Первые сведения о свободноживущих инфузориях планктона озера Аслы-Куль(Республика Башкортостан, Южное Приуралье)
")
Автор: Быкова С.В., Андреева В.А.
Журнал: Самарская Лука: проблемы региональной и глобальной экологии @ssc-sl
Рубрика: Научные сообщения
Статья в выпуске: 4 т.29, 2020 года.
Бесплатный доступ
В статье приводятся первые сведения о видовом составе свободноживущих инфузорий планктона, их распределении в толще воды разных экотопов (пелагиаль, открытая литораль, заросли макрофитов) памятника природы оз. Аслы-Куль. Выявлено бедное видовое богатство (по 2-12 видов в пробе) и низкое обилие инфузорий, характерное для многих солоноватоводных водоемов. Численность варьировала в диапазоне 4-789 тыс. экз./м3; биомасса - 0.2-44.8 мг/ м3. Сапробность среды по индикаторным видам инфузорий соответствовала β'-мезосапробной зоне.
Инфузории, планктон, видовое разнообразие, пространственное распределение, сезонная динамика, экологическое состояние водоема
Короткий адрес: https://sciup.org/148315300
IDR: 148315300 | УДК: 593.17:574.52(28):470.57 | DOI: 10.24411/2073-1035-2020-10352
The first data on the free-living ciliates in the plankton of lake Aslykul (Bashkortostan Republic, Southern Urals)
This article gives the first information about the species composition of free-living ciliates and its distribution in the water column from different ecotopes (pelagial, open littoral, the zone of macrophytes) of the lake Aslykul that is a natural monument. Poor species richness (2-12 species in the sample) and low ciliates abundance that is typical for brackish water bodies were revealed. The number varied in the range of 4-789 thousand copies/m3; the biomass - 0.2-44.8 mg / m3. The water saprobity values that were determined by the indicator ciliates species corresponded to the β' -mesosaprobic zone.
Текст научной статьи Первые сведения о свободноживущих инфузориях планктона озера Аслы-Куль(Республика Башкортостан, Южное Приуралье)
Key word: infusoria, plankton, species diversity, spatial distribution, seasonal dynamics, ecological state of the reservoir.
Озеро Аслы-Куль (встречается разное написание: Аслыкуль или Асликуль, башк. Асылыкүл) – самое крупное в Башкортостане: площадь водного зеркала составляет 23,5 км2. Оно является гидрологическим памятником природы регионального значения и находится в Давлекановском районе на территории одноименного природного парка. Водоем (координаты: 54°18′46″ с. ш. 54°34′38″ в. д.) расположен в бассейне р. Дема (приток р. Белой) у северо-восточной окраины Бугульминско-Белебеевской возвышенности [8]. Помимо эстетического, хозяйственного и т.д., оно имеет огромное рекреационное значение,
привлекая к себе ежегодно тысячи отдыхающих, и, кроме того, очень важно для научных исследований. Водоем является транзитным для многих перелетных птиц. Огромный научный интерес представляют собой мелкие организмы, обитающие в толще воды (планктонное сообщество в целом) данного слабосолоноватого водоема; их исследование необходимо как с точки зрения изучения биоразнообразия экосистемы озера, так и с точки зрения мониторинга процессов, происходящих в озере и определяющих его экологическое состояние. Исследование планктона любого водоема традиционно связано, в первую очередь, с изучением фитопланктона и зоопланктона как основных его составляющих. На оз. Аслы-Куль исследованием фитопланктона в разное время занимались Ф.Б. Шкундина, Г.А. Гуламанова [13], Н.Г. Тарасова [9] исследованием зоопланктона – О.В. Мухортова, Р.З.
BB%D1%8C) [15]. Вероятно, это определяет химический состав и минерализацию в целом, наличие минеральной взвеси в толще воды и, конечно же, ее своеобразный цвет. Вода в озере сульфатно-гидрокарбонатная кальциймагниевой группы с суммарной минерализацией 1,74 г/л [8]. По данным наших сотрудников (Уманская, Горбунов, настоящий сборник), минерализация составляла 1,87 г/л в 2010 г. и 1,80 г/л в 2012 г. Сумма ионов в роднике (западный берег, ст. 3 – напротив) – значительно ниже 703 мг/л. По степени минерализации вод [6], Аслы-Куль считается олиго-галинным (слабосолоноватым). Трофический индекс Карлсона (TSI=36,9) по прозрачности воды озера в 2005-2007 гг. указывал на олиготрофное состояние водоема [12]. Однако усиление темпов антропогенного эвтрофиро-вания, связанное с интенсификацией сельского хозяйства, а также с развитием рекреационного использования природных парков является основным направлением изменения состояния озера [13]. Так, в 2010 г., по данным авторов О.В. Мухортовой и Р.З. Сабитовой [7], оз. Аслы-Куль по «шкале трофности», построенной с использованием биомассы зоопланктона относилось к переходному от олиготрофного к мезотрофному типу. При этом «в зарослях макрофитов, показатели биомассы зоопланктона достигали значений, свойственных эвтрофным водам». Еще одна проблема, затронувшая озеро в последнее время – сильное обмеление. И объясняют его не только естественной цикличностью изменения уровня, но и негативным антропогенным воздействием.
Во время исследования во всей толще воды практически всегда регистрировали гомотермию, за исключением июня 2010 г., когда в условиях безветренной погоды и сильной инсоляции наблюдался значительный прогрев поверхностных слоев с формированием подповерхностного термоклина: перепад температур от поверхности до глубины 1 м составил 1,70С. В целом, разность температур между поверхностными и придонными горизонтами только в июне 2010 г. составила З.бОС, в то время как в другие даты отбора -не превышала 0.70С. Средние в столбе воды величины температур в разные даты отбора составили: 21.3ОС и 19.40С в июне и августе 2010 г.; 16.9ОС, 27.50С, 14.9 0 с — в мае, июле, сентябре 2012 г., соответственно. Кислородный режим был благоприятным для гидробионтов во всей толще воды. Прозрачность варьировала в пределах 1,3-3,6 м. Относительно невысокая, в сравнении с аналогичны-24
Сабитова [7]. Для исследования гетеротрофного компонента планктона пристального внимания заслуживает также изучение простейших, в частности инфузорий. В данной работе приводятся результаты первого исследования свободноживущих инфузорий планктона в разных биотопах водоема.
Цель работы – оценить видовое разнообразие и количественное развитие свободножи-вущих инфузорий планктона в солоноватых условиях оз. Аслы-Куль.
МАТЕРИАЛЫ И МЕТОДЫ
Исследование свободноживущих инфузорий проводили в рамках комплексных экспедиций сотрудников лаборатории экологии простейших и микроорганизмов, исследовавших фито-, зоо-, бактериопланктон и бактериобентос в июне, августе 2010 г. и в мае, июле и сентябре 2012 г. [9, 11]. Пробы отбирали батометром Рутнера в пелагической части озера (ст. 1), литорали (ст. 2 и ст. 3) и зарослях прибрежно-водных (тростники) и погруженных (рдесты) макрофитов (рис. 1). Литоральная станция 3 располагалась напротив места впадения родника. На пелагической станции для учета вертикального распределения показателей развития инфузорий отбор проб проводили через каждый метр до дна, в открытой литорали и зарослях макрофитов – только с поверхностных горизонтов. Сравнение сообществ из разных экотопов проводили по пробам из поверхностных горизонтов. Для идентификации видового состава и количественного учета инфузорий использовали известные протозоологические и гистохимические методы. Для оценки видового разнообразия использовали общеизвестные индексы (Шеннона, Пиелу, Симпсона).
РАЙОН ИССЛЕДОВАНИЯ
Происхождение озера – палеоэрозионное [1, цит. по: Г.В.Вахруше [4]]. Питание смешанное, снеговое, подземное и дождевое. В озеро впадает ручей Шарлама и многочисленные родники. Морфометрические характеристики водоема: длина – 7,1 км, средняя ширина – 3,3 км, средняя глубина – 5,1 м (максимальная – 8,1 м), объем – 119 млн. м3, длина береговой линии – ~ 20 км (Реестр особо охраняемых…, 2010). В строении водосбора принимают участие верхнепермские отложения с известняками, глинами, а также гипсово-ангидритовая соленосная и ангидритоводоломитовая толщи Кунгурского яруса %D0%BB%D0%B8%D0%BA%D1%83%D0% ми солоноватоводными водоемами, прозрачность, обусловлена, вероятно, значительным количеством минеральной взвеси, присутствующей в толще воды. По показателям активной реакции среды (pH = 8.2-8.9), водоем относится к олигощелочному типу. Активная реакция среды воды из родника на западном берегу озера близка к нейтральным значениям (pH = 7,0-7.6).
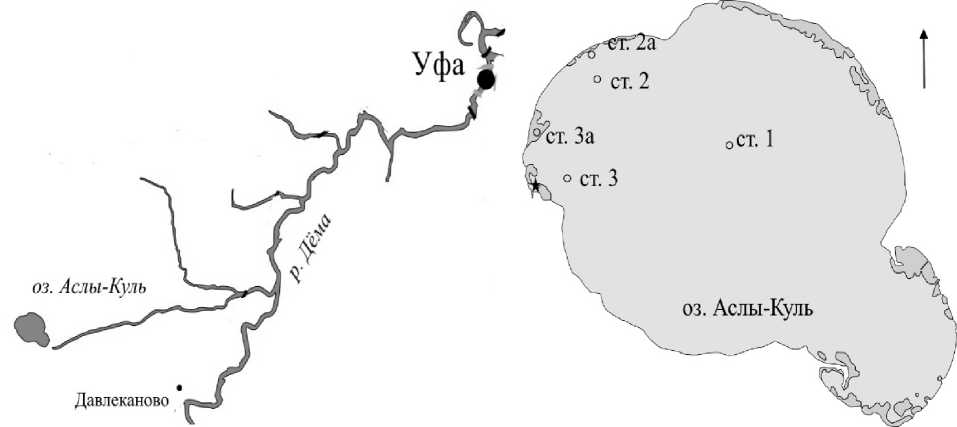
Рис. 1. Схема расположения озера и станций отбора проб по его акватории: ст. 1 – пелагиаль, ст. 2 и с т. 3 – открытая литораль, ст. 2а – заросли тростника обыкновенного (Phragmites australis (Cav.) и ст. 3а – заросли рогоза узколистного (Typha angustifolia L.). Более темной заливкой показаны заросли макрофитов; звездочкой обозначен родник
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Неоднородность сообщества инфузорий по акватории озера. Фауна инфузорий оз. Аслы-Куль довольно бедна. За период исследования в планктоне озера было зарегистрировано всего 34 вида инфузорий: 27 видов выявлено в пелагической части (в поверхностном слое пелагиали – 15 видов), 16 видов в литорали и 20 видов в зарослях макрофитов (табл. 1). В отдельных пробах регистрировалось не более 12 видов. В составе сообщества инфузорий около 40% видов встретилось лишь в 1 пробе (в основном, в зарослевых экотопах). И всего лишь 11% (4 вида: Pelagostrombidium mirabile (Penard, 1916), Rimostrombidium hyalinum (Mirabdulaev, 1985), Tintinnopsis cylindrata Kof. & Cam., 1892, Codonella cratera (Leidy, 1887)) встречались в каждой второй пробе (частота встречаемости более 50%). Вероятно, это свидетельствует о не полно выявленном видовом составе, но возможно, и о гетерогенности среды обитания и неоднородности распределения инфузорий по акватории. Наиболее сходными по фауне инфузорий были сообщества пелагической части и открытой литорали (коэффициент сходства –
64,5%, наименее – пелагиали и фитали (К=51.4%).
Прибрежные экотопы, особенно заросле-вые, богаче видами. Видовое разнообразие в целом тоже выше в прибрежной части. Однако вследствие обнаружения крупных видов в зарослевых экотопах, по биомассе структура сообщества инфузорий в них, в отличие от открытой литорали, менее выровнена, что сказалось и на индексе Шеннона (табл. 1). По численности же видовое разнообразие (индекс Шеннона и индекс Пиелу) закономерно увеличиваются в ряду «пелагиаль-открытая литораль – фиталь». Что же касается количественного развития, то и численность, и биомасса инфузорий в прибрежных экотопах ниже, чем в глубоководной части озера. Вероятно, в зарослевых экотопах функцию потребителей органического вещества в большей мере берут на себя перифитонные инфузории. В подтверждении тому, нами было отмечено развитие крупных колоний (до 500 и более зооидов в колонии) Epistylis plicatilis Ehrb., 1831 на тростнике. Кроме того, в планктонных пробах зарослевых экотопов массово встречались отдельные зооиды данного вида.
Таблица 1
Видовой состав и характеристика сообществ инфузорий в разных экотопах оз. Аслы-Куль в 2010 и 2012 гг.
|
Показатель |
Численность, % |
Биомасса, % |
|||||||
|
Экотоп |
43 В cd в |
43 ч Он о |
В н В о и св 5 |
43 В cd в |
43 Ч cd о ч |
н в •е о св 5 |
Он 0х ю Р cd о н о |
||
|
Горизонт |
0 м-дно 0 м |
0 |
м 0 |
м |
0 м-дно |
0 м |
0 м 0 |
м |
|
|
Amphileptus pleurosigma (Stokes, 1884) |
0.0 |
0.0 |
0.0 |
1.8 |
0.0 |
0.0 |
0.0 |
2.8 |
2.6 |
|
Askenasia acrostomia Krainer & Foissner, 1990 |
0.2 |
0.2 |
0.0 |
0.4 |
0.1 |
0.1 |
0.0 |
0.0 |
7.9 |
|
Askenasia sp. |
0.5 |
0.4 |
1.1 |
2.3 |
0.5 |
0.4 |
1.1 |
0.8 |
26.3 |
|
A. chlorelligera Krainer & Foissner, 1990 |
0.3 |
0.0 |
0.6 |
0.4 |
0.1 |
0.0 |
0.2 |
0.04 |
10.5 |
|
A. volvox (Eichwald, 1852) Kahl, 1930 |
1.8 |
1.9 |
1.7 |
0.6 |
0.3 |
0.3 |
0.3 |
0.04 |
21.1 |
|
Balanion planctonicum Foissner et al., 1994 |
0.04 |
0.0 |
0.0 |
0.0 |
0.004 |
0.0 |
0.0 |
0.0 |
2.6 |
|
Cinetochilum margaritaceum Perty, 1852 |
0.0 |
0.0 |
0.0 |
0.4 |
0.0 |
0.0 |
0.0 |
0.01 |
2.6 |
|
Codonella cratera (Leidy, 1887) |
8.4 |
3.1 |
16.2 |
13.0 |
5.4 |
2.0 |
10.5 |
3.0 |
50.0 |
|
Coleps cf. elongatus Ehrenberg, 1830 |
0.04 |
0.0 |
0.0 |
0.0 |
0.01 |
0.0 |
0.0 |
0.0 |
2.6 |
|
Coleps hirtus (Müller, 1786) Nitzsch, 1827 |
0.0 |
0.0 |
0.6 |
0.0 |
0.0 |
0.0 |
0.2 |
0.0 |
2.6 |
|
Cyclidium glaucoma Müller, 1773 |
0.1 |
0.0 |
0.0 |
1.5 |
0.0 |
0.0 |
0.0 |
0.03 |
7.9 |
|
Epicarchesium pectinatum (Zacharias 1897) |
5.0 |
4.5 |
10.1 |
3.1 |
52.4 |
51.2 |
45.6 |
2.8 |
31.6 |
|
Epistylis plicatilis Ehrb., 1831 |
0.0 |
0.0 |
0.0 |
0.5 |
0.0 |
0.0 |
0.0 |
72.3 |
2.6 |
|
Frontonia cf. acuminata (Ehrb., 1834) |
0.04 |
0.0 |
0.0 |
0.0 |
0.1 |
0.0 |
0.0 |
0.0 |
2.6 |
|
Halteria grandinella (O.F. Muller, 1773) |
1.2 |
0.1 |
8.4 |
0.0 |
0.2 |
0.0 |
1.2 |
0.0 |
28.9 |
|
Holophrya spp. |
0.4 |
0.0 |
0.6 |
0.2 |
0.5 |
0.0 |
0.7 |
0.0 |
13.2 |
|
Hypotricha sp. 1 |
0.0 |
0.0 |
0.0 |
0.4 |
0.0 |
0.0 |
0.0 |
0.0 |
2.6 |
|
Hypotricha sp. 2 |
0.0 |
0.0 |
0.0 |
0.2 |
0.0 |
0.0 |
0.0 |
0.1 |
2.6 |
|
Lagynophrya acuminata Kahl, 1935 |
0.1 |
0.2 |
0.0 |
1.0 |
0.1 |
0.2 |
0.0 |
0.3 |
7.9 |
|
Limnostrombidium pelagicum (Kahl, 1932) |
0.1 |
0.1 |
0.0 |
0.0 |
0.02 |
0.04 |
0.0 |
0.0 |
2.6 |
Окончание таблицы 1
|
L. viride (Stein, 1867) Krainer, 1995 |
0.2 |
0.2 |
0.0 |
0.0 |
0.2 |
0.3 |
0.0 |
0.0 |
5.3 |
|
Mesodinium sp. |
0.1 |
0.0 |
0.0 |
0.0 |
0.004 |
0.0 |
0.0 |
0.0 |
2.6 |
|
Paramecium aurelia complex |
0.04 |
0.0 |
0.6 |
0.0 |
0.1 |
0.0 |
0.9 |
0.0 |
5.3 |
|
Pelagohalteria viridis (Fromentel, 1876) |
5.5 |
5.9 |
2.2 |
4.5 |
1.8 |
2.0 |
0.7 |
0.5 |
47.4 |
|
Pelagostrombidium mirabile (Penard, 1916) |
16.4 |
18.6 |
44.5 |
15.6 |
12.2 |
14.1 |
33.7 |
4.2 |
76.3 |
|
Pseudovorticella fasciculata (Müller 1773) |
0.1 |
0.0 |
1.7 |
0.0 |
0.03 |
0.0 |
0.7 |
0.0 |
5.3 |
|
Pseudovorticella sp. |
0.1 |
0.0 |
0.0 |
0.0 |
0.1 |
0.0 |
0.0 |
0.0 |
2.6 |
|
Rimostrombidium hyalinum (Mirabdulaev, 1985) |
6.5 |
2.6 |
7.6 |
7.8 |
1.4 |
0.6 |
1.7 |
0.6 |
52.6 |
|
Strobilidium caudatum (Fromentel, 1876) |
0.3 |
0.0 |
0.0 |
31.7 |
0.2 |
0.0 |
0.0 |
9.1 |
18.4 |
|
Strongylidium lanceolatum Kowal., 1882 |
0.0 |
0.0 |
0.6 |
0.0 |
0.0 |
0.0 |
0.6 |
0.0 |
2.6 |
|
Thuricola sp. |
0.04 |
0.0 |
0.0 |
0.0 |
0.1 |
0.0 |
0.0 |
0.0 |
2.6 |
|
Tintinnopsis cylindrata Kof. & Cam., 1892 |
51.6 |
61.1 |
3.1 |
1.6 |
23.5 |
28.2 |
1.4 |
0.3 |
50.0 |
|
Urotricha spp. |
0.1 |
0.1 |
0.0 |
0.0 |
0.02 |
0.04 |
0.0 |
0.0 |
2.6 |
|
Vorticella spp. |
0.9 |
0.9 |
0.6 |
13.2 |
0.6 |
0.7 |
0.4 |
3.0 |
39.5 |
|
Суммарный показатель (N, тыс. экз./м3; B, мг/ м3) |
127.6 |
307.9 |
51.0 |
61.8 |
4.2 |
10.0 |
1.7 |
5.7 |
- |
|
Число видов, экз. |
27 |
15 |
16 |
20 |
27 |
15 |
16 |
20 |
- |
|
Индекс Шеннона, бит/ экз. |
2.40 |
1.92 |
2.66 |
3.07 |
2.05 |
1.84 |
2.08 |
1.65 |
- |
|
Индекс Пиелу |
0.51 |
0.49 |
0.67 |
0.71 |
0.43 |
0.47 |
0.52 |
0.38 |
- |
Видовая структура. Несмотря на относительно значительное сходство сообществ инфузорий разных экотопов (коэффициент сходства Съеренсена выше 50%), доминирующие виды, занимающие первую позицию в видовой структуре во всех трех экотопах, были представлены разными видами (табл. 1): в пелагической части по численности лидировал T. cylindrata (61%), в литоральной – Pela-gostrombidium mirabile (45%), а в поясе макрофитов – Strobilidium caudatum (Fromentel, 1876) (32%).
По биомассе везде первую позицию занимали колониальные виды: Epicarchesium pectinatum (Zacharias 1897) – в пелагиали и открытой литорали (51и 46% общей биомассы, соответственно) и Epistylis plicatilis – в заросле-вой части водоема (72% биомассы). Обращает на себя внимание четкое перераспределение доминирования разных видов тинтиннид (отр. Tintinnida) на разных участ-к d н в ш C е д 1
Сезонное изменение видового богатства, численности и иомассы инфузори. ри относительно бедном видовом составе, максимум видового богатства (20 видов) был зарегистрирован в июле, в то время как в мае встречалось всего лишь 5 видов, а в сентябре – 10 видов. Сезонная сукцессия инфузорий выражена не так явно, как для фитопланктона, тем не менее происходила смена доминирования некоторых видов от весны к осени. Численность Pelagostrombidium mirabile снижалась к осени, несмотря на то, что вид доминировал в планктоне озера в целом весь исследуемый период. Весной, помимо данного вида, домиинировали T. cylindrata и E. pectinatum , летом – C. cratera и мелкие Halter-ia grandinella и R. hyalinum , осенью – снова колониальный E. pectinatum и C. cratera . Разные виды тинтиннид развивались в противофазе и демонстрировали расхождение не только по экотопам, но и по времени: вклад в численность T. cylindrata в среднем по акватории озера максимален весной (23%), а C. cratera – осенью (32%).
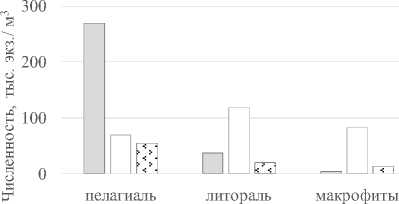
Сезонная динамика численности и биомассы инфузорий в 2012 г. в разных экотопах акватории озера значительно отличалась. В пе- лагической части водоема максимум развития инфузорий приходился на весенний период, в прибрежной части – максимум численности и биомассы (за небольшим исключением) – на летний (рис. 2).
В целом, от весны к осени численность инфузорий в среднем по акватории уменьшается от 103 до 30 тыс. экз./м3, а биомасса от 8.9 до 1.1 мг/ м3. В более жарком 2010 г. значения численности и биомассы, в среднем были ненамного выше – 122 тыс. экз./м3 численности и 4,3 мг/ м3 биомассы, чем в 2012 г. (в среднем 70 тыс. экз./м3 численности и 3,7 мг/м3 биомассы, соответственно). В 2010 г. численность варьировала в пределах 8-789 тыс. экз./м3, биомасса – 0,2-44,8 мг/ м3, а в 2012 г. – 4-462 тыс. экз./м3 и 0,05-33,3 мг/ м3, соответственно. Уровень количественного развития инфузорий в оз. Аслы-Куль можно считать довольно низким и соответствующим ультраолиготрофному статусу водоема [14].
s св
S о s
^

пелагиаль литораль макрофиты экотоп май июль сентябрь
май июль сентябрь
экотоп
Рис. 2. Сезонное изменение численности и биомассы инфузорий в оз. Аслы-Куль в 2012 г.
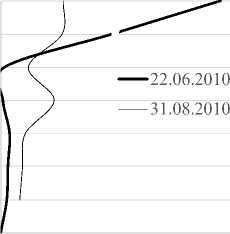
Вертикальное распределение инфузорий в толще воды °C. В июне в условиях большего прогрева поверхностных слоев воды (рис. 3) значительная часть видов была сосредоточена у поверхности (11 видов). Второй пик формировался на горизонте 4-5 м (6-7 видов). В августе, в условиях более равномерно прогретой толщи воды, профиль видового богатства более выровнен с незначительными пиками на глубине 1 м и у дна. Изменение численности и биомассы с глубиной происходит синхронно, с преобладанием значений данных показа-
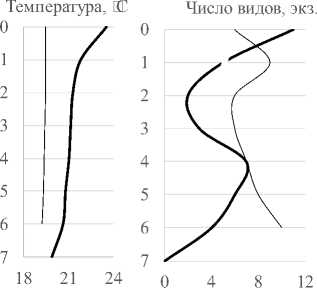
Рис. 3 Вертикальные профили температуры, видового богатства, численности и биомассы сообщества инфузорий в 2010 г. ( разрыв линии на 1м - отсутствие данных)
Численность, тыс. экз./м3
0 300 600 900
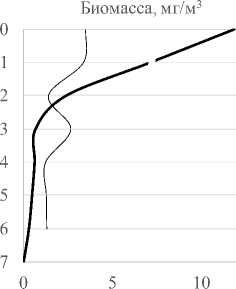
зателей в поверхностном слое воды в июне. Из-за отсутствия температурной и химической стратификации распределение видов не имеет закономерностей, характерных для распределения видов в стратифицированных водоемах с дефицитом кислорода в нижних слоях. Тем не менее, роль отдельных видов меняется с глубиной. В июне 2010 г. T. cylindrata формируют максимальные вклады в общую численность на поверхности , E. pectinatum – на глубине 2 м, Pelagohalteria viridis (Fromentel, 1876) – на глубине 4 м, инфузории р. Vorticella – на глубине 5 м, A. volvox (Eichwald, 1852) Kahl, 1930 на глубине 6 м и т.д.
В августе, в отсутствие колониальных видов, у поверхности сосредоточиваются тин-тинниды , в средних слоях к ним присоединяется R. hyalinum .
Сапробность среды, определенная по инфузориям-индикаторам сапробности, в целом за исследованный период, соответствует верхней границе β'-мезосапробной зоны (S = 1.99). В более жарком 2010 г. индикаторные виды инфузорий указывали на немногим более высокий уровень органического загрязне- ния (S = 2.03, β"-мезосапробная зона), чем в 2012 г. (S = 1.91, β'-мезосапробная зона) (табл. 2). На станции около родника сапробность минимальна (S = 1.61). В оба года в глубоководной части, по сравнению с прибрежными участками, весной и летом сапробность выше. Сезонное изменение уровня органического загрязнения по инфузориям-индикаторам в глубоководной и прибрежной частях происходит в противофазе: в пелагиали сапроб-ность среды от весны к осени снижается, а в открытой литорали и, особенно, в литорали, заросшей тростниками, напротив, увеличивается (табл. 2). Вероятно, весной при отсутствии или слабом развитии макрофитов, выполняющих барьерную функцию, все органическое вещество, попадающее с весенним половодьем с водосборной территории попадает в толщу воды и распределяется по всей акватории. Осенью же, пояс макрофитов «защищает» пелагиаль, «усваивая» часть органического вещества, при этом сами высшие растения, отмирая, являются источником вторичного загрязнения. Поэтому индекс сапробно-сти в фитали, повышаясь к осени, превышает индекс сапробности в пелагиали (табл. 2).
Таблица 2
Сапробность воды (по инфузориям-индикаторам) оз. Аслы-Куль
|
Экотоп Месяц, год |
пелагиаль |
литораль |
макрофиты |
Вся акватория |
|
Июнь 2010 г. |
2.14 |
- |
1.54 |
2.01 |
|
Август 2010 г. |
2.12 |
1.6 |
1.74 |
2.07 |
2010 г. в целом 2.03
|
Май 2012 г. |
1.98 |
1.74 |
1.75 |
1.94 |
|
Июль 2012 г. |
1.99 |
1.97 |
1.77 |
1.92 |
|
Сентябрь 2012 г. |
1.68 |
2.06 |
1.87 |
1.78 |
2012 г. в целом 1.91
ЗАКЛЮЧЕНИЕ
Исследования инфузорий солоноватоводного водоема оз. Аслы-Куль показали, что для водоема такой площади видовой состав сво-бодноживущих инфузорий довольно беден (34 вида), а их обилие – низко. Вероятно, обусловлено это повышенной минерализацией.
Полагаем, что именно различием в минерализации вод двух рядом расположенных крупнейших озер данного региона – оз. Аслы-Куль (минерализация вдвое выше) и оз. Кандры-Куль (природный парк «Кандры-Куль»), обусловлены значительные различия в характеристиках их сообществ инфузорий. Число видов инфузорий в планктоне оз. Аслы-Куль почти в 4 раза, численность – в 7 раз (2010 г.) и в 26 раз (2012 г.), а биомасса – в 2 (2010 г.) и 4 (2012 г.) раза ниже, чем в планктоне оз. Кандры-Куль [3]. Как утверждают С.В. Быкова и В.В. Жариков и другие авторы [2, 5], полного аналога исследованному водоему среди изученных нами солоноватоводных водоемов Среднего Поволжья нет, но по видовому богатству и уровню количественного развития сообщества инфузорий, оз. Аслы-Куль близко к водоемам Вятского поднятия, схожих по минерализации [10]: например, оз. Большое Голубое и оз. Голубая Старица. Оценивая экологическое состояние оз. Аслы-Куль по индикаторным видам и количественным характеристикам инфузорного сообщества, можно констатировать, что трофический статус водоема близок к ультраолиготрофно-му уровню, а сапробность среды относится, в основном, к Р' -мезосапробной зоне.
Список литературы Первые сведения о свободноживущих инфузориях планктона озера Аслы-Куль(Республика Башкортостан, Южное Приуралье)
- Абдрахманов Р.Ф. Пресные подземные и минеральные лечебные воды Башкортостана // Уфа: Изд-во «Гилем» НИК «Башкирская энциклопедия», 2014. С. 416.
- Быкова С.В., Жариков В.В. Сравнительный анализ сообществ инфузорий (СШор^га) высокоминерализованных водоемов двух тектонических поднятий Поволжья // Поволжский экологический журнал. 2012. № 2. С. 142-156.
- Быкова С.В., Жариков В.В., Андреева В.А., Горбунов М.Ю., Уманская М.В. Инфузории озера Кандры-Куль (респ. Башкортостан): состав, пространственное распределение, сезонная динамика и экологическое состояние водоема по данным их сообщества в 2012 г. // Известия Самарского научного центра РАН. 2014. Т. 16, № 5(5) С. 1748-1757.
- Вахрушев Г.В. Минеральные воды и грязи Башкирии. Уфа: Башкнигоиздат, 1961. 156 с. -Цит. по: Абдрахманов, 2014.
- Жариков В.В., Горбунов М.Ю., Уманская М.В., Тарасова Н.Г., Быкова С.В., Шерышева Н.Г., Мухортова О.В., Сабитова Р.З., Краснова Е.С. Современное состояние экосистемы озера Кандры-Куль. Тольятти: Анна, 2018. 229 с.
- Китаев С.П. Основы лимнологии для гидробиологов и ихтиологов. Петрозаводск: Карельский научный центр РАН, 2007. 395 с.
- Мухортова О.В., Сабитова Р.З. Зоопланктон озера Асликуль (Республика Башкортостан) // Известия Самарского научного центра РАН. 2014. Т. 16, № 5. С. 257-263.
- Реестр особо охраняемых природных территорий Республики Башкортостан. Изд. 2-е, пере-раб. Уфа: Изд. центр «МедиаПринт», 2010. 414 с.
- Тарасова Н.Г. Фитопланктон озера Асликуль (республика Башкортостан) // Материалы IX Междунар. науч.-практ. конф. «Татищевские чтения: актуальные проблемы науки и практики» / Актуальные проблемы экологии и охраны окружающей среды. Тольятти: Волжский университет им. В.Н. Татищева, 2012. С. 202-210.
- Уникальные экосистемы солоноватовод-ных карстовых озер Среднего Поволжья / под ред. А.Ф. Алимова, Н.М. Мингазовой. Казань: Изд-во Казан. гос. ун-та, 2001. 256 с.
- Шерышева Н.Г., Ракитина Т.А., Повет-кина Л.П. Бактериобентос оз. Асликуль в 2010, 2012 гг. (Республика Башкортостан) // Известия Самарского научного центра РАН. 2014. Т. 16, № 5(5). C. 1727-1733.
- Шкундина Ф.Б., Гуламанова Г.А. Биологическое разнообразие автотрофного планктона озер Республики Башкортостан (Россия) // Альгология. 2011. Т. 21, № 3. С. 329-345.
- Шкундина Ф.Б., Гуламанова Г.А. Основные направления изменения экосистем озер на территории Республики Башкортостан // Геология, полезные ископаемые и проблемы геоэкологии Башкортостана, Урала и сопредельных территорий. Материалы VIII Межрегион. науч.-практ. конф. Уфа: ДизайнПолиграфСервис, 2010. С. 252-254.
- Beaver J.R., Crisman T.L. The role of ciliated protozoa in pelagic freshwater ecosystems // Microbial Ecology. 1989. V. 17, No. 2. P. 111-136.
- URL:https://ru.wikipedia.org/wiki (дата обращения: 20.07.2020).
