Питание гоголей Bucephala clangula в Кандалакшском заливе

Автор: Бианки В.В., Бойко Н.С., Хайтов В.М.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 225 т.12, 2003 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140149648
IDR: 140149648
The food of common goldeneyes Bucephala clangula in the Kandalaksha bay
Текст статьи Питание гоголей Bucephala clangula в Кандалакшском заливе
Гоголь Bucephala clangula — утка, гнездящаяся на таёжных водоёмах, а изредка и на морских побережьях. В зависимости от места обитания, гоголи проводят полную летнюю смену оперения на пресных водоёмах (например, на юге Западной Сибири) или на морских мелководьях (например, на Белом море). Зимует большинство гоголей на незамерзающих озёрах, участках рек (например, истоки Ангары) и на мелководьях морей (например, Датские проливы). Те птицы, которые часть года проводят на море, дважды в году полностью меняют свой рацион, переходя с пресноводных беспозвоночных на морских и обратно.
В Кандалакшском заливе гоголи заселяют искусственные гнездовья на заповедных островах Кандалакшских шхер (рис. 1), а также используют морскую акваторию во время летней линьки и перед отлётом на зимовку. На морских мелководьях они находят более обильную пищу, чем на материковых водоёмах, и испытывают меньше беспокойства от хищников и людей. Это позволяет им собираться здесь как небольшими, так и многосотенными стаями.
Вершина Кандалакшского залива Белого моря
• Точки взятия дночерпательных проб
Рис. 1. Картосхема района проведения исследований.
Таким образом, гоголь по своему строению и, соответственно, питанию стоит между нырковыми утками и крохалями, которые адаптированы прежде всего к питанию рыбой. По строению клюва и пищеварительного тракта, а также по составу кормов, к гоголю близка морянка Clangula hyemalis, гнездящаяся в тундре.
Ныряют гоголи неглубоко, предпочитая глубины меньше 5 м. Они питаются, как правило, животными “среднего” размера, которых берут поштучно, не прибегая к щелокчению. По нашим наблюдениям в Кандалакшском заливе (Бианки, Бойко, Шутова 1995) гоголи предпочитают мидий Mytilus edulis с длиной раковины 2-12 , в среднем 7.7±0.2 мм (Игл 1.5-19; и = 216). Зимой у побережий Дании величина их добычи обычно не превышает в длину 12 мм для моллюсков, 40 мм для креветок, но может доходить до 150 мм в случае использования в пищу миног (Madsen 1954). Величина дрейссен Dreissena polymorpha, поедаемых зимой на оз. Унтер-зе в предгорьях Альп, составляет 0.5-16, в среднем 4.0 мм (Suter 1982).
Многочисленные публикации о питании гоголей в разные сезоны и в разных местах ареала говорят о большом разнообразии их рациона (Немцев 1956; Зиновьев 1973; Брагин 1974; Bauer, Glutz v. Blotzheim 1969; Suter 1982; Blumel, Krause 1990; Семенов-Тян-Шанский, Гилязов 1991, Бианки и др. 1995; Бианки 1999; и др.). Будучи обитателем пресноводных таёжных водоёмов, он предпочтительно использует в пищу личинок ручейников Trichoptera, а также звонцов Chironomidae, подёнок Ephemeroptera, стрекоз Odonata и других, имаго водных жуков Coleoptera, гребляков Corixidae, равноногих ракообразных Isopoda, а также мелких моллюсков Mollusca, рыбок Osteichthyes и иных. Во время линьки, миграции и зимовки на морских мелководьях гоголи питаются мелкими моллюсками, бокоплавами, многощетинковыми червями Polycheta, другими беспозвоночными эпифауны и мелкой рыбой. Из растительности они используют семена рдеста Pota-mogeton, урути Myriophyllum, горца Poligonum scabrum (= Persicaria tomentosa), ежеголовника Sparganium simplex и других водных и околоводных растений, реже их вегетативные части. Порой вегетативные части водных растений они заглатывают случайно, вместе с обитающими на них животными.
Целью нашего исследование было изучение питания гоголя на морских и пресноводных акваториях в вершине Кандалакшского залива Белого моря.
Материал и методика
При отлавливании насиживающих самок в конце мая - июне мы получали от них порцию экскрементов, которая составляла одну пробу (табл. 1). В 19941999 гг. полученные пробы экскрементов высушивались, в 1999-2002 гг. фикси-
Таблица 1. Количество капрологических проб, полученных от насиживающих самок в разные годы
Выяснение пищевого рациона по анализу помёта имеет значительные преимущества, будучи прижизненной методикой, позволяющей неоднократно получать данные от одной и той же особи, не принося ей вреда. Существенный недостаток этой методики заключается в сильном изменении объектов питания при прохождении через пищеварительный тракт, а некоторые “мягкие” объекты могут не сохраняться при этом совсем.
Для выявления наиболее часто поедаемых видов корма мы использовали анализ частотного распределения встречаемости. Те виды корма, встречаемость которых формировала отдельные пики в области высоких значений, рассматривались как наиболее частые.
Для описания места кормежки гоголей в море в двух точках их обычного пребывания (рис. 1) в сентябре 2001 были взяты по три пробы дночерпателем Петерсена с площадью захвата 1/40 м2. После промывки проб через сито с диаметром ячеи 1 мм, их разбирали в кюветах. Из проб были выбраны все животные и растения, которых, после определения, подсчитали и взвесили. Вес определяли с точностью до 1 мг после обсушивания на фильтровальной бумаге. Дополнительную информацию мы получили при взятии нескольких качественных проб шлюпочной драгой близ острова Большой Ломнишный (рис. 1), где также держатся гоголи.
Результаты
Концентрация гоголей на морской акватории Кандалакшских шхер происходит в тех местах, где глубины не превышают 5 м. На этих глубинах формируются многочисленные мидиевые банки и заросли бурой водоросли Laminaria saccharina. Качественные дражные пробы показали, что в местах скопления птиц располагаются мощные сплетения нитчатых водорослей Desmarestia aculeata, растущих на ризоидах ламинарий, которые прикрепляются к отдельным камням, вкрапленным в илистые поля.
Точки, в которых были взяты количественные дночерпательные пробы, располагались в окрестностях довольно мощных мидиевых банок. В них было отмечено 24 таксона беспозвоночных (табл. 2). Эти выборки незначительно отличаются по видовому составу за счёт малочисленных видов. В обеих точках самыми обильными организмами оказались мидии, представленные, главным образом, молодыми особями, живущими среди нитчатых водорослей, биомасса которых уступает только биомассе мидий. По визуальным наблюдениям нитчатые водоросли покрывают здесь от 50 до 100% грунта. Меньшую биомассу, но довольно высокую численность в этих биотопах имеют полихеты Polydora quadrilobata, молодь гастропод рода Littorina, а также моллюски Epheria vincta, Hydrobia ulvae, Macoma balthica и личинки комаров Cricotopus vitripennis. 1
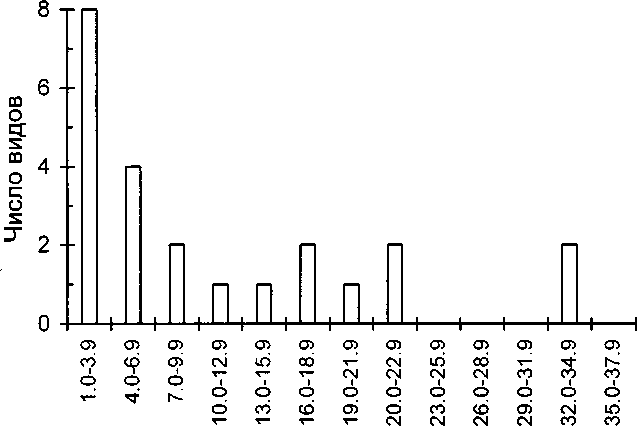
Анализ капрологических данных позволил выявить 36 разновидностей объектов (табл. 3). Из 35 биологических объектов наибольшего разнообразия достигали представители членистоногих —16 объектов, 85.8% и 55.0% средней доли. Из них 34.5% и 21.2% соответственно составляли бокоплавы и 23.0% и 11.8% — ручейники, а также 17.7% и 6.2% — Cricotopus vitripennis и 15.9% и 4.3% — жуки и клопы.
На рисунке 2 видно, что частотное распределение встречаемости видов корма имеет полимодальный характер. Несколько пиков в распределении свидетельствуют о наличии нескольких группировок видов, отличающихся по своей встречаемости. Первая группа (менее 13%) — это виды довольно редкие и, возможно, относительно случайно попадающие в пищу птиц. Две другие группы, соответствующие встречаемости видов равной 13-23% и 3235%, характеризуют значительную часть рациона в местах гнездования. В эту группу попадают: Mytilus edulis, Gammaridea, Osteichthyes, Trichoptera, Cricotopus virtipennis, Hydrobia ulvae и растительные остатки. По величине средней доли наиболее важными компонентами являются: Gammaridea, Mytilus edulis, Trichoptera, Osteichthyes, Cricotopus vitripennis. Если же рыб объединить с их икрой, то они выходят на второе место.
Таблица 2. Средняя плотность поселения (Л/, экз./м2) и средняя биомасса (В, г/м2) видов бентоса на местах кормежки гоголей около острова Ряшкова
|
Вид |
У Восточной косы |
У Западного берега |
Средние по двум акваториям |
|||||||||||||||
|
N |
в |
N |
в |
N |
в |
|||||||||||||
|
Halicryptus spinulosus |
13 |
+ |
13.3 |
3.33 |
+ |
3.333 |
27 |
+ |
26.7 |
0.13 |
+ |
0.133 |
20 |
+ |
13.7 |
1.73 |
+ |
1.655 |
|
Nemertini |
740 |
+ |
283.5 |
1.23 |
+ |
0.831 |
714 |
+ |
577.1 |
7.18 |
+ |
3.369 |
727 |
+ |
287.6 |
4.21 |
+ |
2.044 |
|
Pygospio elegans |
27 |
+ |
26.7 |
0.03 |
+ |
0.027 |
147 |
+ |
109.1 |
0.11 |
+ |
0.071 |
87 |
+ |
57.0 |
0.07 |
+ |
0.038 |
|
Polydora quadrilobata |
1854 |
+ |
1059.3 |
1.70 |
+ |
1.135 |
320 |
+ |
227.4 |
0.20 |
+ |
0.144 |
1087 |
+ |
593.6 |
0.95 |
+ |
0.612 |
|
Harmothoe imbricata |
128 |
+ |
128.5 |
1.55 |
+ |
1.551 |
13 |
+ |
13.3 |
0.07 |
+ |
0.067 |
71 |
± |
63.2 |
0.81 |
+ |
0.770 |
|
Phyllodoce groenlandica |
13 |
+ |
13.3 |
0.13 |
+ |
0.133 |
— |
— |
— |
— |
7 |
+ |
6.7 |
0.07 |
+ |
0.067 |
||
|
Eteone longa |
— |
— |
— |
— |
13 |
+ |
13.3 |
0.04 |
+ |
0.040 |
7 |
+ |
6.7 |
0.02 |
+ |
0.020 |
||
|
Fabricia sabella |
27 |
+ |
26.7 |
2.16 |
+ |
2.160 |
107 |
+ |
70.6 |
0.05 |
+ |
0.035 |
67 |
+ |
38.2 |
1.11 |
+ |
1.075 |
|
Tubificoides benedeni |
94 |
+ |
94.0 |
0.09 |
+ |
0.094 |
133 |
+ |
70.6 |
0.12 |
+ |
0.061 |
114 |
+ |
53.3 |
0.11 |
+ |
0.050 |
|
Littorina saxatilis |
248 |
+ |
125.2 |
7.65 |
+ |
5.939 |
359 |
+ |
339.4 |
10.06 |
+ |
9.859 |
304 |
+ |
163.7 |
8.85 |
+ |
5.176 |
|
Littorina obtusata |
— |
— |
— |
— |
27 |
+ |
13.3 |
4.63 |
+ |
4.042 |
13 |
+ |
8.4 |
2.31 |
+ |
2.083 |
||
|
Littorina sp. —juv. |
1574 |
+ |
1510.0 |
1.99 |
+ |
1.885 |
1078 |
+ |
938.1 |
1.14 |
+ |
0.728 |
1326 |
+ |
802.7 |
1.57 |
+ |
0.924 |
|
Hydrobia ulvae |
378 |
+ |
222.7 |
2.75 |
+ |
1.612 |
4097 |
+ |
2921.8 |
37.85 |
+ |
28.247 |
2237 |
+ |
1552.1 |
20.30 |
+ |
14.890 |
|
Onoba aculeas |
142 |
+ |
122.4 |
0.45 |
+ |
0.357 |
— |
— |
— |
— |
71 |
+ |
63.2 |
0.23 |
+ |
0.189 |
||
|
Epheria vincta |
7332 |
+ |
6448.8 |
58.53 |
+ |
53.799 |
124 |
+ |
84.2 |
0.90 |
+ |
0.720 |
3728 |
+ |
3304.0 |
29.71 |
+ |
27.295 |
|
Mytilus edulis |
531307 |
+ |
311604.3 |
2829.64 |
+ |
1958.67 |
81291 |
+ |
65202.2 |
2092.5 |
+ |
1903.9 |
306299 |
+ |
174342.9 |
2461.07 |
+ |
1232.6 |
|
Macoma balthica |
511 |
+ |
356.8 |
5.39 |
+ |
5.007 |
1848 |
+ |
567.7 |
37.00 |
+ |
12.301 |
1179 |
+ |
423.4 |
21.19 |
+ |
9.232 |
|
Atylus carinatus |
709 |
+ |
669.9 |
5.08 |
+ |
3.941 |
67 |
+ |
13.3 |
2.53 |
+ |
0.035 |
388 |
+ |
332.3 |
3.81 |
+ |
1.852 |
|
Jaera sp. |
13 |
+ |
13.3 |
0.01 |
+ |
0.013 |
13 |
+ |
13.3 |
0.01 |
+ |
0.013 |
13 |
+ |
8.4 |
0.01 |
+ |
0.008 |
|
Orchominella minuta |
13 |
+ |
13.3 |
0.03 |
+ |
0.027 |
— |
— |
— |
— |
7 |
+ |
6.7 |
0.01 |
+ |
0.013 |
||
|
Gammaridea —juv. |
— |
— |
— |
— |
27 |
+ |
13.3 |
0.03 |
+ |
0.013 |
13 |
+ |
8.4 |
0.01 |
+ |
0.008 |
||
|
Diastylis glabra |
— |
— |
— |
— |
13 |
+ |
13.3 |
0.39 |
+ |
0.387 |
7 |
+ |
6.7 |
0.19 |
+ |
0.193 |
||
|
Cricotopus vitripennis |
10638 |
+ |
8290.1 |
2.56 |
+ |
1.563 |
133 |
+ |
58.1 |
0.08 |
+ |
0.023 |
5386 |
+ |
4389.0 |
1.32 |
+ |
0.892 |
|
Asterias rubens |
121 |
+ |
120.7 |
0.21 |
+ |
0.215 |
13 |
+ |
13.3 |
0.01 |
+ |
0.013 |
67 |
+ |
59.4 |
0.11 |
+ |
0.106 |
|
Fucus vesiculosus |
— |
— |
— |
— |
— |
— |
60.00 |
+ |
60.000 |
— |
— |
30.00 |
± |
30.00 |
||||
|
Вес нитчатых водорослей |
1992.33 |
+ |
1353.82 |
432.73 |
+ |
340.26 |
1212.53 |
+ |
715.08 |
|||||||||
Таблица 3. Встречаемость (%) и средняя доля (%) различных пищевых объектов в помёте насиживающих самок гоголя
|
Объекты |
Происхождение |
Встречаемость |
Средняя доля |
|
Foraminifera |
М |
2.7 |
+ |
|
Nereis sp. |
М |
0.9 |
0.02 |
|
Arenicola marina |
М |
3.5 |
1.06 |
|
Spirorbidae |
М |
0.9 |
+ |
|
Polynoidae |
М |
0.9 |
0.71 |
|
Hydrobia ulvae |
М |
16.8 |
3.02 |
|
Littorina sp. |
М |
4.4 |
0.19 |
|
Skeneopsis planorbis |
М |
0.9 |
+ |
|
Mytilus edulis |
М |
36.3 |
13.14 |
|
Mya arenaria |
М |
1.8 |
0.93 |
|
Musculus sp. |
М |
0.9 |
0.02 |
|
Macoma balthica |
М |
0.9 |
0.62 |
|
Gammaridea gen, sp. |
? |
34.5 |
21.19 |
|
Harpacticoidea |
М |
0.9 |
+ |
|
Ostracoda |
М |
2.7 |
+ |
|
Balanus sp. |
м |
0.9 |
+ |
|
Halacarina |
м |
1.8 |
+ |
|
Cricotopus vitripennis |
м |
17.7 |
6.20 |
|
Chironomidae (larvae) |
? |
5.3 |
3.05 |
|
Brachicera (larvae) |
? |
0.9 |
0.04 |
|
Culicidae (imago) |
н |
0.9 |
0.27 |
|
Nematocera (imago) |
н |
0.9 |
0.04 |
|
Trichoptera (larvae) |
п |
23.0 |
11.84 |
|
Trichoptera (imago) |
н |
0.9 |
0.04 |
|
Odonata (Zygoptera, larvae) |
п |
2.7 |
1.68 |
|
Ephemeroptera (larvae) |
п |
1.8 |
1.05 |
|
Sigara sp. |
п |
4.4 |
1.11 |
|
Coleoptera (imago) |
п |
11.5 |
3.24 |
|
Formicidae |
н |
1.8 |
0.49 |
|
Aphidinea (imago) |
н |
0.9 |
+ |
|
Insecta, ближе не опред. |
? |
14.2 |
4.92 |
|
Osteichthyes |
? |
23.9 |
8.77 |
|
Икра рыбы |
? |
8.8 |
6.45 |
|
Нитчатые водоросли |
? |
8.8 |
+ |
|
Растительные остатки |
? |
19.5 |
3.71 |
|
Песок |
? |
5.3 |
+ |
Обозначения:М — морские объекты; П — пресноводные объекты; Н — наземные объекты; “?” — происхождение неопределённо. “+” — доля вида ничтожна.
Встречаемость, %
Рис. 2. Частотное распределение встречаемости различных видов корма в помёте насиживающих самок гоголя.
Встречаемость, %
|
Виды корма |
Встречены в помёте |
Встречаемость |
Средняя доля |
|
Hydrozoa |
— |
7.5 |
0.2 |
|
Polynoidae gen. sp. |
+ |
2.5 |
0.3 |
|
Nereis sp. |
+ |
25.0 |
2.9 |
|
Piste maculate |
— |
2.5 |
2.0 |
|
Arenicola marina |
+ |
2.5 |
1.3 |
|
Tonicelle marmorea |
— |
2.5 |
0.3 |
|
Testudinalia tesselata |
— |
2.5 |
1.7 |
|
Littorina sp. |
+ |
5.0 |
0.8 |
|
Onoba sp. |
— |
2.5 |
0.3 |
|
Epheria vincta |
— |
5.0 |
0.2 |
|
Margarites helicinus |
— |
2.5 |
0.0 |
|
Mytilus edulis |
+ |
80.0 |
54.9 |
|
Musculus spp. |
+ |
30.0 |
8.3 |
|
Macoma balthica |
+ |
10.0 |
7.8 |
|
Mya arenaria |
+ |
2.5 |
0.5 |
|
Hiatella arctica |
— |
2.5 |
0.4 |
|
Cilietocardium ciliatum |
— |
2.5 |
0.1 |
|
Astartidae gen. sp. |
— |
2.5 |
1.5 |
|
Bivalvia indet. |
— |
2.5 |
2.2 |
|
Balanus sp. |
+ |
2.5 |
0.0 |
|
Gammarus sp. |
— |
7.5 |
1.4 |
|
Caprella spp. |
— |
5.0 |
2.4 |
|
Gammaridea gen. sp. |
+ |
12.5 |
2.7 |
|
Chironomidae gen. sp. |
+ |
2.5 |
0.1 |
|
Phyllophora |
— |
12.5 |
0.5 |
|
Laminaria ризоиды |
— |
5.0 |
0.1 |
|
Fucales |
— |
15.0 |
1.9 |
|
Нитчатые водоросли |
+ |
20.0 |
1.1 |
|
Empetrum nigrum |
— |
2.5 |
2.5 |
|
Растительные остатки |
+ |
17.5 |
1.9 |
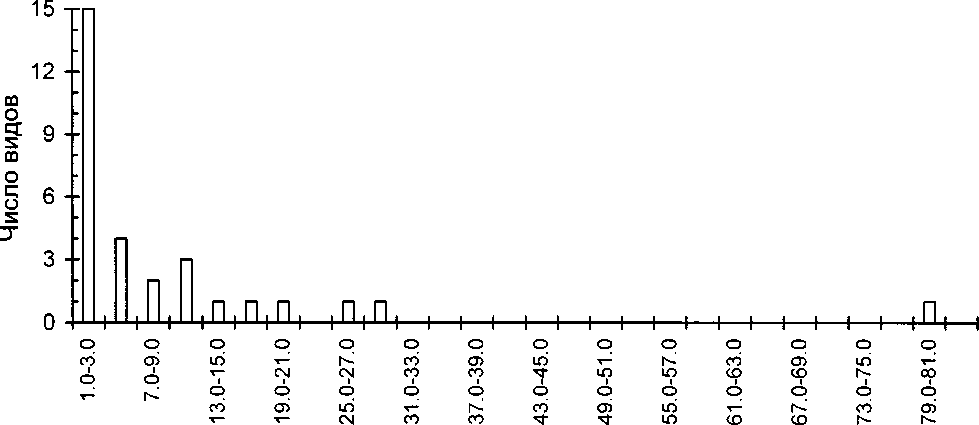
Суммируя полученные данные по питанию насиживающих самок и линяющих селезней видно, что главными объектами питания гоголей здесь на море являются мидии и бокоплавы, важными объектами служат мелкая рыба и икра рыб, комарики-звонцы, мускулюсы и макомы. В пресных водоёмах самки используют в первую очередь личинок ручейников, имаго жуков, а также, вероятно, рыб и их икру, бокоплавов. Их рацион здесь сходен с таковым на других водоёмах Северной Фенноскандии (Семёнов-Тян-Шанский, Гилязов 1991; Бианки 1999). Этот набор форм можно считать наиболее характерным составляющим рациона гоголя в Кандалакшском заливе.
При сравнении рациона гоголей, добытых в Кандалакшских шхерах и за их пределами (табл. 5) видно, что большее разнообразие условий в аква-
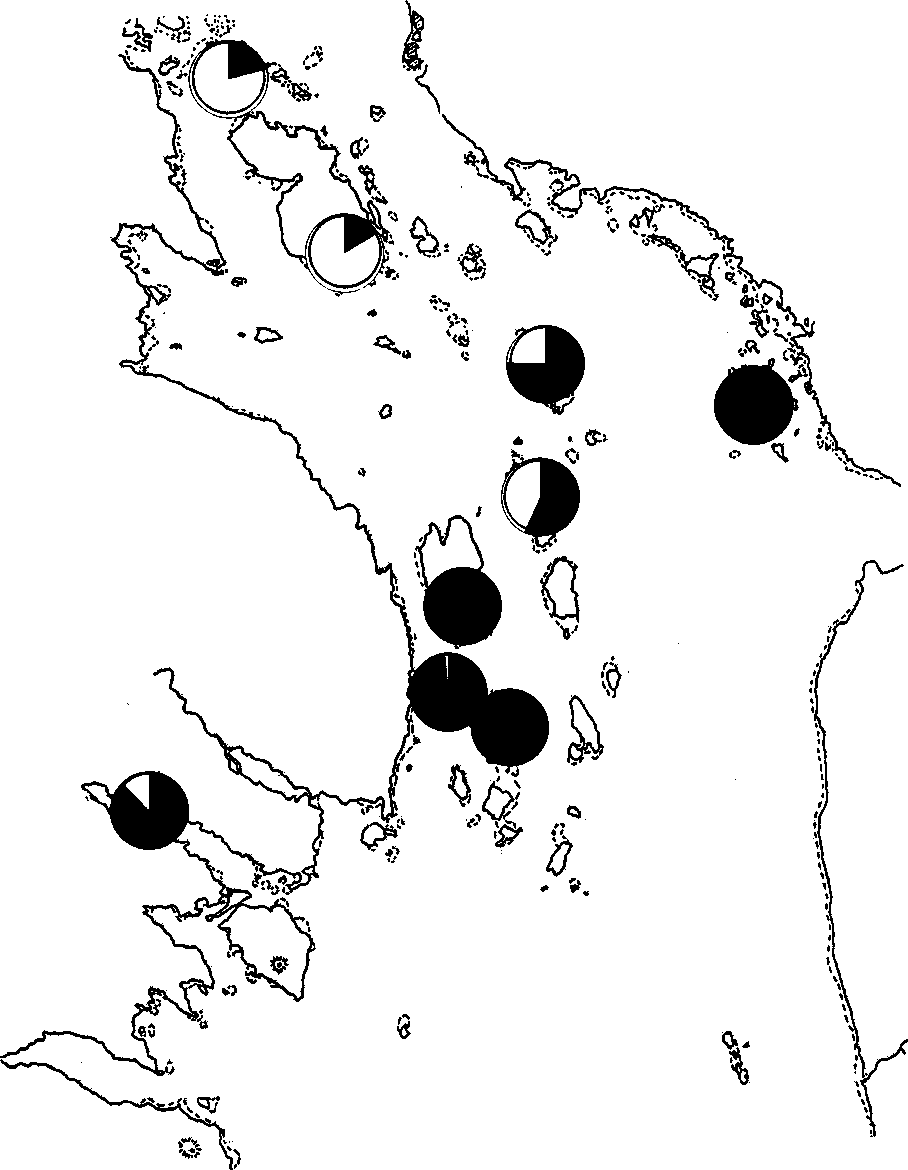
Рис. 4. Соотношение морских (чёрная заливка) и пресноводных (белая заливка) видов корма на разных островах в помёте гнездящихся самок гоголя.
В рационах линяющих селезней и насиживающих самок заметны различия. Из наиболее часто поедаемых объектов общими в них были только мидии и бокоплавы. Это объясняется, по-видимому, временем сбора материала и спецификой мест кормёжки птиц. Самки кормятся преимущественно около нижней границы литорали, самцы — в сублиторали. Материал по питанию самцов получен в годы, когда заросли взморника морского Zostera marina в Кандалакшском заливе ещё не восстановились после гибели в 1950-х годах, и подход к берегам трёхиглых колюшек Gasterosteus aculeatus на икрометание был незначителен.
Таблица 6. Многолетняя динамика средней доли (%) морских и пресноводных объектов в питании гоголей на острове Лодейном
|
Годы |
Морские виды |
Пресноводные виды |
|
1995-1996 |
28.9 |
34.4 |
|
1997-1998 |
16.7 |
0.0 |
|
1999-2000 |
49.1 |
1.0 |
|
2001-2002 |
31.6 |
41.8 |
Из таблицы 3 видно, что в помёте гнездящихся самок встречаются объекты пресноводного, морского и даже наземного происхождения. На рисунке 4 приведено соотношение средней доли морских и пресноводных объектов у самок, гнездящихся в разных местах. На островах Лодейном, Оленьем и Телячьем, где есть подходящие для кормёжки озёра, гоголи используют обитающих в них насекомых, ракообразных и рыб, хотя могут вылетать кормиться и на море. Их рацион включает как морские, так и пресноводные организмы. В целом по нашему материалу средняя доля объектов заведомо морского происхождения составляет 25.9%, пресноводного — 18.9%. Встречаемость пресноводных компонентов (36.3%) также несколько ниже, чем морских (50.4%).
Более детальный анализ многолетнего материала с о-ва Лодейного, где находится несколько озёр, показывает, что предпочтение морских и пресноводных объектов менялось*. С 1995 по 1997 доля пресноводных объектов в питании насиживающих самок снижалась, затем в течение 4 лет оставалась низкой, а в 2001-2002 снова увеличилась (табл. 6). Доля морских кормов изменялась противоположно. К сожалению, ряд объектов, которые часто встречаются в помёте гоголей на Лодейном, мы не можем с точностью идентифицировать как морских или пресноводных. К их числу относятся, например, бокоплавы и рыбы, которые обитают и в море, и в озёрах. Поэтому при анализе многолетнего изменения средней доли использовали только личинок ручейников, как типичных пресноводных объектов, и мидий, как типичных морских объектов. Полученный график (рис. 5) по- казывает, что с 1995 по 1996 гнездящиеся на Лодейном самки питались преимущественно на озёрах, где они и гнездились, а в 1997-2000 в значительной мере переключились на морские корма. В 2001-2002 произошло возвращение к питанию на озёрах. По всей видимости, это связано с нахождением в 1998-2000 годах на озёрах Лодейного американской норки Mustela vison, которая там дважды выращивала потомство.
Особо надо сказать о рационе гоголей, гнездящихся на о-ве Анисимове. Несмотря на отсутствие на нём озера, доля пресноводных видов в их питании довольно велика. Ближайшие же озёра находятся на о-ве Лодейном, куда, вероятно, самки летали кормиться, хотя там могла быть значительная конкуренция с местными птицами. Расстояние между этими островами — около 2 км, и некоторые самки гнездятся то на одном острове, то на другом.
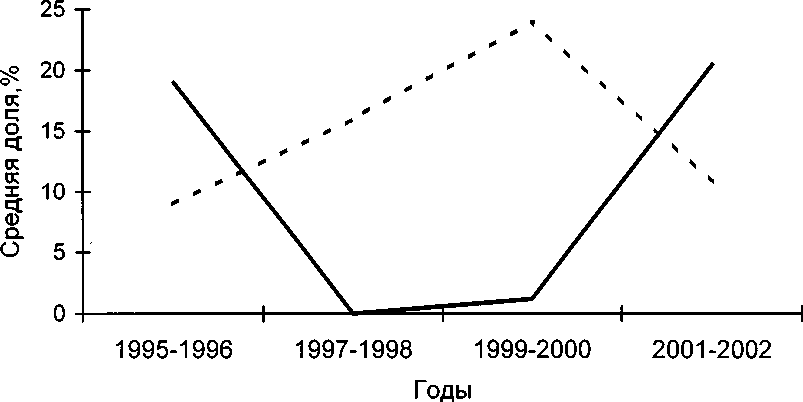
Рис. 5. Многолетняя динамика средней доли мидий Mytilus edulis и личинок ручейников Trichoptera в помёте самок гоголя, собранном на о-ве Лодейный.
У некоторых особей можно проследить сохранение привычных мест и (или) объектов кормёжки в течение нескольких лет. Например, экскременты утки (С-459707) гнездившейся в 1998-2001 на Лодейном в гнездовье № 8, на 80-100% состояли из остатков личинок звонцов, но в 2002 году их состав резко изменился, и они состояли из остатков Sigara sp. У другой самки (ES-006859), гнездившейся на том же острове в гнездовье № 37, в 1998-2000 экскременты на 100% состояли из остатков Mytilus edulis, в 2001 — из личинок подёнок (99%), а в 2002 — личинок ручейников (70%) и имаго водных жуков (30%). Последнее также подтверждает, что при наличии на озёрах норки самки вынужденно кормились на море, а с её уходом снова стали кормиться на озёрах.
Анализ полученных материалов по питанию насиживающих самок показывает, что состав его зависит от места гнездования и, соответственно, от места питания. Самки, гнездящиеся на островах, где нет пресных водоёмов, питаются на море у берега невдалеке от гнезда. Состав их рациона сходен с питанием гоголей, которые держатся на акватории Кандалакшских шхер (табл. 5). Важное значение в питании приобретают бокоплавы и рыба. Кроме того, в их питании появляется литоральный объект — личинки звонцов Chironomidae, и нередко встречаются гидробии, которых не было в рационе линяющих самцов. В одной пробе находилось очень много остатков от водяных клопов Sigara sp.
Самым частым морским компонентом оказывается мидия. Аналогов такому объекту в пресных водах очень мало, и они не играют существенной роли в питании гоголей. Мидия — моллюск неподвижный, стало быть нападение на него отличается от атаки на подвижный объект. Однако, это противоречие кажущееся.
Итак, значительная часть пищевых объектов гоголя на море представлена обитателями зарослей нитчатых водорослей, которые растут на небольшой глубине и колеблются под действием течений и волн. Этот “движущийся объект” со съедобными животными и привлекает гоголей.
