Перекисное окисление липидов дикорастущих галофитов в условиях Приэльтонья

Автор: Нестеров Виктор Николаевич, Богданова Елена Сергеевна, Табаленкова Галина Николаевна, Розенцвет Ольга Анатольевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Проблемы прикладной экологии
Статья в выпуске: 5 т.16, 2014 года.
Бесплатный доступ
Изучали интенсивность процессов перекисного окисления липидов (ПОЛ) - как одного из универсальных механизмов регуляции внутриклеточного метаболизма, у дикорастущих галофитов с разной стратегией соленакопления. Установлено, что для крино- и гликогалофитов процессы ПОЛ играют более существенную роль в сравнении с эугалофитами, исходя из большего количества конечных продуктов окисления (МДА). Однако у этих же типов растений выявлена и большая активность антиокислительной защиты.
Галофиты, перекисное окисление липидов, мембраны
Короткий адрес: https://sciup.org/148203314
IDR: 148203314 | УДК: 581.1
Lipid peroxidation of wild-growing halophytes in the conditions of the Prieltonye
Study of processes of the lipid peroxidation - one of universal mechanisms of regulation of an intracellular metabolism a wild-growing halophytes with different strategy of a salt tolerance. It is established that for crino-and glycohalophyte processes of lipid oxidation play more essential role in comparison with euhalophyte, proceeding from bigger quantity of the final products of oxidation (MDA). However at the same types of plants also big activity of anti-oxidizing protection is revealed.
Текст научной статьи Перекисное окисление липидов дикорастущих галофитов в условиях Приэльтонья
По имеющимся оценкам одной из общих тенденций изменения биосферы является аридизация климата и расширение площади пустынных территорий, сопровождаемых, в том числе, усилением засоления почвы [7]. В этой связи засоление почв является экономической и экологической проблемой многих стран мира [17]. Засоленные в разной степени почвы и солончаки разного генезиса являются обычными местообитаниями растений галофитов. Поиск потенциально солевыносливых, наиболее продуктивных природных галофитов в различных районах мира с целью их культивирования в природоохранных, хозяйственных, медицинских интересах становится важным направлением в современной экологии, физиологии и биохимии растений [12, 13, 16].
Известно, что галофиты в зависимости от устойчивости к засолению разделяются на три группы: эугалофиты – накапливающие в своих органах соли, криногалофиты – выделяющие избыток соли на поверхность листьев и гликогалофиты – исключающие проникновение солей в надземные органы растений [3]. Некоторые исследователи выделяют еще одну группу галофитов – гиперэугалофиты [17]. Растения в этих группах используют различные механизмы адаптации в одних и тех же условиях произрастания.
Устойчивость растений к почвенному засолению обеспечивается следующими приспособлениями: избирательное накопление или исключение ионов; контроль ионного поглощения корнями и транспорта в листья; компартментализация ионов на уровне клетки и целого растения; синтез совместимых растворенных веществ; изменение пути фотосинтеза; изменение в структуре мем-
бран; индукция антиоксидантных ферментов; индукция гормонов растений [5, 18].
Одним из универсальных механизмов управления внутриклеточным метаболизмом является перекисное окисление липидов (ПОЛ), индуцированное образованием активных форм кислорода [4]. Известно, что в нормальных условиях жизнедеятельности клетки постоянно присутствует определенный уровень ПОЛ, благодаря многоуровневой антиоксидантной системе защиты [14]. Усиление этого процесса ведет к образованию избыточного количества свободных радикалов, что нарушает состояние клеточных мембран и коллоидное состояние протоплазмы [2]. Поэтому при стимуляции процессов ПОЛ в мембранах уменьшается содержание липидов, а также меняются их микровязкость и электростатический заряд. При более глубоком окислении нарушается структура липидного бислоя и появляются дефектные зоны в мембранах клеток, что нарушает не только барьерную функцию мембран, но и функциональную активность мембранносвязанных белков.
Малоновый диальдегид (МДА) является одним из конечных продуктов окисления липидов [15], а его содержание – интегральной характеристикой соотношения процессов анаболизма и катаболизма биополимеров [10]. Изменения, вызванные продуктами ПОЛ, могут рассматриваться как основа для последующих восстановительных процессов [11].
В клетках растений окислительным превращениям подвергаются полиненасыщенные жирные кислоты полярных глико- (ГЛ) и фосфолипидов (ФЛ), стерины, которые являются основными компонентами клеточных мембран, а также нейтральные липиды [2].
Цель работы – оценить интенсивность процессов ПОЛ дикорастущих галофитов, различающихся по стратегии соленакопления солей.
МАТЕРИАЛ И МЕТОДИКА
Район исследования расположен на территории Волгоградской области, характеризуется бли- зостью залегания грунтовых вод, засоленностью почвогрунтов, что обуславливает формирование солончаковости и солонцеватости почв и определенного типа растительности [1].
Растительный материал отбирали в первой половине дня в середине июня 2013 г. на 2 площадках (20 х 20 м), расположенных в устьевых участках рек Большая Смарогда (биотоп 1) (49°07´с.ш., 46°50´в.д.) и Чернавка (биотоп 2) (49°12´с.ш., 44°40´в.д.).
Объектами исследования были выбраны представители эугалофитов – однолетники Salicornia perennans Willd. (Sp), Suaeda linifolia Pall. (Sl), полукустарничек Halocnemum strobilaceum (Pall.) Bieb. (Hs) из семейства Chenopodiaceae , кринога-лофитов – травянистый многолетник Limonium gmelinii (Willd.) O. Kuntze (Lg) из семейства Plumbaginaceae , гликогалофитов – полукустарничек Artemisia santonica L. (As) из семейства Aste-raceae .
Особенности отбора растений, анализа почвы на содержание солей, а так же методы экстракции, идентификации, анализа липидов и белков в растительном материале описаны ранее [8, 9]. Тип засоления почвы определялся по составу анионов в специализированной лаборатории ФГБУ «САС «Самарская»».
Интенсивность ПОЛ в листьях и корнях растений определяли по накоплению в МДА, определяемого по цветной реакции с тиобарбитуровой кислотой [6].
Активность супероксиддисимутазы (СОД) определяли согласно рекомендациям, описанным в работе [14].
В таблицах и рисунках использованы средние арифметические значения из трех независимых биологических проб для каждого биотопа. Статистическую обработку результатов анализов проводили с использованием программ Statistica 6.0 for Windows, Microsoft Excel 2007 и Statgraphics Centurion XVI.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Исследованные растения в систематическом плане представляют три семейства, в которых реализованы три стратегии эу-, крино- и глико-галотолерантности. В условиях Приэльтонья растения произрастали в двух биотопах, различающихся по эдафическим характеристикам. Особенностью территории является наличие слабощелочных почв с хлоридно-сульфатным типом засоления. Но общее содержание солей в почве прикорневой зоны исследуемых растений в биотопе 1 составило 0,92 мг/мл почвенной вытяжки, для биотопа 2 – 0,49. Однако в биотопе 2 содержание сульфат аниона составляло около 2 ммоль/л, в то время как в биотопе 1 – 0,3 ммоль/л.
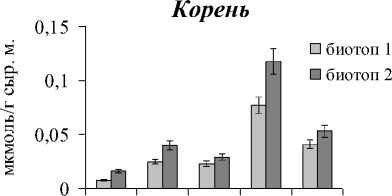
Содержание МДА в листьях варьировало от 0,014 до 0,085, в корнях – от 0,008 до 0,077 мкмоль/г сыр.м.. Причем как в листьях, так и корнях крино- и гликогалофитов уровень процессов ПОЛ был в 2 и более раз выше, чем в эугало-фитах. Кроме того, для крино- и гликогалофитов выявлено достоверно более высокое содержание МДА в растениях, произрастающих в биотопе 2, что может быть связано с более высоким содержанием сульфатов в почве.
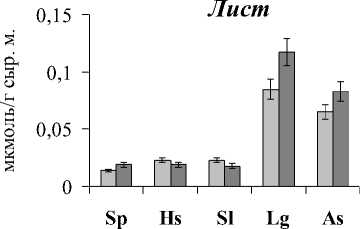
Рис. 1 . Содержание МДА в листьях и корнях дикорастущих галофитов. Обозначения: Sp - Salicornia perennans , Sl - Suaeda linifolia, Hs - Halocnemum strobilaceum, Lg - Limonium gmelinii, As - Artemisia santonica .
Sp Hs Sl Lg As
Поскольку основной вклад в структуру мембран вносят ГЛ и ФЛ, уровень процессов ПОЛ оценивался именно в этих группах липидов. Данные табл. показывают, что максимальное содержание мембранных липидов у растений в обоих биотопах наблюдали у крино- и гликогалофитов. Например, для L. gmelinii и A. santonica произрастающих в устье Б. Сморогды (биотоп 1) содержание ГЛ в листьях составило 7,4 и 7,6 мг/г сыр. м., соответственно, а для S. perennans, H. strobila- ceum и S. linifolia – 2,3, 3,4 и 2,4, соответственно. Для листьев L. gmelinii отмечено также и большее в сравнении с другими видами содержание ФЛ (2,6 мг/г сыр. м.). Аналогичная тенденция обнаружена и в суммарном содержании белков (СБ): в листьях растений биотопа 2 содержание белков было выше, чем в растениях биотопа 1. В подземной части большинства исследуемых видов растений содержалось меньше и липидов и белков. Исключением стали корни L. gmelinii, в которых обнаружено существенно более высокое галофитов содержание липидов и белков в 2-3
содержание СБ. Что касается количественных раза ниже, чем у крино- и гликогалофитов.
показателей в разных группах растений, то у эу-
Таблица. Содержание липидов и белков в листьях и корнях дикорастущих галофитов, мг/г сыр. м
|
Биотоп 1 |
Биотоп 2 |
|||||
|
ГЛ |
ФЛ 1 |
СБ |
ГЛ |
ФЛ 1 |
СБ |
|
|
Лист |
||||||
|
Sp |
2,3 |
0,8 |
4,3 |
2,6 |
1 |
6,4 |
|
Hs |
3,4 |
1,1 |
10,1 |
2,7 |
0,7 |
11,0 |
|
Sl |
2,4 |
1,0 |
8,8 |
1,8 |
0,6 |
10,0 |
|
Lg |
7,4 |
2,6 |
14,6 |
6,5 |
2,3 |
20,5 |
|
As |
7,6 |
1,8 |
109,0 |
6,3 |
1,8 |
121,0 |
|
Корень |
||||||
|
Sp |
0,1 |
0,7 |
2,5 |
0,1 |
1,1 |
4,6 |
|
Hs |
0,1 |
0,8 |
8,7 |
0,1 |
0,9 |
9,3 |
|
Sl |
0,1 |
0,8 |
7,6 |
0,2 |
1,0 |
9,0 |
|
Lg |
0,2 |
0,7 |
200,0 |
0,2 |
0,9 |
197,5 |
|
As |
0,1 |
0,9 |
60,0 |
0,1 |
1,0 |
47,6 |
Примечание: стандартная ошибка составляет не более 10%.
Расчет содержания разных типов мембранных липидов на количество МДА показал идентичную картину для ГЛ (ГЛ/МДА) и ФЛ (ФЛ/МДА), что говорит о том, что эти липиды в равной степени подвержены окислительным процессам. Анализ полученных данных показал, что самое высокое отношение липиды/МДА наблюдалось в листьях и корнях растений эугалофита S. perennans, произрастающих в биотопе 1 (рис. 2), а более низкое – в листьях и корнях криногалофита L. gmelinii и гликогалофита A. santonica в биотопе 2.
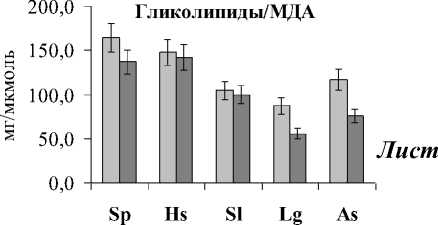
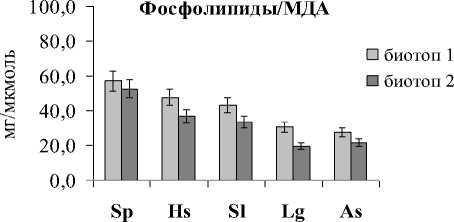
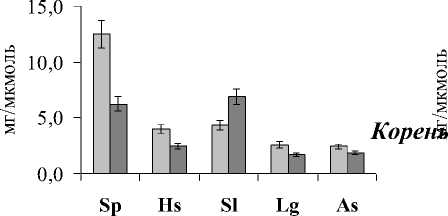
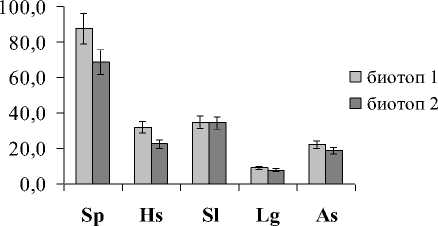
Рис. 2 . Отношение липиды/МДА в листьях и корнях галофитов. Обозначения как на рис. 1.
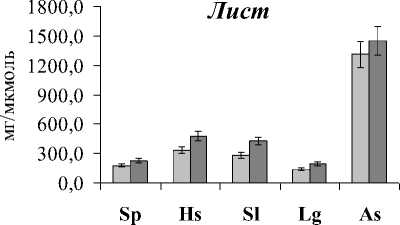
Выявленная закономерность позволяет предположить в сочетании с данными по уровню МДА, что клеточные мембраны эугалофитов более стабильны, в меньшей степени подвержены окислительным процессам, чем крино- и гликогалофитов. Анализ отношения ВБ/МДА показал прямо противоположную картину в сравнении с показателем липиды/МДА в листьях и корнях исследуемых растений (рис. 3). Отношение ВБ/МДА в целом было наибольшим у A. santonica и L. gmelinii, а наименьшим – у S. perennans. Об- щеизвестно, что ВБ растений, как правило, состоят из нескольких компонентов с антиокислитель-ной активностью (амилазной, инвертазной, каталазной, пероксидазной, фитазной, дезоксирибо-нуклеазной и цистинредуктазной), т.е. могут защищать клетки, органы и ткани от свободных радикалов и продуктов ПОЛ. Как видно в исследованных видах растений при большей деградации липидов происходит более активный синтез ВБ в листьях и корнях. Это означает, что у кри-но- и гликогалофитов более эффективна система антиоксидантной защиты. Причем у растений биотопа 2 степень окисления ГЛ и ФЛ была выше, чем биотопа 1, поэтому в листьях растений, произрастающих в переделах биотопа 2, отношение ВБ/МДА было выше для всех исследуемых видов.
3000,0 2500,0 2000,0
1500,0
1000,0
500,0 0,0
Корень
Sp Hs Sl Lg As
Рис. 3 . Отношение Водорастворимые белки/МДА в листьях и корнях галофитов.
Обозначения как на рис. 1
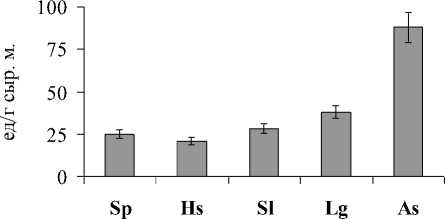
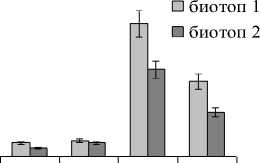
Рис. 4 . Активность СОД в листьях дикорастущих галофитов. Обозначения как на рис. 1.
Данные об активности одного из антиокисли-тельных ферментов супероксиддисмутазы (СОД) показывают, что увеличение активности СОД коррелировало с увеличением содержания МДА (рис. 4). Это подтверждает компенсаторную роль водорастворимых белков в ответ на активацию процессов ПОЛ, характерную для крино- и гликогалофитов.
Таким образом, исследованные дикорастущие виды галофитов, произрастающие в условиях Приэльтонья, различаются не только по своему систематическому положению, стратегии солена-копления, но и содержанию мембранных липидов, мембранносвязанных и цитозольных белков, а также уровню процессов ПОЛ. Для солевыде-лющих и ограничивающих поступление солей в надземную часть галофитов процессы ПОЛ играют более существенную роль, исходя из большего количества конечных продуктов окисления (МДА). Однако у этих же типов растений выявлена и большая активность антиокислительной защиты.
Работа выполнена при поддержке гранта РФФИ № 12-04-01110-а. Авторы выражают признательность к.б.н. Лысенко Т.М. за определение видов галофитов.
Список литературы Перекисное окисление липидов дикорастущих галофитов в условиях Приэльтонья
- Атлас почв СССР/под ред. И.С. Кауричева, И.Д. Громыко. М.: Колос, 1974. 168 с.
- Владимиров Ю.А., Арчаков А.И. Перекисное окисление липидов в биологических мембранах.М.: Наука, 1972. 252 с.
- Генкель П.А. Физиология жаро-и засухоустойчивости растений. М.: Наука, 1982. 280 с.
- Гонтарь О.Б., Жиров В.К., Хаитбаев А.Х., Говорова А.Ф. Возрастные аспекты адаптации растений в экстремальных условиях//Вестник МГТУ. 2006. Т. 9. № 5. С. 729-734.
- Креславский В.Д., Карпентиер Р., Климов В.В., Мурата Н., Аллахвердиев С.И. Молекулярные механизмы устойчивости фотосинтетического аппарата к стрессу//Биологические мембраны. 2007. Т. 24. № 3. С. 195-217.
- Лукаткин А.С., Голованова В.С. Интенсивность перекисного окисления липидов в охлажденных листьях теплолюбивых растений//Физиология растений. 1988. Т. 35. Вып. 4. С. 773-780.
- Пьянков В.И., Мокроносов А.Т. Основные тенденции изменения растительности земли в связи с глобальным потеплением климата//Физиология растений. 1993. Т. 40. № 4. С 515-531.
- Розенцвет О.А., Нестеров В.Н., Богданова Е.С., Лысенко Т.М. Липиды Suaeda Salsa (L.) Pall.: содержание, динамика, вариабельность в естественных условиях произрастания вида//Изв. Самар. НЦ РАН. 2013. Т. 15. № 3. С. 145-150.
- Розенцвет О.А., Нестеров В.Н., Богданова Е.С. Состав мембран дикорастущих галофитов с различными механизмами регуляции солевого обмена в зависимости от абиотических факторов среды//Биологические мембраны. 2014. Т. 31. № 2. С. 137-146.
- Тарчевский И.А. Метаболизм растений при стрессе (избранные труды). Казань: Фэн, 2001. 448 с.
- Хочачка П., Сомеро Д. Биохимическая адаптация. М.: Мир, 1988. 568 с.
- Шамсутдинов З.Ш., Савченко И.В., Шамсутдинов Н.З. Галофиты России, их экологическая оценка и использование. М.: ООО Эдель-М, 2001. 399 с.
- Ahmad P., Prasad M.N.V. Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change. Springer Science+Business Media, New York, 2012. 515 p.
- Frankel E.N. Lipid oxidation. The oily press ltd, Glasgow, 1998. pp. 79-99.
- Labudda M. Lipid peroxidation as a biochemical marker for oxidative stress during drought. An effective tool for plant breeding//E-wydawnictwo, Poland. 2013. Http://www.e-wydawnictwo.eu/Document/Document Preview/3342. P.1-12.
- Lambers H. Dry land salinity: A key environmental issue in southern Australia//Plant Soil. 2003. V. 257. P. 5-7.
- Mamedov E.Y., Esenov P.E., Durikov M.K., Zverev N.E., Tsukanova S.K. Experience of Halophyte cultivation on saline soils. Ashgabat, 2009. 44 p.
- Parida A.K., Das A.B. Salt tolerance and salinity effects on plants: a review//Ecotoxicol. Environ. Saf. 2005. V. 60. P. 324-349.
