Первые сведения о мшанках (Bryozoa) литорали полуострова Кони, Охотское море
 литорали полуострова Кони, Охотское море")
Автор: Грищенко А.В.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Зоология
Статья в выпуске: 4, 2014 года.
Бесплатный доступ
При исследовании коллекции мшанок (Bryozoa), собранной в среднем горизонте трёх участков скалисто-валунной литорали северо-западного побережья полуострова Кони (Тауйская губа Охотского моря), обнаружено десять видов мшанок, в т.ч. семь видов - впервые отмечены для литорали северного побережья Охотского моря, а пять - новые для фауны Охотского моря. По численности и площади проективного покрытия субстратов доминируют Porella alba Nordgaard, 1906, а также три субдоминатных вида - Cauloramphus multispinosus Grischenko, Dick et Mawatari, 2007, Electra asiatica Grischenko, Dick et Mawatari, 2007 и Cauloramphus magnus Dick et Ross, 1988. Все найденные виды имеют инкрустирующие колонии и встречены на твёрдых субстратах. Фауна представлена комплексом бореально-арктических и бореальных элементов. Низкое видовое богатство литорального таксоцена, сочетающееся с высокой численностью массовых видов, является спецификой локальной бриозоофауны и указывает на высокую динамику элиминации колоний мшанок под воздействием стрессоген-ных факторов в зимний период.
Мшанки, литораль, полуостров кони, охотское море
Короткий адрес: https://sciup.org/147204692
IDR: 147204692 | УДК: 594.7
First data on intertidal Bryozoa of the Koni peninsula, Sea of Okhotsk
Examination of the bryozoan collection gathered from three sites within the middle horizon of rocky-boulder intertidal along northwestern shoreline of the Koni Peninsula (Tauyskaya Inlet, Sea of Okhotsk), has revealed ten species. Seven species have been registered for the first time for the northern coast of the Sea of Okhotsk, and five species are new to the bryozoan fauna of this sea. The most abundant are Porella alba Nordgaard, 1906, Cauloramphus multispinosus Grischenko, Dick et Mawatari, 2007, Electra asiatica Grischenko, Dick et Mawatari, 2007 and Cauloramphus magnus Dick et Ross, 1988. All species form encrusting colonies on the hard substrata (rock, pebbles, small boulders). Biogeographically, this fauna represented equally by Boreal-Arctic and Boreal elements. Low overall species diversity of intertidal taxocene combined with a high abundance of the mass species seem to be specific peculiarity for local bryozoan fauna, and is indicating a high dynamics of elimination of colonies as affected by environmental stressors during winter period.
Текст научной статьи Первые сведения о мшанках (Bryozoa) литорали полуострова Кони, Охотское море
Информация о фауне мшанок приливно-отливной полосы Охотского моря остаётся во многом поверхностной и фрагментарной. В то время как данные по охотоморской фауне Bryozoa литорали Курильских о-вов опубликованы в нескольких фаунистических обзорах [Гонтарь, 1978а, б, 1980; Gontar, 1981], а также специализированных исследованиях [Изюмова и др. 1974; Kubanin, 1996; Кубанин, 1997], сведения по континентальному побережью, особенно его северному сектору, до сих пор крайне скудны. Е.А. Изюмова [1977] отметила 7 видов хейлостомат для всего Западно-Камчатского побережья. В единственной узкоспециализированной работе А.А. Кубанин [1976] описал 7 видов (все из отр. Cheilostomata) с литорали о. Завьялова (Тауйская губа), в т.ч. один вид, Cryptosula zavjalovensis, новый для науки. В обобщающей публикации по фауне мшанок литорали дальневосточных морей России А.А. Кубанин [1997] указал лишь 16 видов (4 из отр. Ctenostomata и 12 из отр. Cheilostomata) для всего северного и северовосточного побережья Охотского моря (включая югозападную Камчатку, Пенжинскую, Гижигинскую, Ямскую и Тауйскую губы, а также Охотск и Аян). Более поздние сведения [Регель, 2005] о нахождении мшанок на литорали Тауйской губы заимствованы из работ А.В, Ильичева и И.А. Рязанцевой [1992], а также А.А. Кубанина [1997], и не содержат новых данных.
Западное (тауйское) побережье п-ва Кони территориально принадлежит Ольскому участку Магаданского государственного заповедника [Государственный…]. Литоральная зона западного побережья п-ва Кони относится ко 2-му био-номическому типу (открытый морской берег) и представлена плоскими скалистыми платформами, шириной до 120 м, чередующимися с поясами гальки, валунов и глыб [Спиридонов, 2013]. Амплитуда приливно-отливной динамимки достигает 4–5 м [Зуенко, 1992].
В августе 2013 г. экспедицией Института океанологии им. П.П. Ширшова РАН, под руководством д.б.н. В.А. Спиридонова, были проведены первые хорологические исследования литорали западного (тауйского) побережья п-ва Кони, в ходе которых осуществлён сбор материалов по мшанкам. В настоящей статье описаны результаты исследования собранной коллекции.
Материалы и методы
Материалы собраны В.А. Спиридоновым и Н.Ю. Неретиным в августе 2013 г. на трёх участках литорали вдоль западного побережья п-ва Ко-
ни (рисунок) в ходе исследований, проведённых с целью описания литоральных фаций и сообществ. Сбор осуществлялся с помощью закладки перпендикулярных берегу трансект, вдоль которых производилось разбиение и съемка профиля берега и дна, описание субстратов, микрорельефа и литорального населения. Плановая привязка начальной и конечной точек профиля осуществлялась с помощью GPS-приемника Garmin Dakota 20. Температура и соленость определялись с помощью анализатора воды ProfiLine Cond 3110. Колонии мшанок были собраны на субстратах: гальке, камнях, мелких валунах и скальных обломках, обнаруженных в расщелинах скал и литоральных ваннах, а также лежащих в подстилающем слое валунного и глыбового навала. Данные станций:
4–2. 31.08.2013, район м. Плоский, 59°9'9.0"с.ш., 151°37'13.6" в.д. – 59°9'9.3" с.ш.,
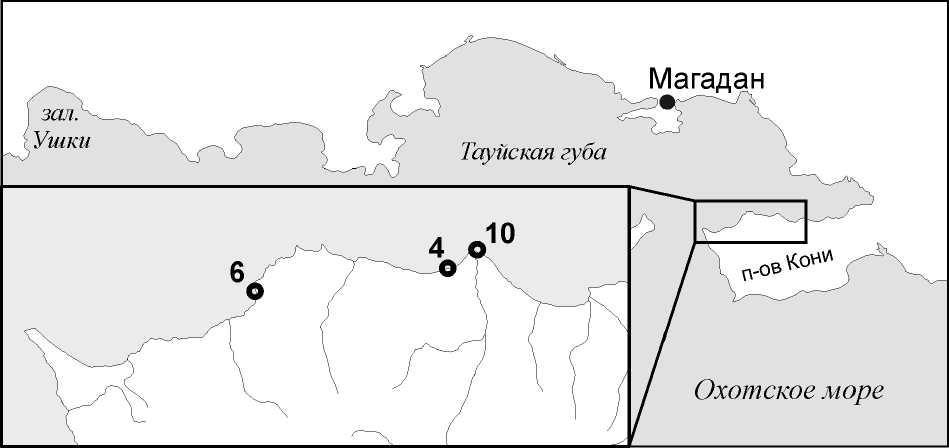
Тауйская губа Охотского моря и северо-западное побережье п-ва Кони с указанием районов сборов
Фиксация материалов осуществлялась путём высушивания. Колонии мшанок были найдены на 29 единицах субстратов (галька, камни, мелкие валуны, скальные обломки). Для идентификации видов использовался бинокуляр МБС–10. Всего исследовано 625 экземпляров (колоний или их фрагментов). Обработанный материал в настоящее время находится на хранении в музее зоологии бесповоночных им. Д.М. Федотова и Д.Е. Харитонова кафедры зоологии беспозвоночных и водной экологии биологического факультета Пермского государственного национального исследовательского университета. В дальнейшем материал будет передан для регистрации и постоянного хранения в Зоомузей МГУ.
Результаты
В приводимом ниже аннотированном таксономическом списке синонимия ограничена и приводится только для видов, ранее отмеченных для северного и северо-восточного побережья Охотского
151°37'13.6" в.д.; большая вода: 03:24/16:00, 4.0/3.6 м; малая вода: 9:48/ 21:54, 2.0/2.3 м; температура у уреза воды: 5.6°С; солёность: 31.2‰; время выполнения: 9:00–12:00. Средний горизонт скалисто-валунной литорали.
6–3. 02.08.2013, район м. Скалистый, 59°8'3.90"с.ш., 151°23'4.60" в.д. – 59°8'3.70" с.ш., 151°23'2.50" в.д.; большая вода: 04:24/16:42, 4.0/3.7 м; малая вода: 10:42/ 22:48, 2.2/2.1 м; температура у уреза воды: 12.2°С; солёность: 28.4‰; время выполнения: 10:00–13:00. Средний горизонт валунной литорали.
10–5. 05.08.2013, район м. Плоский, 59°9'37.90"с.ш., 151°39'42.90" в.д. – 59°9'39.60" с.ш., 151°39'47.10" в.д.; большая вода: 07:06/20:36, 4.0/3.7 м; малая вода: 01:00/ 14:00, 2.5/1.6 м; температура у уреза воды: 14.0°С; солёность: 26.5‰; время выполнения: 13:40–17:00. Средний горизонт валунной литорали.
моря. Виды, впервые обнаруженные в этом районе, отмечены знаком (*).
Отряд Ctenostomata Busk, 1852
Подотряд Euctenostomata Jebram, 1973
Надсемейство Alcyonidioidea Johnston, 1847
Семейство Alcyonidiidae Hincks, 1880
Род Alcyonidium Lamouroux, 1813
Alcyonidium mytili Dalyell, 1847
Alcyonidium mytili : Кубанин, 1997, с. 120.
Материал. 52 экз.
Экология. В основном материал представлен мелко-размерными колониями (порядка 0.5 × 0.5 – 1 × 1 см). Эпизодически встречающиеся крупные колонии формируют обширный, непрерывный покров субстрата (достигающий размеров 10 × 12 см) в эпибиозах скал и мелких валунов.
Распространение. Бореально-арктический циркумполярный вид. Неоднократно отмечался в Беринговом и Охотском морях [Клюге, 1953, 1961, 1962], в т.ч. для литорали северного и северо-восточного побережий Охотского моря [Кубанин, 1997].
Отряд Cheilostomata Busk, 1852
Подотряд Malacostegina Levinsen, 1902 Надсемейство Membraniporoidea Busk, 1852 Семейство Electridae d'Orbigny, 1851 Род Electra Lamouroux, 1816
Electra asiatica Grischenko, Dick et Mawatari, 2007*
Electra crustulenta var. baltica : Кубанин, 1976, с. 31.
Electra baltica : Кубанин, 1997, с. 121.
Материал. 116 экз.
Экология. Массовый вид. Колонии с зооидами, расположенными в радиально расходящиеся сегменты, формируют пятна неправильной округлой формы (преимущественно до 1 см, редко до 3 см в диаметре), которые ввиду их близкого расположения сливаются и т.о. фрагментарно или полностью изолируют субстрат на площади в несколько десятков см2. Обильно представлен в эпибиозах гальки, а также на нижней поверхности камней и мелких валунов, лежащих поверх скал.
Распространение. Тихоокеанский азиатский широкобореальный вид. До уточнения видовой пренадлежности [Grischenko et al. 2007] был известен из литоральной зоны нескольких районов Дальневосточных морей России, в т.ч. северного побережья Охотского моря, под названием Electra baltica (Borg, 1931) [см. Кубанин, 1976, 1997].
Подотряд Neocheilostomina d’Hondt, 1985 Надсемейство Calloporoidea Norman, 1903 Семейство Calloporidae Norman, 1903 Род Cauloramphus Norman, 1903
Cauloramphus magnus Dick et Ross, 1988*
Материал. 113 экз.
Экология. Массовый вид. Многочисленные колонии округлой формы (в основном 0.3–0.5 см в диаметре, редко до 2 см) формируют пятнистый покров на поверхности гальки и крупных камней. Плотность поселений достигает 94 экз. на ~100 см2 поверхности.
Распространение. Тихоокеанский широкобореальный вид. Новый вид для фауны Охотского моря. Ранее был известен по первоописанию с литорали о. Кадьяк, залив Аляска [Dick et Ross, 1988], Командорских о-вов [Кубанин, 1997; Грищенко, 1997, 2004; Grischenko, 2002], из района Кетчикан, Аляска [Dick et al. 2005] и б. Аккеши, тихоокеанское побережье о. Хоккайдо [Grischenko et at. 2007]. По неопубликованным данным автора, C. magnus также зарегистрирован на литорали средних и южных Курильских о-вов, а также малой Курильской гряды (б. Броутона, о. Симушир; б. Консервная, о. Итуруп; б. Крабовая, о. Шикотан соответственно).
Cauloramphus multispinosus Grischenko, Dick et Mawatari, 2007*
Cauloramphus pseudospiniferum : Кубанин, 1976, с. 30, рис. А–Г.
Материал. 119 экз.
Экология. Массовый вид. Колонии в виде пятен неправильной формы (в основном до 1 см, редко до 3.5 см в длину) отмечены на нижней поверхности камней и мелких валунов, лежащих поверх скал.
Распространение. Тихоокеанский азиатский широкобореальный вид. Новый вид для фауны Охотского моря. Ранее был известен на литорали б. Аккеши, тихоокеанское побережье о. Хоккайдо [Grischenko et at. 2007], а также отмечен на литорали б. Консервная, охотоморское побережье о. Итуруп, южные Курильские о-ва (неопубликованные данные автора).
Род Septentriopora Kuklinski et Taylor, 2006
Septentriopora ? nigrans (Hincks, 1882)
Tegella aff. japonica : Кубанин, 1976, с. 32, рис. Д–Е.
Hincksina nigrans : Кубанин, 1997, с. 122.
Материал. 13 экз.
Замечания. По ряду ключевых таксономических признаков исследованные экземпляры близки к Septentriopora karasi Kuklinski et Taylor, 2006, а также S. denisenkoae Kuklinski et Taylor, 2006. Для уточнения видовой принадлежности необходимо дополнительное исследование с использованием сканирующей электронной микроскопии.
Экология. Небольшие корковые колонии (размером менее 1 × 1 см) найдены в эпибиозах камней.
Распространение. Бореально-арктический циркумполярный вид. Прежде отмечался во всех дальневосточных морях России [Клюге, 1961], в т.ч. для литорали северного и северо-восточного побережий Охотского моря [Кубанин, 1997].
Надсемейство Hippothooidea Busk, 1859
Семейство Hippothoidae Busk, 1859
Род Celleporella Gray, 1848
Celleporella ? reflexa Dick et Ross, 1988*
Материал. 1 экз.
Замечания. По ряду ключевых таксономических признаков изученный экземпляр близок к Celleporella nodasakae Dick, Grischenko et Mawatari, 2005. С целью уточнения видовой принадлежности требуется дополнительное исследование с применением сканирующей электронной микроскопии.
Экология. Обнаружен на скальном субстрате. Колония подвергалась абразии и была мертва на момент сбора.
Распространение. Тихоокеанский высокобореальный вид. Новый вид для фауны Охотского моря. До настоящего обнаружения был известен по первоописанию с литорали о. Кадьяк, залив Аляска [Dick et Ross, 1988], Командорских о-вов [Грищенко, 1997, 2004; Grischenko, 2002], из района Кетчикан, Аляска [Dick et al. 2005], а также б.
Датч-Харбор, о. Уналашка, Алеутские о-ва [Grischenko, 2002; Грищенко, Фостер, 2012].
Надсемейство Lepralielloidea Vigneaux, 1949
Семейство Bryocryptellidae Vigneaux, 1949
Род Porella Gray, 1848
Porella alba Nordgaard, 1906*
Материал. 159 экз.
Экология. Самый массовый вид. Корковые округлые колонии (в большинстве менее 1 см в диаметре, редко до 2.5 см) формируют на некоторых камнях непрерывное проективное покрытие субстрата благодаря высокой плотности поселений (до 32 экз. на ~100 см2 поверхности).
Распространение. Тихоокеанский высокобореальный вид. Новый вид для фауны Охотского моря. В регионе северо-западной Пацифики ранее был известен из района Командорских о-вов и вдоль юговосточного побережья Камчатки [Кубанин, 1997; Грищенко, 1997, 2004; Grischenko, 2002].
Семейство Umbonulidae Canu, 1904
Род Rhamphostomella von Lorenz, 1886
Rhamphostomella bilaminata (Hincks, 1877)*
Материал. 1 экз.
Экология. Обнаружен на скальном субстрате. Колония подвергалась абразии и была лишена живых зооидов на момент сбора.
Распространение. Бореально-арктический циркумполярный вид. В регионе дальневосточных морей России зарегистрирован в Беринговом проливе [Gontar, Denisenko, 1989], Авачинской губе [Гон-тарь, 1989; Grischenko, 2002], а также на шельфе западной Камчатки, Охотское море [Грищенко, 2001; Grischenko et al. 1999].
Надсемейство Schizoporelloidea Jullien, 1883
Семейство Stomachetosellidae Canu et Bassler, 1917
Род Stomachetosella Canu et Bassler, 1917
Stomachetosella producta (Packard, 1863)*
Материал. 1 экз.
Экология. Обнаружен на скальном субстрате. Колония подвергалась абразии и не содержала живых зооидов на момент сбора.
Распространение. Бореально-арктический циркумполярный вид. Новый вид для бриозоофауны Охотского моря. Для северо-западной Пацифики указывался для района о. Зелёный (малая Курильская гряда) под названием Pachyegis producta [см. Гонтарь, 1980, с. 11].
Семейство Cryptosulidae Vigneaux, 1949
Род Cryptosula Canu et Bassler, 1925
Cryptosula zavjalovensis Kubanin, 1976
Cryptosula zavjalovensis : Kubanin, 1976, с. 33, рис. Ж–З; 1997, с. 125.
Материал. 50 экз.
Экология. Корковые колонии с неровными округлыми контурами (достигающие размеров 4 × 5 см) создают толстые наросты на поверхности кам- ней и мелких валунов, эпизодически замуровывая трубки седентарных полихет.
Распространение. Тихоокеанский широкобореальный вид. Один из самых массовых видов на литорали многих районов всей Северной Пацифи-ки. В дальневосточных морях России отмечен в следующих районах: Берингово море – заливы Лаврентия, Анадырский, Олюторский, Корфа, Ка-рагинский, Камчатский, Кроноцкий, Авачинский, Командорские о-ва [Клюге, 1961; Кубанин, 1997; Грищенко, 1997, 2004]; Охотское море – о. Завьялова [Кубанин, 1976], залив Анива, южный Сахалин [Клюге и др., 1959], Курильские о-ва: Пара-мушир, Онекотан, Уруп, Итуруп, Кунашир, Шикотан [Клюге, 1961; Клюге и др., 1959; Гонтарь, 1980, 1992; Gontar, 1981; Кубанин, 1997]; Японское море – Татарский пролив, западное прибрежье южного Сахалина [Андросова, 1958; Клюге, 1961], о. Монерон [Кубанин, Тарасова, 1985], заливы Петра Великого, Восток и Посьета [Андросова, 1971; Кубанин, 1975, 1987, 1997, 1998].
В пределах среднего горизонта трёх участков литорали западного побережья п-ва Кони зарегистрированы 10 видов мшанок (1 из отр. Ctenostomata и 9 из отр. Cheilostomata) (табл. 1), в т.ч. семь видов – впервые отмечены для литорали северного побережья Охотского моря, а пять видов – новые для фауны Охотского моря. Все идентифицированные виды Bryozoa представлены инкрустирующими (корковыми) многорядными, либо однорядными ( C. ? reflexa ) колониями. Виды с вертикальной конструкцией колоний отсутствуют. Обнаруженные виды населяют исключительно жёсткие субстраты – гальку, камни, мелкие валуны и скальные обломки.
Таксономический портрет фауны и состав жизненных форм указывают на типичное сообщество об-растателей, характерное для эпифауны жёстких грунтов (валуны, глыбы и подстилающего их слоя камней и гальки) среднего горизонта литорали высокобо-рельной подзоны северо-западной Пацифики.
Ядро литорального таксоцена исследованного района составляют четыре массовых вида: P. alba , C. multispinosus , E. asiatica и C. magnus , превалирующие по численности и площади проективного покрытия субстратов. Характерно, что кроме мшанок, на исследованных скальных субстратах не было выявлено других сидячих форм зообентоса, за исключением мелких циррипедий и трубок спи-рорбид. Конкуренция за субстрат (отмеченные случаи взаимного, зачастую многослойного обрастания) таким образом происходит в основном между колониями одного или разных массовых видов Bryozoa.
Фауна представлена комплексом бореальноарктических (4 вида) и бореальных (6 видов) элементов, (в процентном соотношении как 40 : 60).
Бореальные элементы включают две группы: ти- океанские высокобореальные (2 вида).
хоокеанские широкобореальные (4 вида) и тихо-
Таблица 1
Обобщенные данные о видовом составе, количественном распределении на станциях, форме роста и биогеографии мшанок литорали западного побережья п-ва Кони
|
Виды |
Станции |
Всего экз. |
Форма роста ко-лоний1 |
Биогеографический 2 статус |
||
|
4–2 |
6–3 |
10–5 |
||||
|
Porella alba |
18 |
- |
141 |
159 |
К.М. |
Т, ВБ |
|
Cauloramphus multispinosus |
5 |
53 |
61 |
119 |
К.М. |
Т, ШБ |
|
Electra asiatica |
41 |
6 |
69 |
116 |
К.М. |
Т, ШБ |
|
Cauloramphus magnus |
103 |
10 |
- |
113 |
К.М. |
Т, ШБ |
|
Alcyonidium mytili |
7 |
- |
45 |
52 |
К.М. |
БА, Ц |
|
Cryptosula zavjalovensis |
4 |
2 |
44 |
50 |
К.М. |
Т, ШБ |
|
Septentriopora ? nigrans |
- |
- |
13 |
13 |
К.М. |
БА, Ц |
|
Celleporella ? reflexa |
1 |
- |
- |
1 |
К.О. |
Т, ВБ |
|
Stomachetosella producta |
1 |
- |
- |
1 |
К.М. |
БА, Ц |
|
Rhamphostomella bilaminata |
1 |
- |
- |
1 |
К.М. |
БА, Ц |
|
Кол-во экз. для станций Кол-во видов для станций |
181 9 |
71 4 |
373 6 |
625 10 |
||
1Форма роста колоний: К.М.: корковые, многорядные; К.О.: корковые, однорядные. 2Биогеографический статус для видов: БА, Ц: бореально-арктический, циркумполярный; Т, ШБ: тихоокеанский, широкобореальный; Т, ВБ: тихоокеанский, высокобореальный.
Обсуждение
Совокупность неблагоприятных факторов среды, специфичных для литоральной зоны (осушение, волновое воздействие, инсоляция, выпадение осадков, приводящих к локальному распреснению, и т.п.) в районе северного побережья Охотского моря, дополяется интенсивным истирающим воздействием ледяного покрова, который может задерживаться здесь до начала лета [Ландшафты… 2006; Ростов и др. 2003]. Продолжительное стрессогенное воздействие на литоральную бриозоофау-ну п-ва Кони наиболее явно отражено на размерновозрастной структуре популяций. Резкое преобладание небольших по размерам колоний, находящихся на ранних стадиях астогенеза, особенно в популяциях массовых видов, указывает на то, что в летний период происходит интенсивное оседание и прикрепление личинок – колонизация субстратов литоральной зоны. Малая доля обнаруженных крупных колоний, в свою очередь, позволяет предположить, что лишь незначительный процент мшанок из числа рекрутов успешно перезимовывает и вырастает на литорали до дефинитивных размеров, достигая зрелости. Множество колоний различных видов не выдерживают долгого стресса и погибают – главным образом из-за истирания и, как следствие, разрушения известковых скелетов. По нашим наблюдениям, не менее 30% исследованного материала (в т.ч. колонии трёх видов, известные по единичным экземплярам – C. ? reflexa ,
R. bilaminata и S. producta ), в той или ной мере подвергалось абразии. Вероятно, именно с высокими темпами элиминации сидячих форм зообентоса в зимний период связано скромное видовое богатство (10 видов) фауны мшанок литоральной зоны обследованного района. В ходе специализированного исследования фауны мшанок литорали о. Завьялова, расположенного в акватории Тауй-ской губы, на удалении 18 км к востоку от м. Таран п-ва Кони, также было обнаружено лишь семь видов хейлостомат [Кубанин, 1976]. Низкое видовое богатство, сочетающееся с высокой численностью монодоминантного, либо немногих субдоминантных видов, было также показано для побережий Гренладского моря [см. Kuklinski, 2009; Kuk-linski, Barnes, 2005a,b; Kuklinski, Taylor, 2006b] и, таким образом, видимо, является общей характерной особенностью литоральной бриозоофауны, обитающей в высоких широтах северного полушария.
Десять видов Bryozoa, найденные на литорали западного побережья п-ва Кони, с семью видами, впервые отмеченными для северного и северовосточного побережья Охотского моря, увеличивают до 23 общее количество видов, известных для этого района. Данные по видовому богатству фауны мшанок литорали различных районов Охотского моря контрастны (табл. 2). В то время как для Курильской островной гряды, протяжённостью около 1200 км, известны 38 видов, для всего сектора западного, северного и восточного побережий моря (>5000 км) обнаружены только 34 вида (включая семь, установленных настоящим исследованием). Показатели для западного и югозападного побережий выглядят ещё ниже. Не вызывает сомнений, что эти данные не отражают фактического богатства литоральной бриозоофау-ны региона. Как указывалось ранее [Грищенко, Фостер, 2012], до начала широкого внедрения «интенсивного подхода», многие литоральные сборы, проведённые вдоль побережий дальневосточных морей России и во всей Северной Пацифике на протяжении XX в. носили спорадический, зачастую случайный характер. Большинство хорологических исследований приливной зоны проведены без участия в них специалистов по мшанкам. Поэтому данные о видовом богатстве локальных литоральных фаун Bryozoa, известные для некоторых районов дальневосточных морей даже по узкоспециализированным работам [Андросова, Гостилов-ская, Изюмова, 1974; Изюмова, Кубанин, 1978; Кубанин, 1976, 1987, 1997; Kubanin, 1996; Грищенко, 2004], вероятнее всего, занижены.
Таблица 2
Сведения по видовому богатству мшанок литорали различных районов Охотского моря (по Кубанин [1997]) с включением данных настоящего исследования (*)
|
Районы Охотского моря |
Отряды Bryozoa |
Кол-во видов |
||
|
Cyclostomata |
Ctenostomata |
Cheilostomata |
||
|
Северное и северо-восточное побережье (юго-западная Камчатка, Пенжинская, Гижигинская, Ямская, Тауй-ская губы, Охотск, Аян) |
– |
4 |
19* |
23* |
|
Западное (Шантарские о-ва, Амурский лиман, побережье о. Сахалин на юг до м. Терпения) |
– |
1 |
6 |
7 |
|
Южное и юго-восточное побережье о. Сахалин (зал. Терпения и Анива) |
– |
3 |
19 |
13 |
|
Северные Курильские о-ва (Парамушир) |
– |
3 |
2 |
5 |
|
Средние Курильские о-ва (Уруп, Симушир, и др.) |
1 |
5 |
13 |
19 |
|
Южные Курильские о-ва (Итуруп, Кунашир, Шикотан, о-ва Хабомаи) |
2 |
10 |
12 |
24 |
Наши неопубликованные данные указывают на наличие неучтённых видов в локальных фаунах Bryozoa литорали Охотского моря. Так, идентификация материалов, собранных автором в сентябре 1992 г. на литорали о. Птичий, расположенного на шельфе западной Камчатки (57°10' с.ш., 156°35' в.д.), выявила 16 видов хейлостомных мшанок. Из них, по меньшей мере, шесть видов – новые для фауны литорали Охотского моря. Помимо того, материалы шести хорологических экспедиций, проведённых за период с 1987 по 2011 г. лабораторией хорологии Института биологии моря (ИБМ, г. Владивосток) на средние и южные Курильские о-ва, а также Малую Курильскую гряду, переданные сотрудником ИБМ А.П. Цурпало, включают 44 вида мшанок, среди которых более 10 – новые для фауны литорали Курильских о-вов, в т.ч. и для охотоморского побережья.
Перечисленные факты свидетельствуют о том, что фауна мшанок приливно-отливной полосы Охотского моря до сих пор находится на раннем этапе инвентаризации. Для выяснения достоверных показателей видового богатства локальных литоральных фаун Bryozoa последующим исследователям необходимо сочетать «интенсивный подход» [см. Dick, Ross, 1988; Грищенко, Фостер, 2012] с дальнейшим анализом образцов с использованием современных методик (прежде всего, сканирующей электронной микроскопии).
Автор признателен В.А. Спиридонову (ИОАН им. П.П. Ширшова) за предоставленный материал и данные сборов, А.В. Чернышёву (ИБМ ДВО РАН им. А.В. Жирмунского) и А.В. Ржавскому (ИПЭЭ РАН им. А.Н. Северцова) за критическое прочтение рукописи и замечания. А.А. Деркачёва (географический факультет МГУ) подготовила карту района сборов.
Настоящее исследование выполнено при финансовой поддержке Министерства образования и науки Российской федерации (основная часть, проект № 3487).
Список литературы Первые сведения о мшанках (Bryozoa) литорали полуострова Кони, Охотское море
- Андросова Е.И. Мшанки отряда Cheilostomata северной части Японского моря//Исслед. дальневосточных морей СССР. 1958. Т. 5. С. 90-205.
- Андросова Е.И. Мшанки залива Посьета Японского моря//Исслед. фауны морей. 1971. Т. 8 (16). С. 144-150.
- Андросова Е.И., Гостиловская М.Г., Изюмова Е.А. Тип Podaxonia, Класс Bryozoa. Список животных литорали Курильских островов//Растительный и животный мир литорали Курильских островов. Новосибирск: Наука, 1974. С. 368-369.
- Гонтарь В.И. Мшанки верхних отделов шельфа Курильских островов//Закономерности распределения и экология прибрежных биоценозов, Ленинград: Наука, 1978а. С. 60-62.
- Гонтарь В.И. Мшанки прибрежных вод острова Итуруп//Биол. моря. 1978б. № 1. С. 10-16.
- Гонтарь В.И. Фауна мшанок отряда Cheilostomata прибрежных вод Курильских островов//Ав-тореф. дис.. канд. биол. наук. Л., 1980. 24 с.
- Гонтарь В.И. Тип Bryozoa. Список макрофитов и беспозвоночных макробентоса Авачинской губы//Гидробиологические исследования в Авачинской губе. Владивосток: ДВО АН СССР, 1989. С. 113.
- Гонтарь В.И. Фауна мшанок отряда Cheilostomata прибрежных вод Курильских островов//ОНП НПЕТ «Верас-Эко» и АН Белоруссии. 1992. 194 c.
- Государственный природный заповедник «Магаданский» . URL: http://www.magterra.ru/(дата обращения: 25.11.2014).
- Грищенко А.В. Эвристомные мшанки (Bryozoa: Ctenostomida, Cheilostomida) шельфа Командорских островов//Донная флора и фауна шельфа Командорских островов. Владивосток: Дальнаука, 1997. С. 153-192.
- Грищенко А.В. Камчатский краб как мобильный субстрат для мшанок на шельфе Западной Камчатки//Сохранение биоразнообразия Камчатки и прилегающих морей: материалы 2 науч. конф. Петропавловск-Камчатский, 2001. С. 46-47.
- Грищенко А.В. Мшанки литорали Командорских островов//Сохранение биоразнообразия Камчатки и прилегающих морей: материалы 5 науч. конф. Петропавловск-Камчатский, 2004. С. 38-43.
- Грищенко А.В., Фостер Н.Р. Мшанки литорали бухты Датч-Харбор, остров Уналашка, Алеутские острова//Вестник Пермского университета, 2012. № 3. С. 23-29.
- Зуенко Ю.И. Приливное перемешивание как фактор биопродуктивности Дальневосточных морей в летний сезон//Океанологические основы продуктивности северо-западной части Тихого океана. Владивосток: ТИНРО, 1992. С. 56-79.
- Изюмова Е.А. Данные по экологии мшанок отряда Cheilostomata Западнокамчатского шельфа//1 Всесоюзная конференция по морской биологии: тез. докл. Владивосток, 1977. С. 59-60.
- Изюмова Е.А., Кубанин А.А. Тип Podaxonia, Класс Bryozoa. Список животных литорали Восточной Камчатки и западного побережья Берингова моря//Литораль Берингова моря и юго-восточной Камчатки. М.: Наука, 1978. С. 170-171.
- Ильичев А.В., Рязанцева И.А. Животный мир литорали острова Талан//Прибрежные экосистемы северного Охотоморья. Остров северного Охотоморья. Остров Талан. Магадан: СВНЦ ДВО РАН, 1992. С. 48-71.
- Клюге Г.А. Класс Bryozoa//Фауна Охотского моря и условия ее существования. Ленинград: Изд-во АН СССР, 1953. С. 169-179.
- Клюге Г.А. Список видов мшанок (Bryozoa) дальневосточных морей СССР//Исслед. дальневосточных морей СССР. 1961. Т. 7. С. 118-143.
- Клюге Г.А. Мшанки северных морей СССР. М.; Л.: Изд-во АН СССР, 1962. 584 с.
- Клюге Г.А., Андросова Е.И., Гостиловская М.Г. Тип Podaxonia, Класс Bryozoa. Список фауны морских вод Южного Сахалина и южных Курильских островов//Исслед. дальневосточных морей СССР. 1959. Т. 6. С. 209-213.
- Кубанин А.А. Мшанки отряда Cheilostomata залива Петра Великого Японского моря//Обрастания в Японском и Охотском морях. Владивосток: ДВНЦ АН СССР, 1975. Т. 3. С. 108-136.
- Кубанин А.А. Мшанки литорали острова Завьялова (Охотское море)//Биол. моря. 1976. № 1. C. 30-35.
- Кубанин А.А. Тип Bryozoa. Предварительный список беспозвоночных животных литорали Дальневосточного государственного морского заповедника//Исследования литорали Дальневосточного морского заповедника и сопредельных районов. Владивосток, 1987. С. 121-122.
- Кубанин А.А. Phyllum Tentaculata, subphyllum Bryozoa//Кусакин О.Г., Иванова М.Б., Цурпало А.П. Список видов животных, растений и грибов литорали дальневосточных морей России. Владивосток: Дальнаука, 1997. С. 119-125.
- Кубанин А.А. Phyllum Bryozoa//Адрианов А.В., Кусакин О.Г. Таксономический каталог биоты залива Петра Великого Японского моря. Владивосток: Дальнаука, 1998. С. 300-306.
- Кубанин А.А., Тарасова Н.А. К фауне мшанок отряда Cheilostomida сублиторали острова Монерон//Бентос шельфа острова Монерон. Владивосток: Изд-во ДВНЦ АН СССР, 1985. С. 102-113.
- Ландшафты, климат и природные ресурсы Тауйского губы Охотского моря. Владивосток: Дальнаука, 2006. 525 с.
- Регель К.В. Морские и солоноватоводные беспозвоночные Тауйской губы Охотского моря//Биологическое разнообразие Тауйской губы Охотского моря. Владивосток: Дальнаука, 2005. С 479-544.
- Ростов И.Д. и др. Атлас по океанографии Берингова, Охотского и Японского морей//Информационные ресурсы ТОИ. Океанография. 2003.
- Спиридонов В.А. Отчёт о научно-исследовательской работе по Договору №1 от 07 июня 2013 г. с Санкт-Петербургской общественной организацией "Экология и бизнес" в рамках проекта ГЭФПРООН "Укрепление морских и прибрежных ООПТ России" по теме: «Исследования литорали государственного природного заповедника Магаданский». Магадан, 2013. 78 c.
- Dick M.H., Ross J.R.P. Intertidal Bryozoa (Cheilo-stomata) of the Kodiak vicinity, Alaska. Belling-ham: Center for Pacific Northwest Studies Occasional Paper 23, Western Washington University, 1988. 133 p.
- Dick M.H., Grischenko A.V., Mawatari S.F. Intertidal Bryozoa (Cheilostomata) of Ketchikan, Alaska//J. Nat. Hist. 2005. Vol. 39(43). P. 3687-3784.
- Gontar V.I. On the Cheilostomata (Bryozoa) of the Kuril Islands//Larwood G.P., Nielsen C., editors. Recent and fossil Bryozoa. Fredensborg: Olsen & Olsen, 1981. P. 101-103.
- Gontar V.I., Denisenko N.V. Arctic Ocean Bryozoa//Herman Y., editor. The Arctic Seas. Climatology, Oceanography, Geology, and Biology. Van Nostrand Reinhold Company, N. Y., 1989. P. 341-371.
- Grischenko A. V. History of investigations and current state of knowledge of Bryozoan species diversity in the Bering Sea//Wyse Jackson P.N., Spencer Jones M.E., editors. Annals of Bryozoology: Aspects of the history of research on bryozoans. International Bryozoology Association. Dublin: Trinity College, 2002. P. 97-116.
- Grischenko A.V., Gordon D.P., Taylor P.D. A unique new genus of Cheilostomate Bryozoan with re-versed-polarity zooidal budding//Asian Mar. Biol. 1999. Vol. 15. P. 105-117.
- Grischenko A.V., Dick M.H., Mawatari S.F. Diversity and taxonomy of intertidal Bryozoa (Cheilostoma-ta) at Akkeshi Bay, Hokkaido, Japan//J. Nat. Hist. 2007. Vol. 41(17-20). P. 1047-1161.
- Kubanin A.A. Phylum Bryozoa//Kussakin O.G., Kostina E.E., editors. The intertidal biota of volcanic Yankich Island (middle Kuril Islands). Publ. Seto Mar. Biol. Lab. 1996. Vol. 37(3/6). P. 201-225.
- Kuklinski P. Ecology of stone-encrusting organisms in the Greenland Sea -a review//Polar Res. 2009. Vol. 28. P. 222-237.
- Kuklinski P., Barnes D.K.A. Bryodiversity on coastal boulders at Spitsbergen//H.I. Moyano et al., editors. Bryozoan studies 2004. London: A.A. Balkema Publishers, 2005. P. 161-172.
- Kuklinski P., Barnes D.K.A. Structure of intertidal and subtidal assemblages in Arctic vs temperate boulder shores. Polish Pol. Res. 2008. Vol. 29. P. 203-218.
- Kuklinski P., Taylor P.D. A new genus and some cryptic species of Arctic and boreal calloporid cheilostome bryozoans//J. Mar. Biol. Ass. U.K. 2006a. Vol. 86. P. 1035-1046.
- Kuklinski P., Taylor P.D. Unique life history strategy in a successful Arctic bryozoan, Harmeria scutulata. J. Mar. Biol. Ass. U.K. 2006b. Vol. 86. P. 1035-1046.
