возрастает содержание пыльцы березы и кипарисовых.
Содержание пыльцы березы до 20%, ольхи до 9% и пыльцы кипарисовых до 11% наряду с участием пыльцы представителей полтавской (Palmae, Ginkgo, Cyatheaceae) и средиземноморской флоры может служить палиностратиграфическим критерием верхнемайкопских (нижнемиоценовых) отложений и признаком их отличия от олигоценовых.
Список литературы Палиностратиграфия и условия формирования Майкопской свиты Шамаха-Гобустанской области Азербайджана
- Ахметьев М.А., Запорожец Н.И. Новые находки пальм из олигоценовых отложений Даррыдага (Нахичеванская АССР)//Бюллетень Московского общества испытателей природы. Отдел геология. 1989. Т.64, вып. 6. С. 57-67.
- Ахметьев М.А., Запорожец Н.И., Головина Л.А., Попов С.В., Сычевская Е.К., Эфендиева М.А., Фейзуллаев А.А., Алиев Ч.С. Новые данные по стратиграфии майкопских отложений Центрального Гобу-стана//Стратиграфия и седиментология нефтегазоносных бассейнов. 2007. № 1, С. 32-53.
- Байрамова Тагиева Е.Н. Палиност-ратиграфия и палеогеография майкопа Абшеронского района//Стратиграфия и седиментология нефтегазоносных бассейнов. 2009. №1. С. 24-32.
- Байрамова Ш.Ш., Тагиева Е.Н. Новые данные о климате и растительности юго-восточной части Большого Кавказа (Гобу-стан) в олигоцене//Известия НАН Азербайджана. Сер. наук о Земле. 2013. №3, С. 43-49.
- Ганзей С.С. Хронология палеогеографических событий позднего кайнозоя Понто-Каспия (по данным метода треков): авто-реф. дис. канд. геогр. наук. М.: Изд-во. МГУ, 1984. 24 с.
- Гричук В.П. Реконструкция скалярных климатических показателей по флористическим материалам и оценка ее точности/Методы реконструкции палеоклиматов. М.: Наука, 1985. С. 20-29.
- Губкин И.М. Майкопский нефтеносный район. Нефтяно-Ширванская нефтеносная площадь//Тр. Геол. ком. Новая серия. СПб., 1912. Вып. 78. 169 с.
- Джабарова Х.С., Касумова Г.М. Дополнительные материалы об ископаемых растениях из майкопских отложений Талыша (на основании отпечатков растений и данных спорово-пыльцевого анализа)//Доклады АН Аз.ССР. 1961. T.XVIII, № 11. С. 1081-1083.
- Зубаков В.А. Глобальные климатические события неогена. Л.: Гидрометеоиздат, 1990. 223 с.
- Касумова Г.М. О флоре нижнего олигоцена в районе Даррыдаг (Нахичевань)//Доклады АН Аз.ССР. 1961. Т. 27, № 1. С. 161-162.
- Касумова Г.М. Флора олигоценовых отложений северо-восточных предгорий Малого Кавказа. Баку. Изд-во АН Аз.ССР, 1966. 85 с.
- Невесская Л.А.,Гончарова И.А., Ильина Л.Б. и др. Региональная стратиграфическая шкала неогена Восточного Паратети-са//Советская геология. 1984. № 9. С. 3749.
- Мамедов А.В., Работина Е.Н. Ландшафтно-климатические условия эоцена и олигоцена на территории Азербайджана//Известия АН СССР. Сер. географическая. 1990. №5. С. 79-87.
- Палибин И.В. Этапы развития флоры прикаспийских стран со времени мелового периода. М.: Изд-во АН СССР, 1936. 213 с.
- Палибин И.В. Нижнеолигоценовая флора горы Даррыдаг в бассейне Аракса (Закавказье)//Тр. Ботанического института АН СССР, 1947. Сер. 1. Вып. 6. С. 7-27.
- Стратиграфический словарь СССР. Палеоген. Неоген. Четвертичная система. Л.: Недра, 1982. 616 с.
- Тагиева Е.Н. Эволюция природных условий Азербайджана в кайнозое//Известия НАН Азербайджана. Серия наук о Земле. 2008. № 2. С. 106-116.
- Хаин В.Е., Шарданов А.Н. Геологическая история и строение Куринской впадины. Баку: Изд-во АН Аз.ССР, 1952. 237 с.
- Халилов Г.А. Палеогеоморфология Азербайджанской части Большого Кавказа в олигоцен-раннемиоценовой эпохе неотектонического этапа//Вестник Бакинского университета. Сер. естественных наук. 2000. №1. С. 204-211.
- Чельцов Г. Абсолютная геохронология. Стратиграфия СССР. Неогеновая система. Полутом Ю. 2. М.: Недра, 1986. С. 387390.
- Шахбазова В.Г. Растительность и климат майкопского времени Восточного Азербайджана по данным спорово-пыльцевого анализа//Azarbaycan Paleontologiya Camiyyatinin 4 sayli toplusu (prof. C.M.Xalilov 100 illiyina hasr olunur). Baki: "Nafta-Press"na§riyyati, 2001. С.119-123.
- Ярмоленко А.В. Ископаемые древесины майкопской свиты юго-восточного Закавказья//Тр. Ботан. ин-та АН СССР. Сер.1. 1941. Вып. 5. С. 13-35.
- Felix J. Untersuchenuberfossilen Holzeraus-dem Caucasus//Ztsch. Deutsch. Geol. Ge-sellsch. Berlin, 1894. Bd. 46. Abt. 1. S. 88-129.
