Пища как фактор плодовитости, продолжительности развития и изменения морфометрических показателей у Henosepilachna vigintioctomaculata (Motschulsky)
")
Автор: Мацишина Наталия Валериевна, Фисенко Петр Викторович, Ермак Марина Владимировна, Собко Ольга Абдулалиевна, Волков Дмитрий Игоревич, Балеевских Антон Геннадьевич
Журнал: Овощи России @vegetables
Рубрика: Защита растений
Статья в выпуске: 5 (61), 2021 года.
Бесплатный доступ
Актуальность. Физиология питания и пищеварения насекомых отражает многообразие их пищевой специализации, направленной на наиболее эффективное использование корма. Выбор объекта питания обеспечивается наличием в нем аттрактантов, а также наличием веществ, необходимых насекомому для прохождения стадий развития и общей питательностью. В свою очередь, у системы «растение-фитофаг» хорошо развиты барьеры, ограничивающие выбор насекомыми растений для их заселения, питания и откладки яиц. В современной литературе очень мало сведений о влиянии пищевых растений на плодовитость, продолжительность развития и изменение морфометрических показателей у двадцативосьмиточечной картофельной коровки Henosepilachna mntioctomacuiata (Motschulsky). Они важны для понимания экологии вредителя и разработки эффективных мер борьбы с ним. Методика исследований. Лабораторная колония Henosepilachna vigintioctomacuiata (Motschulsky) создана в 2019 году. Взрослые особи были собраны в разных местах по всему Приморскому краю (Россия). Для внедрения в культуру инсектария собирали имаго, яйцекладки и личинки в естественных местообитаниях. Использовали стандартные методы содержания и разведения культур насекомых, направленные на оптимизацию параметров среды, плотности содержания и кормовой обеспеченности. Результаты. Выявлено неравномерное влияние культур на разные аспекты онтогенеза картофельной коровки. Наиболее благоприятным кормом в большинстве экспериментов оказался картофель. При питании этой культурой наблюдались наименьший инкубационный период яиц, наибольшие размеры личинок начальных возрастов, наибольшие размеры куколок, плодовитость, низкая смертность, и как следствие выявлены наибольший коэффициент роста (3,22±0,22) и наименьший период развития от яиц до имаго (21,3±0,81). Обнаружено различное влияние культур как на отдельные показатели, так и на онтогенетические периоды фитофага. При питании томатом, огурцом, тыквой, липой была отмечена высокая общая смертность. Липа стимулировала развитие яиц и рост личинок младших возрастов, но провоцировала низкую плодовитость и общее удлинение сроков развития. По совокупности характеристик, наименее благоприятными для питания оказались тыква, огурец и липа.
Фитофаг, картофельная коровка, растительные объекты питания, пасленовые, картофель, онтогенез, морфометрия
Короткий адрес: https://sciup.org/140290362
IDR: 140290362 | УДК: 632.763.79 | DOI: 10.18619/2072-9146-2021-5-81-88
Food as a factor of fertility, development duration, and changes in morphometric parameters in Henosepilachna vigintioctomaculata (Motschulsky)
Relevance. The physiology of insects nutrition and digestion reflects the diversity of their food specialization, aimed at the most effective use of feed. The choice of the object of nutrition is provided by attractants in it, as well as the presence of substances necessary for the insect to pass the stages of development and general nutrition. On the other hand, the "plant-phytophage” system has well-developed barriers that limit the choice of plants by insects for settling, feeding, and eggs laying. In the modern literature, there is very little information on the effect of food plants on fertility, development duration, and changes in morphometric parameters in 28-spotted potato ladybird Henosepilachna vigintioctomaculata (Motschulsky). However, they are important for understanding the ecology of the pest and developing effective control measures. Research methodology. The laboratory colony of Henosepilachna vigintioctomaculata (Motschulsky) was established in 2019. Adults were collected in various locations throughout Primorsky Territory, Russia. For an introduction into the culture of the insectarium, imagos, clutches, and larvae were collected in natural habitats. Standard methods of keeping and breeding insect cultures were used, aimed at optimizing the parameters of the environment, the density of the content, and the feed supply. Results. The uneven influence of crops on different aspects of the ontogenesis of the potato ladybird was revealed. The potato was the most favorable food in most experiments. When feeding with this crop, the smallest incubation period of eggs, the largest sizes of larvae of initial ages, the largest sizes of pupae, fertility, low mortality was observed, and as a result, the largest growth coefficient (3.22±0.22) and the shortest period of development from eggs to imago (21.3±0.81) were revealed. The remaining forage plants used in the study showed a less unambiguous result. The different influence of crops on both individual indicators and the ontogenetic periods of the phytophage was found. When eating tomato, cucumber, pumpkin, lime, high overall mortality was noted. At the same time, linden stimulated the development of eggs and the growth of young larvae but provoked low fertility and a general prolongation of the development period. According to the set of characteristics, pumpkin, cucumber, and linden were the least favorable for nutrition.
Текст научной статьи Пища как фактор плодовитости, продолжительности развития и изменения морфометрических показателей у Henosepilachna vigintioctomaculata (Motschulsky)
Forcitations:Matsishina N.V., Fisenko P.V., Ermak M.V., Sobko O.A., Volkov D.I., Baleevskih A.G. Food as a factor of fertility, development duration, and changes in morphometric parameters in Henosepilachna vigintioctomaculata (Motschulsky). Vegetable crops of Russia. 2021;(5):81-88. (In Russ.)
П ища играет очень важную роль в развитии и выживании любого организма. При этом для насекомых необходимой называется та пища, которая способствует развитию личинок и яйцекладке. Та, которая используется только в качестве источника энергии и продлевает выживание, называется альтернативной [1]. Фитофаги – наиболее активные компоненты в системе «фитофаг-растение». В основе взаимоотношения фитофагов с повреждаемыми ими растениями лежит удовлетворение пищевых потребностей. Физиология питания и пищеварения насекомых отражает многообразие их пищевой специализа-ции,направленной на наиболее эффективное использование корма [2]. Хорошо развитые ноги и крылья в сочетании с высокочувствительной системой органов чувств позволяют насекомым-фитофагам активно выбирать кормовые растения для питания и откладки яиц. Подавляющее большинство видов на имагиналь-ной фазе ведет независимый от растений образ жизни [3]. Взрослые насекомые легко переселяются с растения на растение на большие расстояния в поисках пищи и мест для откладки яиц. В тесном кон- такте с кормовыми растениями насекомые находятся на фазе яйца, личинки и реже – куколки [4]. Установлено, что в обеспечении энергетическими и пластическими веществами, к которым насекомые чрезвычайно требовательны, большое значение имеют соответствие между набором и активностью их пищеварительных ферментов и структурой биополимеров кормовых растений [5]. К тому же, растение используется насекомым в пищу, если оно содержит природные соединения – аттрактанты. Содержание в растениях того или иного аттрактанта в оптимальном для фитофага количестве усиливает интенсивность питания [6]. Предполагается, что у полифагов пищевые реакции вызываются аттрактантами, имеющимися у широкого круга растений. В свою очередь, у системы «растение-фитофаг» хорошо развиты барьеры, ограничивающие выбор насекомыми растений для их заселения, питания и откладки яиц. Эти барьеры приводили и приводят к снижению коэффициента полезного действия пищедобывающей деятельности фитофагов.
В данный момент практически нет сведений о влиянии пищевых растений на плодовитость, продолжительность развития и изменение морфометрических показателей у двадцативосьмиточечной картофельной коровки Henosepilachna vigintioctomaculata (Motschulsky). Однако они важны для понимания экологии вредителя и разработки эффективных мер борьбы с ним, поскольку картофельная коровка – многоядный вредитель. Наиболее сильно страдают от нее картофель, томат, баклажан, кабачок, перец, огурец, тыква, дыня, фасоль. Характер повреждения одинаков для жуков и личинок: они бороздчато скелетируют листья, в результате чего те засыхают. В любой стадии своего развития вредитель поедает мягкую ткань листа, оставляя нетронутыми жилки. Такие листья быстро погибают. В результате урожаи сильно снижаются. Картофельная коровка является переносчиком некоторых болезней картофеля [7], поскольку питается на сорных и культурных растениях-резерва- торах вирусной инфекции и этим также наносит урон [8]. Кроме того, важно знать, как складывались пищевые предпочтения современных вредителей в прошлом, поскольку понимание этих процессов может помочь в прогнозировании возникновения новых патогенных видов. В связи с этим,нами была поставлена серия экспериментов по изучению морфометрических параметров в зависимости от кормового растения.
Материалы и методика
Лабораторная колония Henosepilachna vigintioctomaculata (Motschulsky) создана в 2019 году на базе лаборатории селекционно-генетических исследований полевых культур (ФНЦ агробиотехнологии Дальнего Востока им. А.К. Чайки). Взрослые особи были собраны в разных местах по всему Приморскому краю (Россия). Для внедрения в культуру инсектария насекомых собирали в естественных местообитаниях: на липе Tilia amurensis , черемухе Padus asiatica , картофеле Solanum tuberosum, томате Solanum lycopersicum и баклажане Solanum melongena , отобрав десять имаго разных полов в разных точках изучаемого региона. Отбирались также яйцекладки и личинки младших возрастов. Первый сбор насекомых был проведен в 2019 году, получено восемь лабораторных поколений. В 2020 и 2021 годах в культуру вводили имаго, собранные в природе, чтобы сохранить полиморфизм линий.
Использовали стандартные методы содержания и разведения культур насекомых, направленные на оптимизацию параметров среды, плотности содержания и кормовой обеспеченности [9]. При создании лабораторной популяции учитывали параметры минимальной смертности, минимальной изменчивости форм и максимальной плодовитости. Для создания экологического оптимума культура была стабилизирована, что исключало неконтролируемые факторы и временной дрейф. Также была исключена динамика суточных и сезонных температур и влажности, близкая к естественной. Насекомых выращивали при температуре 25±1,05°C и относительной влажности 85±2,25%, при 16±1,25 часов света в день в тканевых изоляторах. Изоляторы размещали на стойках, подключенных к таймеру (реле времени). Стеллажи были укомплектованы лампами для выращивания растений Quantum line ver. 1 (lm281b + pro 3000K + SMD 5050, 660 нм) (Samsung, Япония). Постоянная температура поддерживалась сплит-системой Rovex RS-07MST1 / RS-07MST1 Aux Air, Китай). Аэрацию как элемент микроклимата обеспечивал аэратор AceLine TFSL-6 (Китай). Уровень влажности контролировали с помощью POLARIS PUH 9105 IQ (Китай). В лаборатории насекомых воспитывали на листьях сорта картофеля Смак, выращиваемого на почве в условиях культуральной комнаты при 25±1,05°C и относительной влажности 85±2,25%, при 16±1,25 ч. световом дне. В то время, когда клубни картофеля нового урожая переживали лечебный и период покоя, насекомых кормили на искусственных питательных средах [10]. Ингредиенты искусственной диеты взвешивали на приборе Сартогосм, Россия (модель МБ 210-А).
Для изучения морфометрических признаков харак- теристик тела были взяты по 25 насекомых различных возрастов (от личинок до имаго) на вариант. Вариантами эксперимента являлись растения-хозяева: картофель, тыква, огурец, фасоль, баклажан, томаты, липа. Измерения длины и ширины яиц, длины и ширины тела разных личиночных возрастов, длины и ширины куколки, длины и ширины головы, груди и надкрылий имаго проводились с использованием стереомикроскопа Nikon SMZ25 и программы NIS-Elements L.
Перед началом постановки эксперимента были собраны свежие яйцекладки с листьев кормовых растений. Яйцекладки содержали в чашках Петри, на фильтровальной бумаге, которую смачивали по мере необходимости. Проводили наблюдения за инкубационном периодом, в результате которых фиксировали процент выводимости для линий. Когорты отродившихся личинок разделяли сразу после отрождения и по десять особей размещали в стеклянные сосуды объемом 80 мл, закрытые плотной бязью. В качестве корма личинкам предлагали свежесобранные листья кормовых растений. Пищу меняли ежедневно. Эксперимент проводили до появления взрослых особей. Данные о продолжительности возрастов личинок, предкуколки, куколки, общего периода развития и репродуктивных фаз самок жука (периода до яйцекладки, яйцекладки и периода после яйцекладки) учитывали отдельно для всех кормовых растений.
Для расчета процента выживаемости личинок и процента появления взрослых особей личинки были разделены на 6 групп по 25 особей в каждой, питающихся основными кормовыми растениями 28-т коровки в Приморском крае: баклажан Solanum melongena , томат Solanum lycopersicum , огурец Cucumis sativus , тыква Cucurbita pepo , картофель Solanum tuberosum , липа Tilia amurensis . Взрослые особи, получившиеся в результате такого содержания, были спарены и разделены после спаривания, чтобы оценить плодовитость самок. Скорость и константа роста, продолжительность инкубационного периода, а также периода дожития были проанализированы с использованием TWOSEX [11]. Полученные результаты обрабатывали статистически и сравнивали с помощью t-критерия Стьюдента. Различия между показателями считали достоверными при p≤0,05. В тексте данные представлены в виде среднего и стандартного отклонения (x±Sx) [12].
Результаты и обсуждение
Проведенные исследования показали, что существенной разницы между длиной и шириной яиц при выкармливании самок различными культурами не было (рис.1). Яйца были типичного для Henosepilachna vigintioctomaculata строения: стебельчатые, блестяще-желтые, удлиненно-овальные, сужающиеся к верхнему концу, с гладкой текстурой, заостренные дистально, сгруппированы вертикально. Потеменения при созревании, которое описывают ряд авторов [13], нами отмечены не были (рис 2.). По нашему мнению,разница в окраске яиц носит скорее морфемный характер и не зависит ни от питания самки, ни от степени созревания. При этом продолжительность инкубационного периода яиц достоверно различалась при питании самок на различных культу-
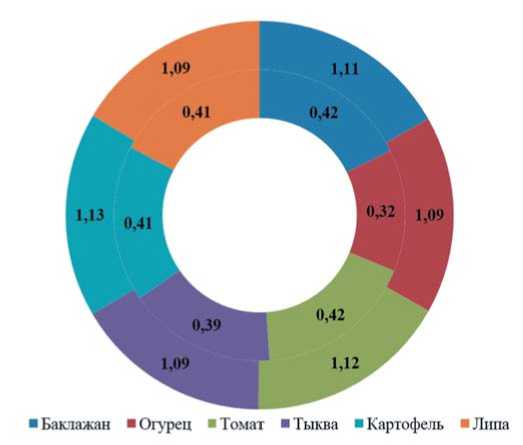
Рис. 1. Соотношение длины и ширины яиц картофельной коровки при питании различными культурами,мм Примечание:по внешнему ряду представлены значения длины,по внутреннему – ширины.
Fig.1. The ratio ofthelength and width ofpotato ladybird eggswhen feeding on variouscrops,mm
Note:theouterrow showsthelength values,theinnerrow shows thewidth values.
рах. Так, инкубационный период на баклажане составил 3,60±0,11 суток, томате – 5,30±1,95, огурце – 5,20±0,87, тыкве – 4,15±0,21, картофеле – 2,58±0,11, липе – 2,01±0,11.
Длина и ширина тела личиночных возрастов также значительно различались. Максимальная длина тела в ранних (I-II) возрастах отмечалась при воспитании на картофеле и липе (табл. 1), в то время, как личинки старших (III-IV) возрастов достигали максимальных значений на томате, огурце, баклажане и тыкве.
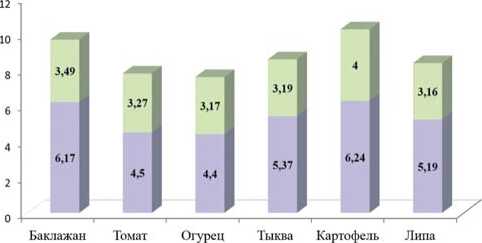
Соотношения длины к ширине тела личинок первого и последнего возраста оставляли для баклажана 2,9:1 и 2,26:1, для огурца – 2,85:1 и 2,4:1, для томата – 2,67:1 и 2,07:1, для тыквы – 2,7:1 и 2,1:1, для картофеля – 3,01:1 и 2,3:1, для липы – 3,5:1 и 2,1:1 соответственно. Измерения длины и ширины тела куколки (рис. 3) достоверно различались у личинок, которых кормили баклажаном и картофелем. Питание на томатах, огурце, тыкве и липе не давало существенной разницы показателей.
Показатели соотношения длины и ширины груди, а также длины и ширины надкрылий имаго картофельной коровки варьировали в пределах видовой нормы, не демонстрируя существенной разницы в зависимости от питания на различных культурах (рис.4).
Следует отметить, что питание на баклажане, томате и картофеле влияло на скорость и константу роста. Максимальные результаты зафиксированы для картофеля, на котором наблюдаются самые высокие показатели этих параметров. Общий период онтогенеза при этом закономерно укорачивался и составлял 21,3±0,81 суток для насекомых, воспитываемых на картофеле, 22,1±0,77 – на баклажане, 23,25±0,61 – на томате.

Рис. 2. Яйца картофельной коровки (фото авторов) Fig. 2. Potato ladybird eggs(photo by theauthors)
Таблица 1. Показатели длины и ширины тела личинок картофельной коровки при питании различными культурами, мм
Table 1. Indicators of the length and width of the body of potato ladybird larvae when feeding on various crops, mm
|
Возраст культура |
Длина, мм |
Ширина, мм |
||||||
|
I |
II |
III |
IV |
I |
II |
III |
IV |
|
|
Баклажан |
1,80±0,04 |
2,93±0,03 |
4,81±0,03 |
6,34±0,09 |
0,62±0,06 |
1,02±0,10 |
2,06±0,02 |
2,80±0,03 |
|
Огурец |
1,14±0,10 |
2,33±0,17 |
4,70±0,40 |
6,60±0,39 |
0,40±0,04 |
0,96±0,12 |
1,89±0,90 |
2,73±0,20 |
|
Томат |
1,90±0,04 |
2,97±0,13 |
4,05±0,27 |
6,80±0,25 |
0,71±0,06 |
1,01±0,07 |
1,79±0,11 |
3,28±0,08 |
|
Тыква |
1,54±0,02 |
2,25±0,02 |
4,06±0,03 |
5,54±0,03 |
0,57±0,02 |
0,94±0,01 |
2,02±0,01 |
2,60±0,03 |
|
Картофель |
2,05±0,27 |
2,88±0,32 |
4,04±0,39 |
5,80±0,70 |
0,68±0,14 |
1,12±0,60 |
1,52±0,34 |
2,52±0,51 |
|
Липа |
2,03±0,27 |
2,52±0,32 |
4,70±0,40 |
5,54±0,03 |
0,57±0,02 |
0,92±0,01 |
2,01±0,01 |
2,60±0,03 |
Средние значения числа яйцекладок (плодовитости) и смертности соотносятся с данными скорости и константы роста. Максимальная плодовитость отмечена на картофеле (735,4±4,90), баклажане (232±7,43) и томате (102±4,90), минимальная – на огурце (72,30±6,13), тыкве (62,30±4,90), липе (42,25±4,90). Самые высокие показатели смертности зафиксированы у особей, питавшихся томатами
(32,7±0,04%), огурцом (30,0±0,04%), тыквой (25,4±0,04%), липой (25,0±0,04%). Самые низкие – на картофеле (10,0±0,04%) и баклажане (10,0±0,04%).
Известно, что плодовитость самок коррелирует с качеством корма [14]. Кроме того, Boczkowska отмечает зависимость плодовитости и развития личинок колорадского жука от сортов картофеля, связывая эту зависимость с анатомическим строением листьев и биохимической спецификой корма [15]. От качества корма также зависит и скорость созревания самок,и длительность яйцекладки [16]. Питание личинок колорадского жука неблагоприятным кормом подтверждает, что успешность прохождения онтогенеза напрямую связана с развитием жирового тела. Его существенное недоразвитие приводит к 100% гибели личинок уже 1-го возраста. При успешном образовании жирового тела развитие личинок и куколок протекает в наиболее короткие сроки и жуки отрождаются большего веса и размера [17][18]. Наименьшая физиологическая подготовленность колорадских жуков связана с их питанием молодым кормом с низким липоцитарным коэффициентом в листьях [19]. Вместе с тем, предпочтения в еде и качество пищи у эпиляхны определяются запахом, вкусом, возрастом растения, а также долей сырой клетчатки, паренхиматозной ткани и содержанием воды [20]. Ранее в наших исследованиях мы показыва- ли, что качество пищи влияет на биологию и экологию как колорадского жука [21,22,23], так и картофельной коровки [24,25 ]. Кроме того, по мнению большинства исследователей, у насекомых, выращиваемых на искусственной диете, могут наблюдаться пониженные уровни экдистероидов, особенно непосредственно перед окукливанием [26]. Исследования зарубежных коллег также подтверждают, что колорадский жук, а значит и картофельная коровка, поскольку им свойственен параллелизм, охотнее поедают старые листья картофеля с более высоким липоцитарным коэффициентом,
■ Дшна. мм Ширина, мм
Рис. 3. Соотношение показателей длины и ширины куколки картофельной коровки при питании различными культурами,мм
Fig. 3. Theratio ofthelength and width ofthe potato ladybird pupa when feeding on variouscrops,mm
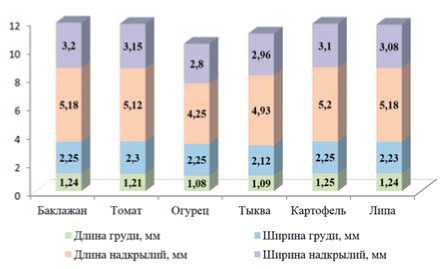
Рис. 4. Показатели соотношения длины и ширины груди,а также длины и ширины надкрылий имаго картофельной коровки при питании различными культурами, мм
Fig.4. Indicatorsofthe ratio ofthelength and width ofthe breast,aswellasthelength and width oftheelytra ofthe potato ladybird imago when feeding on variouscrops,mm
успешнее завершают свой развитие на нём и уходят в диапаузу [27].
Следует отметить, что липу необходимо считать скорее контрольным вариантом, поскольку до появления картофеля на Дальнем Востоке двадцативосьмипятнистая коровка была вредителем лесных насаждений. А.И. Куренцов [28] отмечал, что жуки картофельной коровки держатся в большом количестве на селе-
Таблица 2. Скорость и константа роста личинок картофельной коровки при питании различными культурами Table 2. The growth rate and constant of potato ladybird larvae when feeding on various crops
|
Культура |
Скорость роста |
Константа роста |
Общий период от яиц до имаго, сут. |
|
Баклажан |
0,26±0,02 |
3,12±0,26 |
22,1±0,77 |
|
Томат |
0,26±0,02 |
3,12±0,26 |
23,25±0,61 |
|
Огурец |
0,13±0,01 |
1,56±0,19 |
25,4±0,77 |
|
Тыква |
0,11±0,02 |
1,63±0,19 |
28,20±0,52 |
|
Картофель |
0,32±0,02 |
3,22±0,22 |
21,3±0,81 |
|
Липа |
0,11±0,02 |
1,63±0,10 |
26,3±0,52 |
зеночнике волосистом (Chrysosplechnium pilosum Maxim.). Они сидят по нескольку экземпляров на одном растении, выедая цветы, почки. Жуки встречаются на тех экземплярах этого растения, которые растут по краям лесосек, по сторонам лесных дорог, там, где больше солнечного света. Весенние колонии жуков часто встречаются на цветущих в это время древесных породах, в основном розоцветных (Rosaceae): черемухе азиатской (Padus asiatica Kom.), яблоне маньчжурской (Malus manshurica (Maxim.) Kom.), боярышнике Максимовича (Crategus maximowiczii Schneid.), груше уссурийской (Pyrrus ussuriensis Maxim.), лесных травах (Chrysospenium pilosum Maxim., Schizopepon bryoniifolius Maxim.) и затем переходят на более характерные и часто встречающиеся сорняки полей: осот полевой (Sonchus arvensis L.), паслен черный (Solanum nigrum L.). Жуки проходят дополнительное питание пыльцой на цветках черемухи, яблони, боярышника. С последних растений коровка переходит на картофель [29,30], но может питаться листьями огурца, томата, баклажана. По нашему предположению, коллеги имели возможность наблюдать процессы, сходные с процессами адаптации колорадского жука к новому корму. Как известно, до ввоза в США культурного картофеля жук питался дикими пасленовыми. Его главными кормовыми растениями были паслен колючий – S. rostratum и S. cornutum. Встречался он также на S. robustum, S. carolinense, S. jasminoides и других. Северной границей распространения жука на Американском континенте, кроме температуры, было распространение его кормовых растений, в том числе диких пасленовых – S. triflorum и S. cornutum. В юго-западные районы США (штаты Колорадо и Небраска) культурный картофель – S. tuberosum был завезен в конце первой половины XIX в. и вскоре стал основным кормовым растением колорадского жука [31]. После перехода на питание картофелем первоначальный барьер распространения вида – недостаток корма – был устранен и колорадский жук стал быстро распространяться по Северной Америке – за 20 лет (1860-1880 годы) им была заражена площадь более 6 млн. км2. Обильная кормовая база способствовала не только расширению ареала вредителя; с переменой кормового растения увеличился потенциал размножения и значительно повысилась экологическая пластичность вида, что, в свою очередь, облегчало ему продвижение в новые районы [32]. Предположение о наличии в кормовом растении колорадского жука веществ, стимулирующих его питание, было высказано уже в 1933-1936 годах. К ним относятся сахара, аминокислоты, некоторые липиды. Из сахаров наиболее выраженную реакцию кусания и поедания вызывает сахароза в концентрациях 0,025-0,1 мк. При концентрации выше 0,1 мк эта реакция слабеет. Кроме сахарозы более слабую положительную реакцию вызывают такие сахара, как глюкоза, фруктоза и манноза. В отличие от других насекомых-фитофагов, которые сильнее реагируют на смесь сахаров, на личинок жука активнее действует одна сахароза [33]. Некоторые аминокислоты являются также хорошими стимуляторами питания колорадского жука. Большая часть аминокислот, вызывающих реакцию питания, относится к группе алифатических. Наиболее актив- ными из них оказались L-аланин, γ-аминомасляная кислота и L-серин. Из аминокислот, содержащих серу, небольшой стимулирующий эффект дает только L-цистеин. Исследования показали, что обязательным условием эффективности аминокислот является их растворимость в воде и, по-видимому, размеры молекул (молекулярный вес аминокислот, стимулирующих питание, не превышал 125) [34]. Полагаем, что всё это актуально и для картофельной коровки. Более питательный, чем дикоросы, картофель оказался гораздо привлекательнее для этого широкого полифага, оставляя привычные кормовые растения в качестве зимовочных биотопов. Наши исследования это подтверждают. Мы видим, что на картофеле, нежели на липе, наблюдается большая выживаемость, жуки и личинки значительно крупнее, самки плодовитее.
Заключение
Хотя картофельная коровка и является широким полифагом, показано значительное влияние различных кормовых растений на протекание онтогенеза и морфометрических изменений в нем. Выявлено неравномерное влияние культур на разные аспекты онтогенеза картофельной коровки. Наиболее благоприятным кормом в большинстве экспериментов оказался картофель. При питании этой культурой наблюдали наименьший инкубационный период яиц, наибольшие размеры личинок начальных возрастов, наибольшие размеры куколок, плодовитость, низкая смертность, и как следствие, выявлены наибольший коэффициент роста (3,22±0,22) и наименьший период развития от яиц до имаго (21,3±0,81). Исключением оказались личинки старших возрастов, наибольшие размеры которы, выявлены при питании другими культурами, в частности, баклажаном, при питании которым также наблюдалась низкая смертность и высокая плодовитость. Остальные кормовые растения, использованные в исследовании, показали менее однозначный результат. Обнаружено различное влияние культур как на отдельные показатели, так и на онтогенетические периоды фитофага. При питании томатом, огурцом, тыквой, липой была отмечена высокая общая смертность. В то же время, липа стимулировала развитие яиц и рост личинок младших возрастов, но провоцировала низкую плодовитость и общее удлинение сроков развития. Наиболее неоднозначная картина наблюдалась при питании коровки на томате. С одной стороны, наблюдались наибольший инкубационный период яиц и высокая смертность личинок, с другой, относительно высокая плодовитость, стимулирование роста личинок старших возрастов, а также средние значения константы роста (3,12±0,26) и общего времени онтогенеза (23,25±0,61). Положительное влияние томата можно объяснить высоким содержанием аттрактантов и общей питательностью, в то время как высокое содержание специфического алкалоида томатина объясняет негативные проявления при питании данной культурой. Столь различное и неравномерное влияние различных пищевых растений на разных стадиях онтогенеза может свидетельствовать о возможной сложной схемы смены пищевых растений (и биотопов) у коровки в зависимости от стадий онтогенеза и/или фенологических периодов в прошлом, до массового появления культурного картофеля в пределах ареала ее обитания.
Об авторах:
Nathalia V. Matsishina – Cand. Sci. (Biology), Senior Researcher laboratory of selection and genetic research of field crops, ,
Petr V. Fisenko – Cand. Sci. (Biology), Leading Scientist, Acting Head of laboratory of selection and genetic research of field crops, ,
Marina V. Ermak – Junior Researcher of laboratory
of selection and genetic research of field crops, ,
Olga A. Sobko – Postgraduate Student, Junior Researcher of laboratory of selection and genetic research of field crops, ,
Dmitry I. Volkov – Postgraduate Student, Head of Department of Potato and Vegetable Growing, ,
Anton G. Baleevskih – Laboratory Assistant of laboratory of selection and genetic research of field crops, ,
Список литературы Пища как фактор плодовитости, продолжительности развития и изменения морфометрических показателей у Henosepilachna vigintioctomaculata (Motschulsky)
- Hodek I. Ecology of Coccinellidae. Kluwer Academic Publishers. Boston: Dordrecht, 1996.
- Фасулати С.Р. Формирование внутривидовой структуры у насекомых в условиях агроэкосистем на примерах колорадского жука Leptinotarsa decemlineata Say (Coleoptera, Chrysomelidae) и вредной черепашки Eurygaster integriceps Рut. (Heteroptera, Scutelleridae) Науковий вiсник Ужгородського унiверсiтету. Серiя Бiологiя. Ужгород. 2010;(29):13-27.
- Heiser C. Nightshades: the Paradoxical Plants. San Francisco, CA: W.H. Freeman and Company, 1969. 200 p.
- Павлюшин В.А., Вилкова. Н.А, Сухорученко Г.И., Нефедова Л.И., Фасулати С.Р. Фитосанитарная дестабилизация агроэкосистем. СПб.: Родные просторы, 2013. 184 с.
- Giordanengo P., Vincent C., Alyokhin A. Insect Pests of Potato: Global Perspectives on Biology and Management. Academic Press is an imprint of Elsevier, 2013. 598 p.
- Capinera J.L. Handbook of Vegetable Pests. San Diego, CA: Academic Press, 2001. 800 p.
- Смирнов, Ю.В. Анализ фитосанитарного риска картофельной коровки Henosepilachna vigintioctomaculata (Motsch.) для территории Российской Федерации. Москва, 2010. 49 с.
- Sobko, O.A., Matsishina, N.V., Fisenko, P.V., Kim, I.V., Didora, A.S., Boginskaya, N.G. & Volkov, D.I. Viruses in the agrobiocenosis of the potato fields. IOP Conference Series: Earth and Environmental Science. 2021;677(5):article 052093.
- Злотин А.З. Техническая энтомология. Киев: «Наукова думка», 1989. 183 с.
- Wang, Z.L., Wang X.P., Li C.R., Xia Z.Z., Li. S.X. Effect of dietary protein and carbohydrates on survival and growth in larvae of the Henosepilachna vigintioctopunctata (F.) (Coleoptera: Coccinellidae). Journal of Insect Science. 2018;18(4):1-7. https://doi.org/10.1093/jisesa/iey067
- Chi, H. TWOSEX-MSChart: a computer program for the age-stage, two-sex life table analysis. National Chung Hsing University, Taichung, Taiwan. 2016. http://140.120.197.173
- McDonald, J.H. Handbook of biological statistics. Third Edition. Sparky House Publishing, Baltimore, Maryland, USA. 2014. 305 pp.
- Hossain M A, El-Taj H F, Haque M A, Ara A, Uddin M N. Biology, food consumption and natural enemies of Epilachna beetle, Epilachna dodecastigma (Wied). Int J Sust Agri 2008;4(2):59-64.
- Венгорек В. Влияние продолжительности дня и качества корма на биологию колорадского жука (Leptinotarsa decemlineata Say). Колорадский жук и меры борьбы с ним. М.: Изд-во АН СССР, 1958;(2):129-135.
- Boczkowska M. Observations suruneseconde generation de Leptinotarsa decemlineata Say obtenusurSolanumdenissum. IXth International Congress of Entomology. Amsterdam, Aug. 17-24. 1951;(1).660.
- Ларченко К.И. Фенологические сроки развития колорадского жука (Leptinotarsa desemlineata Say) и их связь с расселением и изменением численности. Колорадский жук и меры борьбы с ним. М.: Изд-во АН СССР,1958. С.106-115.
- Ушатинская Р.С. Некоторые физиологические и биохимические особенности диапаузы колорадского жука (Leptinotarsa decemlineata Say). Колорадский жук и меры борьбы с ним. M.: Изд. АН СССР, 1958. С.150- 186.
- Jones C G, Hoggard M P, Blum M S. Pattern and process in insect feeding behavior: a quantitative analysis of the Mexican bean beetle, Epilachna varivestis. Entomologia Experementalis et Applicata.1981;30(3):254-264. https://doi.org/10.1111/j.1570-7458.1981.tb03108.x
- Kaur R, Mavi GS. Biology of Epilachna vigintioctopunctata (Fabricius) (Coleoptera: Coccinellidae) on brinjal in Ludhiana, Punjab. Crop Research. 2005;29(1):141-144.
- Мацишина Н.В. К вопросу устойчивости сортов картофеля традиционной селекции к колорадскому жуку Leptinotarsa decemlineata Say, 1824 (Coleoptera, Chrysomelidae) в условиях муссонного климата Приморского края. Овощи России. 2015;(2):80-83. https://doi.org/10.18619/2072-9146-2015-2-80-83
- Мацишина Н.В., Рогатных Д.Ю. Инвазия колорадского жука на Дальнем Востоке. Вестник защиты растений. 2013;(4):64-68.
- Мацишина Н.В.К Биологии колорадского жука Leptinotarsa decemlineata Say, 1824 (Coleoptera, Chrysomelidae) на Юге Дальнего Востока России. Евразиатский энтомологический журнал. 2011;10(3):330-336.
- Мацишина Н.В., Шайбекова А.С., Богинская Н.Г., Собко О.А., Волков Д.И., Ким И.В. Предварительная оценка сортов картофеля отечественной и зарубежной селекции на устойчивость к картофельной двадцативосьмиточечной коровке Henosepilachna vigintioctomaculata Motch. (Fabricius, 1775) в Приморском крае. Овощи России. 2019;(6):116-119. https://doi.org/10.18619/2072-9146-2019-6-116-119
- Мацишина Н.В., Фисенко П.В., Собко О.А. Морфологические аномалии в онтогенезе картофельной коровки Henosepilachna vigintioctomaculata (Motschulsky, 1857) (Coleoptera:Coccinellidae). Вестник Дальневосточного отделения Российской академии наук. 2021;3(217):57-62. https://doi.org/10.37102/0869-7698_2021_217_03_09
- Gelman D. B, Carpenter J. E., Greany P. D. Ecdysteroid levels/profiles of the parasitoid wasp, Diapetimorpha introita, reared on its host, Spodoptera frugiperda and on an artificial diet. Journal of Insect Physiology. 2000;46(4):457-465. https://doi.org/10.1016/S0022-1910(99)00132-8
- Noronha Ch., Cloutier C. Effects of Potato Foliage Age and Temperature Regime on Prediapause Colorado Potato Beetle Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Environmental Entomology. 2006;35(3):590- 599. https://doi.org/10.1603/0046-225X-35.3.590
- Куренцов А.И. Новые данные по биологии картофельной коровки. Тр. Горнотаежной станции Дальневосточного филиала Академии наук СССР. Владивосток, 1946;(5):257-266.
- Куренцов А.И. Проблема сельскохозяйственного освоения горнотаежных районов в Приморском крае и вредные насекомые. Тр. Горнотаежной станции Дальневосточного филиала Академии наук СССР. ВорошиловУссурийский, 1941;(4):15-17.
- Кузнецов В.Н. Зоогеографический анализ фауны кокцинеллид (Coleoptera, Coccinellidae) Приморского края. Энтомофаги Советского Дальнего Востока. Владивосток, 1975. С.153-155.
- Hare J D. Ecology and management of the Colorado potato beetle. Annual Review Entomology. 1990;(35):81-100. https://doi.org/10.1146/annurev.en.35.010190.000501
- Hoffmann M.P., Frodsham A.C. Natural Enemies of Vegetable Insect Pests. Ithica: Cornell University, 1993. 63 p.
- Ferro, D.N., J.A. Logan, R.H. Voss, and Elkinton J.S. Colorado potato beetle (Coleoptera: Chyrsomelidae) temperature-dependent growth and feeding rates. Environmental entomology. 1985;14(3):343-348.
- Domek J.M., Cantelo W.W., Deahl K.L. A meridic diet for the Colorado potato beetle. Journal of Entomological Science. 1997;32(4):430-444. https://doi.org/10.18474/0749-8004-32.4.430
- Kowalski, S.P., Domek, J.M., Deahl, K.L., Sanfordl L. L. Performance of Colorado potato beetle larvae, Leptinotarsa decemlineata (Say), reared on synthetic diets supplemented with Solanum glycoalkaloids. American Journal Potato Research.1999;76(5):305-312. https://doi.org/10.1007/BF02853629
