Параметр Q вместо процента возврата

Автор: Бардин Александр Васильевич
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 1 т.5, 1996 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140148608
IDR: 140148608
Q parameter instead of a percentage of recovery
Текст статьи Параметр Q вместо процента возврата
К настоящему времени предложен целый ряд методов оценки численности и сохраняемости птиц по данным кольцевания и повторного отлова (см. обзоры: Cormack 1979; Nichols et al. 1981; Pollock 1981; Seber 1982; Pollock et al. 1990 и др.). Однако для их применения нужны материалы многолетних исследований, выполненных по заранее продуманной и адаптированной к изучаемому виду программе. Это, в свою очередь, требует больших затрат средств и времени, что не всегда возможно на практике.
Например, несмотря на почти 40-летнее массовое кольцевание воробьиных птиц на Куршской косе, только для одного вида — Fringilla coelebs — удалось собрать материал, более или менее удовлетворяющий условиям применения стохастических моделей мечения и повторного отлова (Бардин 1990). Для остальных же видов отлов большими рыбачинскими ловушками оказался неэффективным способом контроля местных популяций. Вероятность отлова особей в течение сезона сильно варьировала между годами и видами и для большинства воробьиных была значительно ниже 10%. Так, для Sylvia curruca она изменялась от 1.1 до 6.2% и в среднем составила 2.6% (Паевский 1992). Столь низкая эффективность контроля населения птиц с помощью отлова рыбачинскими ловушками оказалась весьма неожиданной. До сих пор на Биостанции ЗИН РАН было принято считать, что с помощью больших ловушек удается поймать 25-30% вернувшихся особей, а "уловистость" ловушек примерно одинакова для разных видов птиц (Соколов 1991).
Отсюда понятно скептическое отношение В.А.Паевского (1985, 1992) к современным методам оценки популяционных параметров: эти методы хорошо "работают" при вероятностях отлова более 50% и становятся практически неприменимыми, если вероятность отлова меньше 10%.
В то же время, если демографические исследования проводить специально, осуществлять направленный поиск окольцованных птиц и применять адекватные экологии и поведению конкретных видов способы контроля, можно значительно повысить эффективность работы. Хороший пример - исследования Ficedula hypoleuca на Куршской косе (Высоцкий и др. 1988), Parus cristatus и Р. montanus в Псковской обл. (Бардин 1988). При вероятностях отлова более 50% качественный материал может быть получен на сотнях и даже десятках окольцованных особей. Если же у исследователя мало шансов повторно встретить меченую особь, недостаточно пометить и десятки тысяч птиц.
Во многих случаях, однако, оценка демографических параметров не является основной целью исследования, и материал собирается попутно при проведении других работ. Собранные таким образом сведения об окольцованных птицах, особенно по малоизученным видам и в неисследованных регионах, также представляют известный интерес, и их часто включают в публикации.
К сожалению, в нашей стране до сих пор сохраняется давно изжитая традиция представлять количественные данные о сохраняемости (возвращаемости) птиц в виде т.н. "процента возврата". Этот показатель отражает главным образом усердие исследователя и поэтому малоинформативен. Попытки улучшить этот показатель путем введения разного рода поправок (например: Соколов 1991) нельзя признать удачными. Этот вопрос уже обсуждался (Бардин 1990, 1993). Следует упомянуть также и об одной очень распространенной ошибке при вычислении доли возврата. Эта величина, по определению, есть отношение количества особей, повторно пойманных на следующий после кольцевания год, к количеству окольцованных. Однако очень часто при ее вычислении в числителе ставят количество окольцованных особей, которые были повторно обнаружены на территории исследования в течение всех последующих после кольцевания лет, пока велись наблюдения. Это ведет к смещению оценки, особенно заметному, когда вероятность отлова мала, а сохраняемость особей велика (Бардин 1993).
Не вызывает сомнений, что использование "процента возврата" в качестве оценки сохраняемости (возвращаемости) абсолютно некорректно. На это уже давно указывали многие авторы (Там же). Данный параметр желательно оценивать с помощью современных стохастических моделей. Однако далеко не всегда и везде возможно проведение детальных популяционных исследований. Поэтому полезно иметь метод, позволяющий сделать хотя бы грубую, прикидочную оценку сохраняемости особей в популяциях по данным мечения и повторного контроля, собранным в привычных для отечественных зоологов условиях работы.
Цель данной статьи - привлечь внимание орнитологов к другому простому, но более корректному способу оценки сохраняемости с помощью метода Q (Бардин 1990).
Перед тем, как приступить к его изложению, надо специально подчеркнуть, что при работе на ограниченных территориях в принципе невозможно разделить смертность и эмиграцию. Нам остается только гадать относительно судеб без вести пропавших особей. Поэтому мы можем оценить лишь количество тех птиц, которые не погибли в течение года и при этом остались жить на изучаемой территории. Эту величину мы называем сохраняемостью (Бардин 1988, 1990, 1993; Высоцкий и др. 1988), а В.К.Рябицев (1993) - показателем возврата. Кстати заметить, что В.А.Паевский (1992) весьма решительно выступает против введения специального понятия сохраняемости. Он по-прежнему считает, что в рамках работы по мечению и повторному отлову птиц на ограниченных территориях все же возможна непосредственная оценка выживаемости. Правда, с недавнего времени ВАПаевский признает, что для этого сначала нужно занять четкую позицию по отношению к возвращаемости выживших взрослых птиц на участок исследования - необходимо допустить, что эта величина всегда составляет 100% (Паевский 1992, с. 82). При этом автор цитируемой статьи полностью игнорирует результаты исследований своего коллеги Л.В.Соколова, чьи взгляды он на словах разделяет и защищает. Согласно этим результатам, полученным на том же материале, на место гнездования возвращается не 100, а лишь 80.0% выживших взрослых S. сиггиса (Соколов 1991, с. 19).
Сохраняемость измеряют различными методами, основанными на разных допущениях. Естественно, что эти оценки могут различаться. Поэтому полезно применять к ним разные обозначения. В частности, показатель, получаемый рассматриваемым ниже методом, бу- дем обозначать через Q. По определению, Q = SF, где S - выживаемость, F- верность особей обследуемой нами территории. Кроме того, обозначим через Р вероятность отлова (встречи) меченой особи, N- количество особей, окольцованных в год z, пк - ожидаемое количество особей из числа N, повторно встреченных в год (z + к).
Как и в случае использования "процента возврата", составных демографических таблиц и их видоизменений, рассматриваемый метод основан на серьезном допущении: Р и Q константны на протяжении периода исследований. Еще одно важное допущение (на котором также основаны и все современные стохастические модели мечения и повторного отлова) - эмиграция и иммиграция особей в популяции необратимы. В ситуациях, когда особи в одни годы могут покидать исследуемую территорию, а в другие возвращаться обратно, оценка сохраняемости будет искажаться в сторону завышения.
В рамках принятых допущений справедливо, что если в год z мы окольцевали N особей, то ожидаемое количество сохранившихся на изучаемой территории меченых птиц на следующий год составит NQ, а количество повторно пойманных (или иным способом зарегистрированных) - NQP. На второй после кольцевания год на данной территории останется N^ меченых особей. Из них ожидаемое количество пойманных составит N^P. При этом NQ^ особей повторно пойманы как в первый, так и второй годы, a N^P(l - Р) - только во второй. Через к лет после кольцевания на изучаемой территории останется N^ окольцованных в год z особей, а отловить удасться N^P, из них N^P(\ - Р)^1 будут пойманы впервые.
Если исходить из предположения о константности Q и Р, то Q можно оценить по отношению п^ к щ\ п._№2Р пх NQP
При небольшом материале и малой вероятности отлова (встречи) меченых птиц может случайно оказаться, что zz2 > «ь Тогда оценка будет превышать 100%, что биологически бессмысленно. Это может произойти и в случае, если во второй после кольцевания год поиск окольцованных птиц велся более тщательно, чем в первый, а также в случае, если в популяции процесс эмиграции-иммиграции обратим. В последней ситуации методы оценки численности и сохраняемости с помощью мечения и повторного отлова вообще неприменимы, а понятие сохраняемости теряет смысл. В первых двух ситуациях, если нет возможности продолжить работу и дополнить материал, можно попробовать дать прикидочную оценку Q следующим вариантом метода:
V
Если работа ведется в течение многих лет, то для оценки усредненного значения Q можно использовать уравнение:
iVW^+nt _ NQ'P+NQ‘P+...+NQLP _ NQ2P(1 +Q+...+Q1"1) _
«,+«,+...+«,_,” NQP + NQ1P+...+NQ‘"’P~ NQPh + Q+...+Q1 ’) ^
При этом в анализ включаются особи, окольцованные не позже, чем за к лет до окончания работы на данной территории. В случае, например, если особь повторно зарегистрирована в годы (z + 1) и (z + 3), ее нельзя включать в число птиц, зарегистрированных в год (z + 2), хотя она, по всей видимости, и тогда присутствовала на участке исследования.
Поскольку данный метод исходит из константности Q и Р, вполне допустимо объединение выборок птиц, окольцованных в разные годы.
В качестве примера приведем расчеты Q по материалам кольцевания Fringilla coelebs на Куршской косе в 1957-1985 гг. (из: Бардин 1990). В эти годы помечено 3179 взрослых особей, ранее уже отмечавшихся и окольцованных на исследуемой территории площадью 300 га. До 1979 г. помечены 2294 особи. Из них в год (z + 1) повторно поймано 268 птиц, (z + 2) - 143, (z + 3) - 91, (z + 4) - 43, (z + 5) -16, (z + 6) - 8. Средняя оценка сохраняемости с помощью стохастической модели Джолли-Себера (Jolly 1965, 1982; Seber 1965, 1982) составила 0.53 (Бардин 1990). Попробуем оценить Q описанными выше вариантами метода:
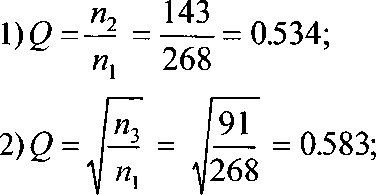
п2 + п3 + и4 + п5 + и6 пх + п2 + п3 + п^ + п5
143 + 91 + 43 + 16 + 8 _301_ 053?
268 + 143 + 91 + 43 + 16 ”561“ ’ ’
Вопроса о стандартном отклонении точечной оценки параметра Q и построении его доверительного интервала мы намерено не касаемся, поскольку столь грубый метод имеет смысл использовать лишь для выборок, не удовлетворяющих условиям применения стохастических моделей мечения и повторного отлова. Для сравнения одно родности частот щ, и2, •••, «к в разных выборках можно использовать критерий хи-квадрат или G-критерий. Однако, различия между частотами в такой таблице не однозначно свидетельствуют о различиях именно в сохраняемости; они могут быть обусловлены другими причинами. Следует добавить, что приводимые в некоторых публикациях стандартные ошибки процента возврата имеют лишь декоративное значение.
Представление первичного материала удобно делать в форме т.н. таблицы Лесли (для примера см. таблицу).
Пример составления таблицы Лесли.
Данные мечения и повторного отлова Parus major в дер. Малая Пачковка (Псковская обл.) в зимний период
|
Годы предыдущей регистрации меченых особей |
Годы кольцевания и контроля |
|||||
|
1972 |
1973 |
1974 |
1975 |
1976 |
1977 |
|
|
1972 |
5 |
3 |
— |
— |
1 |
|
|
1973 |
ЖЯМ |
18 |
8 |
2 |
— |
|
|
1974 |
имя |
ЖЖЖ |
ЯвИ |
36 |
11 |
1 |
|
1975 |
23 |
9 |
||||
|
1976 |
1ЖМ8 |
19 |
||||
|
Общее число пойманных |
20 |
63 |
147 |
86 |
107 |
82 |
|
Число повторных поимок |
0 |
5 |
36 |
44 |
36 |
30 |
|
Число впервые помеченых |
20 |
58 |
71 |
42 |
29 |
52 |
По данным из таблицы вычислим Q по первому уравнению:
3 + 8 + 11 + 9..... = 31=038 и, 5 + 18 + 36 + 23 82
Метод Q выгодно отличается от метода вычисления процента возврата также тем, что в расчеты не включается общее количество окольцованных птиц N. Число N часто дезориентирует орнитолога, т.к. создает впечатление, что сколь угодно высокую “точность” можно получить, просто увеличивая количество окольцованных птиц. Это позволяет не особенно утруждать себя тщательными поисками ранее помеченых. Между тем очевидно, что материалом для оценки сохраняемости является количество повторных отловов или встреч индивидуально меченых особей. Общее количество окольцо-ваных птиц важно лишь постольку, поскольку увеличивает количество особей, регистрируемых повторно. Поэтому основные усилия желательно направлять на обеспечение максимальной вероятности контроля уже маркированных птиц. Это достигается как усилением интенсивности направленного поиска на участке исследования, так и увеличением площади обследуемой территории.
В заключение следует подчеркнуть, что сохраняемость есть демографический параметр совокупности особей, определенным образом распределенных в пространстве. Поэтому величину сохраняемости (возвращаемости) следует приводить только с указанием величины контролируемой территории. Размеры изучаемого участка нужно выбирать, учитывая размеры участков обитания особей исследуемого вида. Желательно, чтобы площадь исследуемого участка более, чем в 100 раз превосходила среднюю площадь участков обитания птиц в данной местности.
