Плейотропное действие генов: факты, заблуждения и перспективы оценки

Автор: Шендаков А.И.
Журнал: Биология в сельском хозяйстве @biology-in-agriculture
Рубрика: Общие и частные вопросы генетики
Статья в выпуске: 1 (22), 2019 года.
Бесплатный доступ
Современная генетика представлена многими научными направлениями, многие направления генетики выделились в самостоятельные отрасли, такие, как цитогенетика, иммуногенетика, генетика растений, генетика животных, генетика человека, медицинская генетика и пр. Научные методы, применяемые в генетике, постоянно совершенствуются, однако целый ряд научных проблем, затрагиваемых генетикой, остаётся нерешённым. Отдельные фундаментальные проблемы одинаково важны для решения как в растениеводстве, так и ряде других отраслей. Одним из таких вопросов является вопрос определения плейотропного действия генов, осмысления его сущности в аспектах эволюции и видообразования. В научной работе сделан обзор современных данных о плейотропном действии генов, приведены частные случаи плейотропии у гороха, пшеницы, грибов, животных, человека и пр. Работа построена на обобщении данных, мнений и выявлении многообразия классификаций плейотропного действия генов; в работе приводятся случаи простой, истинной, косвенной, антагонистической и мультифакторной плейотропии. Отмечено, что плейотропия является побочным действием эволюции, однако для процесса селекции плейотропия крайне важна при её грамотном и целенаправленном использовании. Процесс проявления плейотропии усиливают мутации как у растений, так и у животных. Автором научной работы приводится собственная классификация разнообразия плейотропии, обозначаются факторы, способные затруднить результативность выявления плейотропного действия генов, а также обозначены перспективы дальнейших фундаментальных и прикладных исследований плейотропного действия генов. Автор статьи подчёркивает тот факт, что многофакторная (мультипликативная) плейотропия является естественным проявлением процесса организации и функционирования генома.
Генетика, плейотропия, гены, аллели, признаки, антагонистическая плейотропия, видообразование, мутации, эволюция
Короткий адрес: https://sciup.org/147230986
IDR: 147230986
Pleiotropic action of genes: facts, mistakes and prospects of evaluation
The present-day genetics is represented by many research areas, and many areas of genetics have separated into independent fields such as cytogenetics, immunogenetics, plant genetics, animal genetics, human genetics, medical genetics, etc. The scientific methods used in genetics are constantly being improved, but a number of scientific problems that are being solved in genetics still remain unresolved. It is equally important for both crop science and a number of other areas to have individual fundamental problems solved. One of the issues is the issue of determining the pleiotropic action of genes and understanding its essence in the aspects of evolution and speciation. The research reviews modern scientific data on the pleiotropic action of genes, presents individual cases of pleiotropy in peas, wheat, mushrooms, animals, humans, etc. The study is based on the generalization of data, opinions and the identification of a variety of classifications of the pleiotropic action of genes; the paper cites cases of simple, true, indirect, antagonistic and multifactorial pleiotropy. Pleiotropy has been noted to be a side effect of evolution, but pleiotropy is extremely important for the selection process if it is used in a competent and purposeful manner. The process of manifestation of pleiotropy increases mutations in both plants and animals. The author of the research provides his own classification of the diversity of pleiotropy, identifies factors that can hamper the effectiveness of detecting the pleiotropic action of genes and outlines the prospects for further fundamental and applied research on the pleiotropic action of genes. The author of the article emphasizes the fact that multifactorial (multiplicative) pleiotropy is a natural manifestation of the organization and functioning of the genome.
Текст научной статьи Плейотропное действие генов: факты, заблуждения и перспективы оценки
Плейотропи́я (от греч. πλείων – «больше» и греч. τρέπειν – «поворачивать, превращать») – это явление множественного действия, выражающегося в способности одного гена влиять на несколько фено типических признаков [44]. Наряду с эпистазом, сверхдоминированием и прочими явлениями наследственных факторов плейотропия представляет собой парадоксальное генетическое явление с точки зрения эволюции. При этом одновременное влияние одного гена нередко прослеживается не только сразу на два признака, а на три и большее количество признаков. Явление, характеризующее плейотропное действие генов, впервые было упомянуто ещё Г. Менделем, обнаружившим на растениях с пурпурными цветами красные пятна в пазухах листьев, а получаемые от таких растений семена всегда были покрыты серой или бурой кожурой. Г. Мендель в связи с этим предположил существование генетических факторов, детерминирующих данные признаки.
В настоящее время, с точки зрения научной проблемы, изучение плейотропии происходит комплексно, в нескольких направлениях, из которых можно выделить как целый ряд фундаментальных вопросов, так и огромное количество исследований случаев частной плейотропии: в растениеводстве и животноводстве, генетике человека и медицинской генетике в частности, в ветеринарии и защите растений, в изучении микроорганизмов, насекомых, грибов, водорослей, мхов, лишайников и пр. Учитывая то обстоятельство, что, по последним научным данным, в геноме млекопитающих присутствуют гены, характерные для других видов, микроорганизмов, червей и даже растений, изучение плейотропии как общебиологического явления представляется чрезвычайно актуальным.
Так, на сегодняшний день в периодической научной литературе приводятся новые данные о плейотропном действии генов у гороха [26, 33], пшеницы [8, 25], тыквы [2], редиса [18], грибов-паразитов [22], насекомых [13], в том числе жуков [17], крупных животных, в том числе птиц и овец [1], кроликов [3], лошадей [16], крупного рогатого скота [4, 37] и пр. Особое внимание к плейотропному действию в медицинской генетике обусловлено изучением предрасположенности к многофакторным заболеваниям [7, 9], в том числе к раковым заболеваниям, заболеваниям внутренних органов, сердечно-сосудистой системы и пр.
С фундаментальной точки зрения, обсуждаются вопросы сущности и причин возникновения плейо-тропного действия генов [11, 14, 15, 24, 36, 38], взаимосвязи проявления плейотропного действия генов с мутациями и общего влияния на эволюцию видов [42, 43]. При этом среди учёных с мировым именем до сих пор вызывают интерес вопросы исследования так называемой антагонистической плейотропии [28, 33], теория которой впервые была озвучена в 1957 году Williams G.C. [43]. К тому же, в настоящее время сформулирован целый ряд подходов при исследовании плейотропии, а сама классификация разновидностей плейотропного действия генов нуждается в систематизации и оценке перспектив её исследования в ближайшей перспективе, что, в целом, и послужило обоснованием актуальности данного обзора.
Простая и мультифакторная плейотропия. В растениеводстве и животноводстве, между тем, существуют разные подходы при интерпретации полученных научных данных: если в растениеводстве нередко основой для выведения нового сорта достаточно нескольких выявленных аллелей, ассоциированных с каким-либо ценным признаком, то в современном животноводстве селекционеры, как правило, имеют дело с количественными признаками (удой, живая масса, количество поросят и пр.), хотя селекция по масти, комолости, извитости шерсти или её отсутствию затруднений уже не вызывает. В равной степени селекционеры сталкиваются с затруднениями при необходимости постоянного увеличения урожайности и улучшения ряда количественных признаков у культурных растений.
При анализе генотипической (в том числе, генетической) изменчивости количественных признаков точность оценки плейотропного действия генов нередко зависит от правильности интерпретации полученных данных и генетико-статистических методов анализа. Проявление положительных или отрицательных генетических корреляций между признаками и вовсе до сих пор имеет два разных обоснования, одно из которых – плейотропия, другое – аддитивное (суммирующее) действие генов [1]. В селекции сельскохозяйственных животных нередко возникают противоречия при анализе генетических корреляций [19, 31, 34, 39], поскольку нельзя с полной долей уверенности при изучении признаков, детерминированных сотнями и, возможно, тысячами генов, однозначно говорить о плейотропии, аддитивном действии генов [20], сцепленном наследовании и пр. явлениях [23]. Однако, предпосылки к математическому выражению доли генов, вызывающих плейотропное действие, существуют, о чём следует говорить поэтапно.
Нередко доминантный ген, детерминирующий один признак, может оказаться рецессивным к другому признаку. Так, у крупного рогатого скота и кур известна коротконогость, вызываемая гетерозиготой, в рецессивном гомозиготном состоянии ген ведёт к летальному исходу. В птицеводстве приводятся примеры влияния генов, детерминирующих форму листовидного гребня, на яйценоскость. В труде «Генетика животных» Ф. Хатт [10] приводит пример мутации, при которой ген одновременно ведёт к зональному отсутствию шерсти на туловище и сосочков на языке у телят. В самом простом виде подобную плей-отропию схематически можно изобразить следующим образом (см. рис. 1).
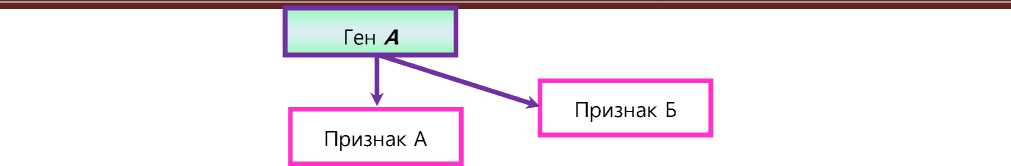
Рис. 1 – Детерминация одним геном двух признаков: простоя плейотропия
Исходя из рисунка, понятно, что у гена А есть основная функция и дополнительная. Всё было бы достаточно просто, если бы в этой ситуации не возникал вопрос, почему и на каком этапе эволюции гены стали выполнять дополнительные функции, не характерные для них, и зачем генам нужно выполнять «дополнительные обязательства». С точки зрения отдельных направлений генетики, плейотропия - побочное явление эволюции [11]. Вопрос определения временных рамок начала развития плейотропного действия генов в живых системах может оказаться нерешаемым, как и вопрос, например, определения точных временных рамок образования половой хромосомы Y.
Между тем, отдельные учёные (Бакай А. В. с соавторами, 2007), утверждает, что все гены в той или иной степени проявляют плейотропный эффект, в результате чего есть основания полагать, что любой наследственный признак определяется многими гена- ми, а точнее, всем генотипом [1]. К сожалению, авторы не уточняют, качественные или количественные признаки определяются всем генотипом, но логика такого суждения понятна, т. е. взаимосвязь генов и признаков приобретает сложнейшую матричную основу, подобную неким нейронным сетям или структурам, которые распутать невозможно априори.
Так или иначе, в уже опубликованных статьях автора данной работы ( А. И. Шендаков и др., 20122017 [12, 40, 41]) вполне определённо высказана теория мультифакторной плейотропии, сделаны попытки математического вычисления комплексного плейотропного эффекта генов через коэффициенты множественных генетических корреляций и даже доли плейотропных генов от общего числа генов, кодирующих тот или иной признак. Схема такой плей-отропии в самом простом варианте может выглядеть следующим образом (см. рис. 2).
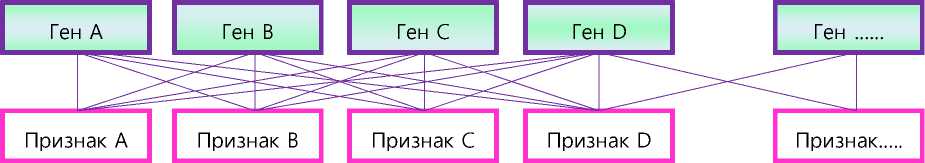
Рис. 2 – Один из вариантов множественной, мультифакторной плейотропии
В решении вопросов множественной, мульти-факторной плейотропии не следует также забывать, что, скорее всего, она может проявляться в большей степени по родственным селекционным признакам. В литературе приводится мнение о плейотропном действии генов на живую массу скота в 6, 10, 12 и 18 месяцев, однако здесь можно говорить ещё и том, что к формированию живой массы скота в 10 месяцев и старше могут подключаться новые кластеры генов, связанные с половым созреванием, развитием внутренних органов и т. п. То же самое можно сказать и об интенсивности роста плодов, их созревании и увядании.
Частные случаи плейотропии. Рассматривая частные случаи оценки плейотропии, следует отметить некоторые работы по растениеводству, животноводству, медицинской генетике и пр. Так, по данным Burstin J. с соавторами (2007), повышенная питательная ценность семян гороха (Pisum sativum) и особенно содержание белка в семенах при сохранении урожайности является важной задачей для дальнейшего развития этой культуры. Являясь сложными и нестабильными показателями, содержание белка и выход белка семян интегрируют все процессы, происходящие в течение жизненного цикла. Семена являются основным поглотителем азота, их потребность в азоте частично удовлетворяется не только корнями, но и азотом, который был получен из вегетативных органов. В восьми генотипах различного происхождения установлено, что как максимальная скорость накопления азота в семенах, так и мощность источника азота варьировали среди генотипов. В большинстве случаев гены Le и Afila, которые контролируют длину междоузлия и преобразование листочков в усики, соответственно, определяют состояние азота в растении. В зависимости от окружающей среды, эти гены были связаны с накоплением семенного белка [26]. (Более подробно с характеристикой этих и других мутаций у гороха можно ознакомиться в работе Костерина О.Э.(2015), опубликованной в обзорной статье в «Вавиловском журнале генетики и селекции» [6]).
Используя серию почти изогенных мутантных гороховых линий, дефектных в биосинтезе крахмала, Hughes R. K.и др. (2001) установили, что гены, влияющие на биосинтез крахмала, оказывали плейотроп-ное воздействие на содержание белка и состав семян гороха [32]. Китайские учёные ( Jing Wu и др, 2011 ) отмечают наличие у пшеницы плейотропных генов, ведущих к карликовости растения [25].
Бухаров А.Ф. с соавторами (2016) отмечает, что наличие высоких значений коэффициентов корреляции между парами признаков «гидроксикоричные кислоты» - «флавоноиды» (в мякоти плодов и в семенах), а также среднего значения коэффициента корреляции признаков «каротин в плодах» - «каротин в семенах», по-видимому, связано с общностью путей их биосинтеза и, возможно, обусловлено не только явлением сцепления, но и явлением плейотропии [2].
Поползухиной Н.А. и Кротовой Л.А. (2009) отмечен плейотропный характер мутантных признаков, когда наряду с основным признаком изменяется целый комплекс других признаков, а на проявление мутантных признаков оказывают влияние условия выращивания и генотипическая среда. При помещении мутантного гена в новую генотипическую среду происходит существенное варьирование всего комплекса признаков [8].
Международный научный коллектив, состоящий из французских, испанских и американских учёных (G. Pierre и др.,2014), отмечает [22], что простейшие грибы, паразитирующие на яблонях, адаптируются к хозяину и могут полноценно функционировать только через общего хозяина после получения ресурсов. Поскольку один ген токсина или аллель вирулентности может дать возможность колонизировать хозяина, адаптация и репродуктивная изоляция ( reproductive isolation ) мгновенно упрощаются, что, в свою очередь, плейотропно вызывает адаптивное видообразование. Исследования по эволюционной геномике грибов дополнительно объясняют самые ранние стадии расхождения в результате потока генов и дают представление о плейотропно индуцированной адаптивной дивергенции у других эукариот.
Amanda S. Lobell, Rachel R. Kaspari, Yazmin L. Serrano Negron and Susan T. Harbison (2017) в статье «The Genetic Architecture of Ovariole Number in Drosophila melanogaster: Genes with Major, Quantitative, and Pleiotropic Effects» отмечают, что плейотропные гены, детерминирующие процессы размножения у дрозофил, могут быть как фенотипически проявившимися, так и «спящими», к плейотропным генам были отнесены гены, содержащие один или несколько полиморфных вариантов, значительно связанных с числом овариолей ( яйцевых трубок в яичнике ), и один или несколько вариантов, значительно связанных с одним «спящим» признаком [13].
По данным Bellone R.R.(2010), охарактеризовано одиннадцать различных генов, которые способствуют вариациям масти у лошадей. К сожалению, мутации, связанные с пигментацией, часто приводят к пагубным последствиям в других системах. В частности, обнаружены шесть плейотропных эффектов, ассоциированных с пигментными генами. К ним относятся неврологические дефекты (летальный синдром белых жеребят и лавандовый синдром – Lаvender Fоаl Syndrоme, LES ), нарушения слуха, нарушения зрения (врожденная ночная слепота и множественные врожденные глазные аномалии), а также специфическая для лошади меланома [16].
Плейотропия играет большую роль в генетике человека и медицинской генетике в частности. Как отмечают Слепцов А.А., Назаренко М.С. (2016), большое количество проведенных полногеномных ассоциативных исследований (GWAS) указывает на широкое распространение плейотропии. Молекулярные меха- низмы плейотропии не только помогают объяснить взаимосвязь между заболеваниями, но также позволяют по-новому взглянуть на патогенез конкретных заболеваний, что приводит к улучшению их профилактики, диагностики и лечения. Однако, как отмечают авторы, большинство плейотропных эффектов остаются неопределёнными, в связи с тем, что их функциональная значимость не была систематически оценена. Такое исследование требует больших усилий и уникального подхода, основанного на последовательном анализе нескольких уровней биологических процессов (транскрипция, трансляция). Появление крупных массивов информации в медицине, таких, как мультиомиксные данные, лабораторно инструментальная визуализация патологических состояний и ведение электронных баз данных историй болезней предоставляет большие возможности для исследования плейотропии [9].
Мнения и подходы при исследовании плей-отропии. В мировой практике, как было уже отмечено, учёные высказывают разные мнения о плейотроп-ном действии генов и строят разные теории о плей-отропии. Еще в 1982 году в труде «Антагонистическая плейотропия, доминирование и генетическая вариация» известный канадский учёный Michael A. Rose высказал мнение, что, по всей вероятности, специфические закономерности действия генов и, следовательно, обоснованность конкретных генетических моделей может быть затруднительна для экспериментального анализа - по чисто практическим соображениям. Тем не менее, возможности для взаимодействия между теорией и экспериментом в популяционной генетике существуют [29].
Annalise B. Paaby, Matthew V. Rockman (2013) [14] отмечают, что плейотропия может возникать из нескольких различных, но потенциально перекрывающихся механизмов, таких, как: 1) плейотропия гена, 2) плейотропия развития и 3) селекционная плейотро-пия. Генная плейотропия возникает, когда продукты деятельности гена взаимодействует с несколькими другими белками или катализирует множественные реакции. Развивающая плейотропия возникает в тех случаях, когда мутации оказывают множественное воздействие на полученный фенотип. Селекционная плейотропия возникает тогда, когда полученный фенотип оказывает большое влияние на выживаемость (в зависимости от таких факторов, как возраст и пол).
В научной работе о функциональных компромиссах генов и оценке плейотропии («Gene Functional Trade-Offs and the Evolution of Pleiotropy») Frédéric Guillaume, Sarah P. Otto (2012) пишут [21], что плей-отропия является свойством генов, затрагивающих множество функций или характеристик. Гены сильно различаются по степени плейотропии, но эта вариация часто считается побочным продуктом их эволюции. Гены, которые вносят вклад в две функции, вносят вклад в одну функцию, уменьшая действие на вторую функцию. Также гены становятся плейотроп-ными или специализируются на одной функции в зависимости от характера так называемых «компромиссов». В целом, когда генный продукт может хорошо функционировать при двух функциях, он развивается. Если же плейотропия сильно нарушает его основную функцию, то она не развивается в последующих поколениях. Следовательно, ослабевающий плейотроп-ный эффект чаще развивается у генов, детерминирующих признак, который в настоящее время наиболее важен. Даже если плейотропия закрепляется и развивается в популяции, то не все гены, как ожидается, становятся равноценно плейотропными. Экспрессия генов способствует плейотропии.
Австралийские учёные Katrina McGuigan, Locke Rowe, Mark W. Blows в работе « Pleiotropy, apparent stabilizing selection and uncovering fitness optima » (2011) [27] отмечают, что, согласно эволюционной теории, эволюцию отдельных признаков нельзя понять изолированно, когда присутствует плейотропия. Широко распространенная плейотропия вызывает проявление стабилизирующего отбора по признакам.
В отзыве «Об определении и измерении плей-отропии» (« On the definition and measurement of pleiot-ropy ») Jianzhi Zhang и Gunter P. Wagner (2013) [24] отмечают, что концепция плейотропии, несмотря на 103-летнюю историю, имеет фундаментальную важность в генетике развития, эволюции и медицинской генетике, однако и по сегодняшний день эмпирических знаний о распространённости и механизмах плейотропии недостаточно. Обсуждая некоторые другие теории (в частности, «плейотропию гена», «плей-отропию развития» и «селекционную плейотропию» Annalise B. Paaby and Matthew V. Rockman [14]), авторы отмечают, что нет необходимости в данных теориях выделять доминантную и рецессивную плейотро-пию, а плейотропия – это, в целом, не свойство гена, а свойство мутации. Плейотропия летальных и (или) стерильных мутаций не является характеристикой эволюционирующей способности генотипа, поскольку эти мутации никогда не будут конкурировать за реализацию в следующем поколении. Уровни плейо-тропных эффектов летальных аллелей малы. Дальнейший продуктивный путь – это расширение и улучшение попыток оценки плейотропии эмпирически, в больших масштабах и для многих видов.
Между тем, Sarah P. Otto в работе « Два шага вперед, один шаг назад: плейотропные эффекты предпочтительных аллелей » (2004) [36] отмечает, что мутантные аллели, которые являются полезными, закрепляются в популяции, а средний селективный эффект мутантных аллелей, которые являются полезными, фиксируется, в том числе, на одну треть плей-отропией. Эти результаты помогают сформировать понимание эволюционной инерции, вызванной плей-отропией.
В актуальной современной публикации «Pleiot-ropy in complex traits: challenges and strategies» Nadia Solovieff, Chris Cotsapas, Phil H. Lee, Shaun M. Purcell, Jordan W. Smoller (2013) [30] делают обзор проблем и стратегий изучения плейотропии. В частности, авторы подчёркивают, что в настоящее время в исследованиях ассоциаций генома выявлено множество вариантов, каждый из которых влияет на комплексные признаки, особенно при аутоиммунных заболеваниях, раковых и нейропсихиатрических заболеваниях человека. Эти результаты дают возможность сделать вывод, что плейотропные эффекты на комплексные черты могут быть широко распространены. Однако си- стематическое обнаружение таких эффектов является сложной задачей и требует новых методологий и основ для интерпретации результатов. Авторы доказывают наличие плейотропии в современных исследованиях генетического картирования, предлагают новые аналитические подходы к выявлению плейотропных эффектов, выявляют источники ложных кросс-фенотипных эффектов, а также дают соображения для будущих направлений исследований плейотропии. Авторы выделяют три разновидности биологической плейотропии: опосредованную плейотропию, ложную (по причине неправильной интерпретации или погрешности вычисления) и ложную «подвижную» плейотропию, когда признаки детерминируются разными генами. При этом, в их понимании, опосредованная плейотропия является реальной формой плей-отропии, в отличие от ложной плейотропии, но суть её состоит в том, что она не прямо влияет на признак, с косвенно, т. е. детерминирует какой-либо иной признак, вызывающий развитие основного, анализируемого признака, в отношении которого исследователи подразумевают истинную плейотропию.
В 2011 году Sandra J. Hasstedt и Alun Thomas с помощью пакета Java Pedigree Analysis Package в множественных локусах количественных признаков выявили не только плейотропные эффекты, но и эпистаз. Так, предполагаемая плейотропия, вызывающая ожирение у человека, детерминирована шестью локусами (на хромосомах 1, 2, 13, 16, 20 и 22) [35].
Существуют вызывающие доверие современные примеры выявления плейотропных эффектов у крупного рогатого скота. Так, результаты работы Schrooten C. (2004), опубликованной в монографии в виде тезисов докторской диссертации “Genomic variation in dairy cattle – Identification and Use» , показали, что процентное содержание жира и белка в молоке коров кодируют гены, расположенные на 6 хромосоме, однако есть плейотропные гены, отвечающие за эти признаки (на 6 и 14 хромосомах), одновременное увеличение % белка и молочного белка в кг (на 6 хромосоме), одновременное увеличение % жира в молоке и количества молочного белка (на 23 хромосоме) и пр.[37].
Вопросы, связанные с теорией антагонистической плейотропии. Эта теория нуждается в отдельном пояснении. Впервые она была предложена Williams G.C. в работе «Pleiotropy, natural selection, and the evolution of senescence», которая была опубликована в 1957 году [42]. Данная теория, объясняющая процесс старения, нашла отражение во многих современных трудах по теории плейотропии и экспериментальном анализе данного генетического явления. Согласно Leroi A.M., Barke A., De Benedictics G., Francecshi C., Gartner A., Feder M.E., Kivisild T., Lee S., Kartal-Ozer N., et al. (2005), теория антагонистической плейотропии объясняет, почему размножение может иметь негативный эффект на долговечность вида и может даже вызвать смерть, как это происходит у одноплодных организмов. Авторы объясняют плейотропное действие генов на процесс старения через накапливающиеся мутации. Так, мутации, которые вызывают перепроизводство половых гормонов, увеличивают тенденцию к усиленному размножению и успех при размножении. В связи с этим мутантные формы сохраняются, несмотря, например, на то, что у таких организмов могут возникать заболевания половых органов на поздних этапах онтогенеза. Потому идея решения возможного компромисса в пользу размножения непосредственно вытекает из теории антагонистической плейотропии [28]. Несмотря на то, что конкретные плейотропные гены долгое время не были идентифицированы в силу отсутствия необходимых методов и знаний, впоследствии теория антагонистической плейотропии была подтверждена в экспериментах с использованием мушки дрозофилы (Drosophila melanogaster) - классического объекта генетических исследований. В исследованиях на дрозофиле учёным удалось увеличить продолжительность жизни за счёт ограничения размножения до позднего возраста. В подтверждение состоятельности результатов наблюдения, авторы привели тот факт, что среди мух-долгожительниц (с помощью селекции и закреплённых мутаций) наблюдалось ослабленная плодовитость.
Аналогичный эффект был обнаружен и среди круглых червей (Caenorhabditis elegans) , где был идентифицирован ряд долгоживущих мутантов, которые отличались пониженной плодовитостью и уступали нормальным представителям популяции во время циклов голодания. В целом, данный результат является спорным относительно интерпретации на млекопитающих и крупных животных. На сегодняшний день исчерпывающих данных об антагонистической плейотропии нет.
Профессор университета в штате Мичиган Roger L. Albin в журнале « Genetica » (1993), отмечает, что теория антагонистической плейотропии является наиболее убедительной теорией для обоснования процесса старения, связанного, в том числе, с накоплением мутаций, а генетические заболевания дают возможность выявить потенциально плейотропные аллели и локусы [33].
Американский профессор из Флориды Thomas F. Hansen (2003) в публикации «Is modularity necessary for evolvability? Remarks on the relationship between pleiotropy and evolvability» строит теорию о том, что увеличение количества плейотропных генов способствует усилению эволюции, а то обстоятельство, при котором все гены были бы плейотропными, привело бы к максимальной эволюции [42]. Автор подтверждает мнение о том, что плейотропно действующие аллели могут иметь как положительный, и так отрицательный плейотропный эффект на другие признаки. Гены с дифференцированными типами плейотропных эффектов могут компенсировать ограничения друг друга. В отличие от теории «скрытого плейотропного эффекта», где положительная и антагонистическая плейотропия исключаются, автор утверждает, что чередующиеся плейотропные эффекты в популяции в целом равны нулю, а наиболее развитой генетической «архитектурой» является та, где гены могут влиять на разные характеристики (признаки), однако различными путями. Универсальная генетическая структура проявляет максимально изменяющиеся плейотропные эффекты. Также автор отмечает, что к плейотропному действию гена может привести любая потенциальная мутация.
Чтобы лучше понять процессы, связанные с плейотропией, Жданова О.Л. и Фрисман Е.Я. (2014) [5] разработали и исследовали математическую модель динамики генетической структуры и численности популяции с двумя возрастными классами, в которой выживаемости зигот и половозрелых особей определялись одним плейотропным локусом. Даже при таких ограничениях, когда внешние проявления сложной полигенной системы были сведены к моно-локусному случаю, рассматриваемая модель демонстрировала широкий спектр различных эволюционных сценариев изменения динамики популяции. Увеличение репродуктивного потенциала и выживаемости сопровождалось переходом от стационарной динамики численности популяции к колебаниям. Однако эволюционный рост самих этих параметров мог быть немонотонным, со значительными флуктуациями; причем в случае антагонистической плейотропии рост одного из этих параметров, как правило, сопровождался закономерным уменьшением другого, что могло привести даже к стабилизации численности и генетического состава возрастных классов. Авторами было показано, что действие отбора на более поздней стадии жизненного цикла сопровождалось нарушением равенств Харди-Вайнберга, связывающих частоты аллелей и генотипов. Было получено балансовое соотношение, позволяющее сравнивать совокупные приспособленности генотипов, и показано, что отбор идет в сторону вытеснения менее приспособленных генотипов. При этом начальные условия в отдельных случаях могли определять как генетический состав популяции, так и характер динамики её численности.
Обобщения, выводы и предложения по дальнейшим исследованиям. Обобщая результаты анализа встречающихся в настоящее время данных о плейотропии, можно резюмировать и подвести некоторый промежуточный итог для систематизации разновидности этого сложного генетического явления.
Несмотря на многообразие данных и большое количество противоречивых мнений об этом генетическом явлении, в общих чертах плейотропию можно систематизировать по продолжительности действия, по биологической значимости, сложности и направленности действия на признаки (качества, черты), характеристики и какие-либо качества исследуемых объектов, включая растения, простейшие организмы, животных, человека и пр. (см. рис. 3).
Основным противоречием в данном случае можно считать тот факт, относить ли к истинной (генетической) плейотропии косвенную (опосредованную) плейотропию и где находятся пределы так называемой «истинности» и «косвенности» плейотропного действия генов. Скорее всего, отвечая на этот вопрос, следует исходить из того, какое количество генов и аллелей ассоциировано с тем или иным признаком. Вполне возможно, что правомочно называть плей-отропию истинной только при анализе качественных признаков, где генетическая составляющая детерминации взаимосвязанных признаков неоднократно доказана эмпирически и подтверждена разными авторитетными научными коллективами. При этом следует подчеркнуть, что определение истинного плейотроп-ного влияния генов будет затрудняться: 1) с увеличением количества генов и аллелей, детерминирующих интересующий признак, процесс, качество и пр.; 2) при наличии других генетических явлений, по своему действию способных интерпретировать их в качестве плейотропии, в т.ч. при сцепленном наследовании генов; 3) при существенном влиянии средовых факторов на проявление анализируемых признаков и взаимосвязи «генотип-среда; 4) в случае активного
возникновения мутаций в процессе жизнедеятельности организма, выращивания, возделывания и эволюции вида; 5) в связи с наличием истинной мультифак-торной и косвенной мультифакторной плейотропии; 6) по причине переменности плейотропного действия генов, ощутимой динамике генетической изменчивости и общей мобильности генома; 7) в связи с вирусными заболеваниями, воздействующими на геном.
Разновидности плейотропии

по продолжительности действия
Список литературы Плейотропное действие генов: факты, заблуждения и перспективы оценки
- Бакай А.В., Кочиш И. И., Скрипниченко Г. Г. Генетика. - М.: КолосС, 2007. - 448 с.
- Бухаров А.Ф., Степанюк Н.В., Бухарова А.Р. Корреляции биохимических показателей антиоксидантной активности и хозяйственно ценных признаков тыквы крупноплодной // Вестник Российского государственного аграрного заочного университета. - 2016. - № 21 (26). - С. 9-14.
- Глазко В.И., Косовский Г.Ю., Глазко Т.Т. Доместикация как генетический феномен // Кролиководство и звероводство. - 2018. - № 1. - С. 5-8.
- Епишко Т.И., Яцына О.А., Смунева В.К., Яцына В.В. Плейотропное действие гена каппа-казеина (CSN3) на воспроизводительные качества быков-производителей //Ученые записки учреждения образования Витебская ордена Знак почета государственная академия ветеринарной медицины. - 2010. - Т. 46. №1-2. - С. 21-24.
- Жданова О.Л., Фрисман Е.Я. Моделирование отбора по плейотропному локусу в двухвозрастной популяции // Генетика. - 2014. - Т. 50. № 8. - С. 996.
