Photosynthetic adaptations and metabolic responses of medicinal plants to elevated CO₂ and abiotic stress

Author: Patni Babita
Journal: Журнал стресс-физиологии и биохимии @jspb
Article in issue: 3 т.21, 2025.
Free access
Photosynthesis is a fundamental process influenced by plant species and environmental factors, particularly elevated atmospheric CO₂ and temperature. Increased CO₂ enhances photosynthetic rates while reducing stomatal conductance, promoting plant growth. However, abiotic stresses such as heat and drought significantly impair photosynthesis, as observed in medicinal plants. Canopy temperature regulation and transpiration rates shift under high temperatures, affecting water-use efficiency and physiological adaptations. Membrane stability, critical under stress conditions, is compromised by lipid peroxidation and oxidative damage, leading to cellular dysfunction. Antioxidant defense mechanisms, including enzymatic responses, mitigate reactive oxygen species (ROS) accumulation under abiotic stress. Elevated CO₂ enhances photosynthesis and growth in several medicinal plants, such as Ocimum sanctum and Gynostemma pentaphyllum , though stressors like drought and heat influence metabolic pathways, secondary metabolite accumulation, and overall yield. Notably, medicinal plant quality is altered by climate variables, affecting chemical composition and therapeutic efficacy. While some species benefit from climate-induced changes in metabolite production, others exhibit declines in potency. The findings highlight the complex interplay between photosynthesis, stress adaptation, and secondary metabolite biosynthesis under changing environmental conditions, emphasizing the need for further research on climate-resilient medicinal plant cultivation.
Climate change, photosynthesis, abiotic stress, medicinal plants, reactive oxygen species (ros), secondary metabolites, metabolic adaptation
Short address: https://sciup.org/143184725
IDR: 143184725
Text of the scientific article Photosynthetic adaptations and metabolic responses of medicinal plants to elevated CO₂ and abiotic stress
PHOTOSYNTHESIS
Plant species and environmental variables influence the magnitude of photosynthetic CO 2 uptake. Increased atmospheric CO 2 at high temperatures promotes plant development by increasing photosynthetic rates. Increased atm. CO 2 causes a decrease in stomatal conductance. One of the most significant effects of elevated CO 2 is an increase in the number of proteins linked to PSI and PSII. It's frequently linked to a boost in photosynthesis (Kumar et al. , 2020). The results of the study were important for Picrorhiza kurroa adaptability to elevated CO 2 and increasing temperature. To begin most proteins are involved in light reactions abundantly if exposed to elevated CO 2 level (Kumar et al. , 2020).
The photosynthetic rate of both Hedychium spicatum and Valeriana jatamansi plants decreased after 5 days modestly (2 to 9%) due to individual and synergistically introduced heat and drought stress. It declined much more under synergistic stress effects (89 to 95%) than the percentage (51 to 84 %) of drought or heat (64 to 77 %) stress 30 days after exposure (Pandey et al. , 2021). Photosynthesis is reduced in Panax quinquefolius under high heat conditions (Jochum et al. 2007). In Podophyllum polyphylla , 40 to 60 % shade was found to be ideal for photosynthesis in abiotic stress trials (Zheng et al. , 2014). A soil moisture decrease lowers chlorophyll content, photosynthetic rate, and light saturation point while increasing dark respiration rate and light compensation point. (Liang et al. , 2014). Differential expression of proteins involved in photosynthesis transcription, metabolism, protein synthesis, defense response, signalling, transport, and cytoskeleton development in Picrorhizakurrooa , as well as reduced photosynthetic rate, stomatal conductance, transpiration and Relative water content (Sanjeeta et al. , 2014)
The optimum temperature for photosynthesis was discovered in alpine and temperate populations of Podophyllum hexandrum grown in environmentally controlled rooms with variable PAR levels and temperature. Nonetheless, increased transpiration rate and reduced WUE were ascribed to the decrease in photosynthesis at higher temperatures, demonstrating that the species is susceptible to high temperatures (Kushwaha et al. 2008). Drought stress showed impaired photosynthesis in Vitex trifolia by controlling chlorophyll fluorescence and PS II production (Dian-xia 2010).
TEMPERATURE OF THE CANOPY ANDTRANSPIRATION
Under high-temperature conditions, plant metabolism may change. With rising temperatures, maintenance of respiration costs commonly rise (Sigurdsson, 2001; Lee et al. , 2005), and plants have greater daily whole-plant respiration rates (photosynthesis ratios under elevated temperature). Plants that survive in low-light situations may become more reliant on nonstructural carbohydrate reserves (NSC) when the temperature rises (Atkin et al. 2007; Ciais et al. , 2005)
Chaturvedi et al. , (2009) studied the effect on Podophyllum hexandrum exposed to control and increased (650 µmol·mol-1) CO 2 concentrations for four months by doubling the CO 2 concentration in OTC. They discovered that the photosynthetic rate increased while transpiration and stomatal conductance decreased dramatically.
Hao et al. (2013) used FACE technology to study the response of an important Chinese medicinal plant, Istatisindigotica, to an enhanced CO 2 level of 550 µmol·mol-1. When compared to control, photosynthetic rate and water usage efficiency increased significantly while transpiration rate and stomatal conductance decreased.
Chaturvedi et al. , (2009) studied the effect of doubling CO 2 content in OTC with an endangered medicinal herb ( Podophyllum hexandrum ) for four months under control and increased (650 µmol·mol-1) CO 2 conditions. They discovered that the photosynthetic rate increased while transpiration and stomatal conductance decreased fastly.
Using an open-top chambers (OTC) technique Singh et al. (2016) investigated the impact of increased CO2 concentration (800 µmol·mol-1) on growth dynamics structure, and function of the Ocimum sanctum (Holy Tulsi) plant. Photosynthesis, intercellular CO2 concentration, carboxylation efficiency, and mesophyll efficiency are stimulated by the increased CO2 concentration The water use efficiency is decreased than control, in terms of transpiration and stomatal conductance Chang et al. (2016) investigated the physiology of Gynostemma pentaphyllum at two CO2 levels: control (360 µmol·mol-1) and enhanced (720 µmol·mol-1). Increased carbon dioxide levels resulted in an increase in the photosynthetic rate, transpiration rate and stomatal conductance of plants, according to the findings. Drought stress also caused differential expression of proteins involved in metabolism photosynthesis, transcription, protein synthesis, defense response, transport, signalling, and cytoskeleton development in Picrorhizakurrooa, as well as reduced photosynthetic rate, stomatal conductance, transpiration and RWC (Sanjeeta et al. 2014)
STABILITY OF MEMBRANES
Low temperature-induced membrane lipid phase transitions have been linked to membrane integrity degradation and physiological impairment. The revelation that chilling stress elicits a complex membrane retailoring response that leads to enhanced fluidity at lower temperatures further implicates membranes and changes in their physical features as having a function in cold stress. Membrane integrity may potentially play a role in the development of irreparable harm during low-temperature stress. It would have a comparable effect to the senescent processes of free radical tissue damage and increasing membrane rigidification (Parkin et al. , 1989).
Malondialdehyde is one of the ultimate disintegration products in biomembranes after lipid peroxidation. It has been regarded as an indication of membrane damage extent (Xue and Liu, 2008), Similarly, water stress increased lipid peroxidation in the leaves of beans Morus alba , and Triticum aestivum , as well as the in the Cucumis sativus leaves and roots (Arasimowicz-Jelonek et al. , 2009). The expression of stress-responsive genes/proteins, particularly heat-induced chaperones and antioxidant proteins, is triggered by an increase in membrane fluidity during signal transduction. Temperature stress, for example, strongly stimulated around 5% (1500 genes) of plant transcriptomes (Rizhsky et al. , 2004; Qin et al. , 2008; Finka et al. , 2011)
The buildup of cellular osmolytes (e.g. polyamines proline, soluble sugars, and glycine betaine) is a biochemical marker for salt stress tolerance, partly due to the function they play in preserving the stability of membranes and other cellular structures.
Oxidative stress is caused by abiotic stressors, and there is an excess of ROS, RNS in many of the plant cell's cellular compartments (Pinheiro et al. , 2004). Plants that can withstand the stress-induced generation of reactive oxygen species (ROS) have a higher toxicity potential for biomolecules and membranes (Scandalios 2005; Miller et al. , 2008).
Water stress has also been shown to increase lipid peroxidation, membrane damage index, H 2 O 2 , and OH-generation in stressed Phaleolus vulgaris leaves (Zlatev et al. , 2006).
PHOTORESPIRATION AND RESPIRATION
Under high-temperature conditions, plant metabolism may be disrupted. With rising temperatures maintenance of respiration costs commonly rise (Sigurdsson, 2001; Atkin and Tjoelker, 2003; Lee et al. 2005), and plants have greater daily whole-plant respiration rates (photosynthesis ratios under elevated temperature). Plants that survive in low-light situations may become more reliant on non-structural carbohydrate reserves (NSC) when the temperature rises (Atkin et al. , 2007; Ciais et al. , 2005). High temperatures affect photosynthesis by affecting the structure of thylakoids, altering the excitation energy distribution, and influencing the activity of the Calvin cycle and other metabolic processes such as photoinhibition, photorespiration, and product synthesis (Berry and Bjorkman, 1980). Numerous studies have found that short-term exposure to increased CO 2 increases photosynthesis by minimizing energy waste in photorespiration (Ainsworth and Long, 2005). According to a water stress study, a decrease in soil moisture lowers chlorophyll content, photosynthetic rate, and light saturation point, while increasing light compensation point and dark respiration rate (Liang et al. 2014).
STATUS OF REDOX
Superoxide dismutase (SOD), peroxidase (POD) catalase (CAT), and ascorbate peroxidase (APX) are antioxidative enzymes that plants can activate to counteract stress reactions. As a result, antioxidant concentration in plants is a conventional criterion for evaluating plant therapeutic potential. Stress stimulates the generation of reactive oxygen species (ROS), and antioxidant enzyme activity rises to counteract these hazardous chemicals. As a result, we believe that a high REE content in D. aphyllum may produce minor salinity stress (Liu, et al., 2021).
The inactivation or denaturation of enzymes may cause inhibition of enzyme activities under high heat conditions. CAT and POD activities of Dendrobium moniliforme , a widely grown medicinal crop, were dramatically elevated during the early stages of drought stress to protect the plants against ROS damage, similar to these findings (Wu et al. 2016).
YIELD
Stutt et al., (2008) tested Scutellaria barbata and Scutellaria lateriflora with CO2 concentrations of 400 1200, and 3000 µmol·mol-1, and found an increase in growth rate and total biomass in both species in response to elevated CO2 (1200 µmol·mol-1) when compared to 400 and 3000 µmol·mol-1. Saravanan and Karti (2014) conducted a similar experiment on Catharanthus roseus, an important medicinal plant (sadabahar). Plants were exposed to control (380 µmol·mol-1) and elevated CO2 (600 and 900 µmol·mol-1) concentrations for five months, with maximum shoot length, a number of leaves, and fresh weight reported in 900 µmol·mol-1, and maximum plant height reported in 600 µmol·mol-1 when compared to 380 µmol·mol-1 concentrations. Singh et al. (2016) investigated growth and morphogenesis in the Ocimum basilicum (Tulsi) plant, finding that CO2 enrichment improved growth dynamics (plant height, collar diameter, branches leaves number, root length, above and below-ground biomass) and structure (leaf shape, size, area, and leaf area index) when compared to control conditions (800 µmol·mol-1). Picrorhiza sp. leaf and root proteome responses to drought stress were investigated. Drought stress reduced leaf protein content by 24% and 37% on a fresh weight and dry weight basis, respectively, but drought stress increased root protein content by 12% on a fresh weight basis and decreased protein content by 9% on a dry weight basis (Sangeeta et al., 2014).
For flowering and blooming, the many therapeutic herbs require a chilly temperature. The output of therapeutic herbs has reduced as the temperature has risen. As the temperature rose, so did the production of Delphinium himalayai . It appears that a rise in temperature has had an impact on Delphinium himalayai growth and production. Because Paris polyphylla prefers warmer temperatures to flourish, there was a positive trend in output, indicating that climate change has a favorable impact on P. polyphylla . The number of Swertia chirayita plants has increased as the temperature has risen in recent years. Climate change has a favorable impact on S. chirayita , according to the findings. (Shahi et al. , 2020). Drought stress during the first year of plant growth greatly raised the amount of valerenic acid in V. jatamansi , whereas combined heat and drought stress significantly decreased it. Heat stress reduced valerenic acid production in the aerial component, implying organ-specific changes in plant metabolism under stress. Linalool concentration in H. spicatum increased modestly in drought and several folds with increased heat stress exposure. (Pandey et al. , 2021). Thymus vulgaris (Vurro et al. , 2009) and Thymus hyemalis both showed an increase in essential oil content when exposed to increased CO 2 (500 µmol·mol-1) (Biel et al. , 2005).
Table 1 : Reactive oxygen species and their roles in cellular damages
|
ROS type |
Role in cellular damages |
|
Superoxide anion Peroxide Hydroxyl radicals |
|
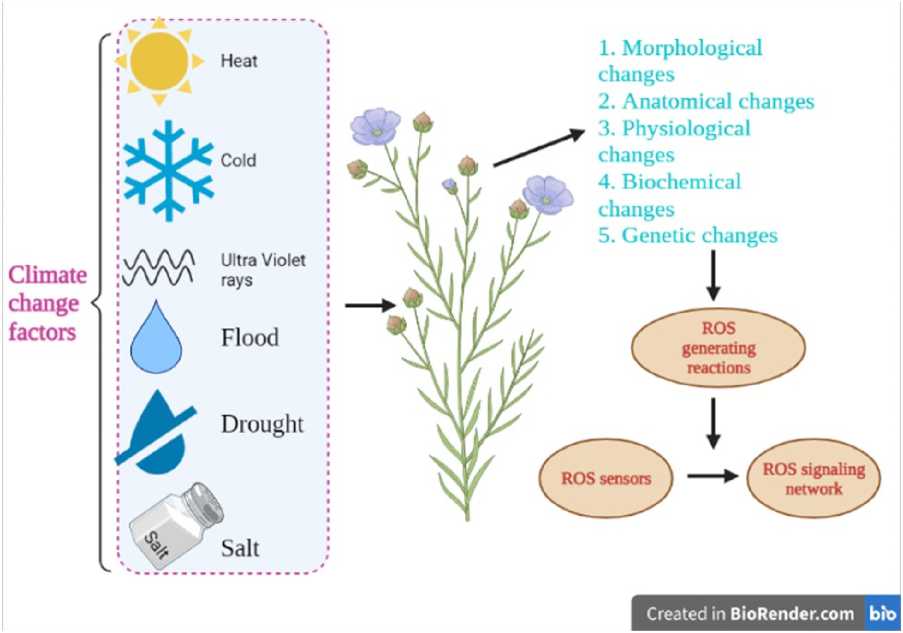
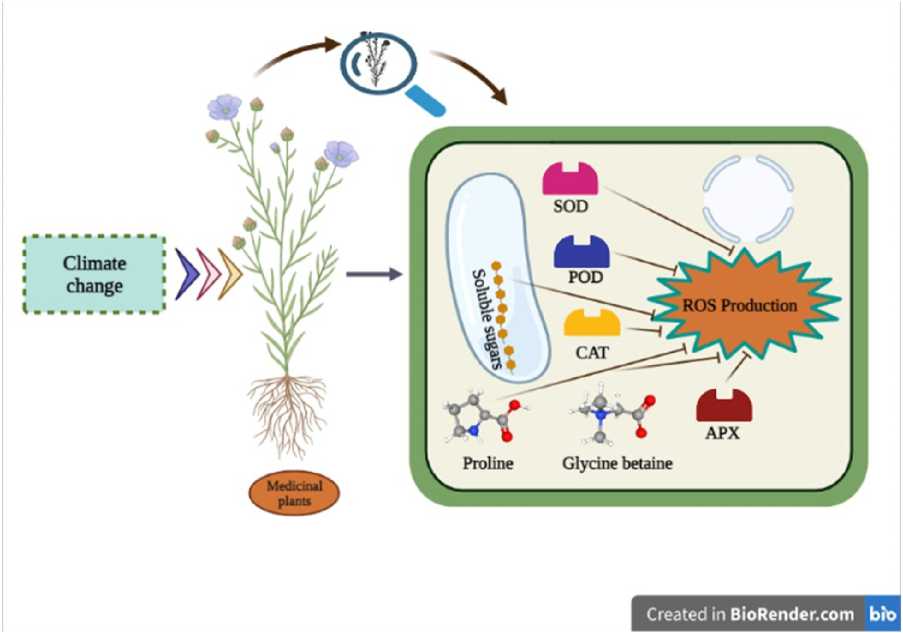
Figure 2. Fusarium head blight symptoms on heads of bread wheat cultivar Bohoth10 inoculated with Fusarium pathogen subjected to varying durations of post-anthesis spray, i.e., 0, 7, 14, 21, and 28 days of spray, under natural climatic conditions compared with negative water control
QUALITY:
Even if a changing environment has no effect on a species' range, it might have a positive or negative impact on its productivity or quality in the case of a medical plant, principally its potency or chemical makeup. While chemical content variations in food plants may be more important to human health than is typically recognized (Welch et al. , 2004., Davis., 2009. Bohn et al. , 2014., Figas et al. , 2015).
Increased environmental pressures may cause changes in chemical content in some species potentially altering the quality or even safety of medical products.
The temperature has a considerable influence on chemical composition in an experimental investigation of Arnica montana (Albert et al. , 2009).
The relationship between altitude and chemical content in bush tea, on the other hand, does not appear to be related to temperature (Nchabeleng et al. , 2012). Medicinal quality will not necessarily improve if montane species whose chemical content is impacted by temperature migrate to higher elevations and hence remain in the same temperature regime, but populations that persist at their original altitudes may fall in quality. Clearly, further research is required to fully comprehend the links between therapeutic potency and elevation in different species.
As proven for American ginseng, high temperatures like drought stress, can result in an elevated concentration of secondary metabolites as a result of drastically lower biomass (Jochum et al. , 2007).
When compared to ambient circumstances, elevated CO 2 increased the essential oil content of V. jatamansi by 17.7%, while elevated temperature lowered it by 4.3 percent. Under Himalayan conditions, an increase in air temperature may not be advantageous to the essential oil content and quality of V. jatamansi in the near future. (Kaundal et al. , 2018)
CONCLUSION
In conclusion, the interplay between photosynthesis environmental stressors, and plant metabolic processes highlights the complex adaptations that plant species undergo in response to changing climatic conditions. Photosynthesis is a fundamental process influenced by plant species and environmental factors, particularly elevated atmospheric CO2 and temperature. Increased CO2 enhances photosynthetic rates while reducing stomatal conductance, promoting plant growth. However, abiotic stresses such as heat and drought significantly impair photosynthesis, as observed in medicinal plants. Canopy temperature regulation and transpiration rates shift under high temperatures affecting water-use efficiency and physiological adaptations. Membrane stability, critical under stress conditions, is compromised by lipid peroxidation and oxidative damage, leading to cellular dysfunction. Antioxidant defense mechanisms, including enzymatic responses, mitigate reactive oxygen species (ROS) accumulation under abiotic stress. Elevated CO2 enhances photosynthesis and growth in several medicinal plants, such as Ocimum sanctum and Gynostemma pentaphyllum, though stressors like drought and heat influence metabolic pathways secondary metabolite accumulation, and overall yield. Notably, medicinal plant quality is altered by climate variables, affecting chemical composition and therapeutic efficacy. While some species benefit from climate-induced changes in metabolite production others exhibit declines in potency. The findings highlight the complex interplay between photosynthesis, stress adaptation, and secondary metabolite biosynthesis under changing environmental conditions, emphasizing the need for further research on climate-resilient medicinal plant cultivation.
CONFLICTS OF INTEREST
The author declare that she has no potential conflicts of interest.
