Плейотропные ферменты апоптоза и синаптическая пластичность гиппокампа белых крыс после окклюзии общих сонных артерий

Автор: Авдеев Д.Б., Акулинин В.А., Степанов А.С., Горбунова А.В., Степанов С.С.
Журнал: Сибирский журнал клинической и экспериментальной медицины @cardiotomsk
Рубрика: Экспериментальные исследования
Статья в выпуске: 3 т.33, 2018 года.
Бесплатный доступ
Цель: исследование было посвящено изучению плейотропных свойств фермента апоптоза каспазы-3, его связи с синаптической пластичностью гиппокампа белых крыс в норме и после 20-минутной окклюзии общих сонных артерий.Материал и методы. С помощью световой (окраска гематоксилином и эозином), электронной (уранилацетат и цитрат свинца) микроскопии, иммуногистохимии (MAP2, синаптофизин, каспаза-3, p53, bcl-2) и морфометрии были изучены общая численная плотность нейронов, ультраструктура синапсов, площадь иммунопозитивных синаптических терминалей stratum radiatum СА1 и stratum lucidum СА3 гиппокампа головного мозга в норме (n=5) и через 1 (n=5), 3 (n=5), 7 (n=5), 14 (n=5) и 30 суток (n=5) после острой ишемии.Результаты и обсуждение. Исследование показало, что в течение 30 суток постишемического периода 33,0% пирамидных нейронов СА1 и 17,4% СА3 подвергалось необратимой деструкции. Среди необратимо поврежденных преобладали нейроны с признаками коагуляционно-ишемического некроза. На этом фоне сначала (1 сутки) уменьшалась, а затем (3, 7 суток) восстанавливалась относительная площадь синаптофизин-позитивного материала. Установлено, что локализация каспазы-3 совпадала с локализацией синаптофизина. Особенно наглядно это было показано на гигантских синапсах stratum lucidum поля СА3. В телах пирамидных нейронов гиппокампа каспаза-3 не выявлялась, она локализовалась только в терминалях аксодендритических, аксошипиковых и аксосоматических синапсов. В динамике постишемического периода наиболее выраженные изменения содержания каспазы-3 отмечались в stratum radiatum поля СА1. Белки регуляции апоптоза (р53, bcl-2) выявлялись в единичных нейронах. В этой связи каспазу-3 необходимо рассматривать в аспекте ее плейотропности, участия в адаптационных и восстановительных процессах за счет постишемической активации нейропластичности на уровне аксонов и синапсов.Заключение. После острой ишемии, вызванной 20-минутной окклюзией общих сонных артерий, активация каспазы-3 способствует защите нервной ткани.
Острая ишемия, гиппокамп, каспаза-3, р53, синаптофизин, мар-2
Короткий адрес: https://sciup.org/149125225
IDR: 149125225 | DOI: 10.29001/2073-8552-2018-33-3-102-110
Pleiotropic enzymes of apoptosis and synaptic plasticity in albino rat hippocampus after occlusion of common carotid arteries
Aim: the aim of the study was to investigate the pleiotropic properties of the apoptotic enzyme caspase-3 and its associations with the synaptic plasticity of the hippocampus of albino rats in healthy animals and in rats after 20-min occlusion of the common carotid arteries.Material and Methods. Total numerical density of neurons, ultrastructure of synapses, and area of immunohistochemically positive hippocampal synaptic terminals of CA1 stratum radiatum and stratum lucidum CA3 were studied by the methods of optical microscopy (hematoxylin and eosin stain), electron microscopy (uranyl acetate and lead citrate as contrast agents), immunohistochemistry (MAP2, synaptophysin, caspase-3, p53, and bcl-2), and morphometry in the brains of intact rats (n=5) and in animals after acute ischemia at day 1 (n=5), 3 (n=5), 7 (n=5), 14 (n=5), and 30 (n=25).Results and Discussion. The study showed that 33.0% of pyramidal neurons in CA1 region and 17.4% of those in CA3 region underwent irreversible damage within 30 days of the post-ischemic period. Among the irreversibly damaged neurons, the cells with signs of coagulative-ischemic necrosis prevailed. In animals subject to ischemia, the relative area of synaptophysin-positive material initially decreased (at day 1) and then recovered (at days 3, 7). We found that caspase-3 colocalized with synaptophysin, which was especially evident in the giant synapses of the stratum lucidum of the hippocampal CA3 region. In the neurosomes of the hippocampal pyramidal cells, caspase-3 was not detected. However, this enzyme was found in the terminals of the axo-dendritic, axo-spine, and axo-somatic synapses. In the course of th e post-ischemic period, the most pronounced changes in the expression of caspase-3 were observed in the stratum radiatum of the CA1 field. Apoptosis regulatory proteins (p53, bcl-2) were detected in the individual neurons. In this regard, caspase-3 should be viewed in the context of its pleiotropy and involvement in the adaptation and recovery processes due to post-ischemic activation of neuroplasticity at the level of axons and synapses.Conclusion. After acute ischemia caused by 20-min occlusion of the common carotid arteries, the activation of caspase-3 contributes to ischemic preconditioning and neuroprotection.
Текст научной статьи Плейотропные ферменты апоптоза и синаптическая пластичность гиппокампа белых крыс после окклюзии общих сонных артерий
Природа постишемической гибели пирамидных нейронов гиппокампа белых крыс после транзиторной ишемии переднего мозга изучена довольно полно. Установлено, что после ишемии в гиппокампе активировались механизмы некроза и апоптоза [1–3]. При развитии апоптоза показана роль каспазы-3. Так, имеются данные о том, что клетки с апоптоз-подобной морфологией и/или экспрессией каспазы-3 составляют менее 10% ишемически измененных нейронов, в то время как подавляющее большинство — это нейроны, погибающие путем некроза. Смерть СА1 пирамидных клеток после транзиторной ишемии переднего мозга инициируется через активацию c-JunN-terminal kinase (JNK). Затем следует апоптоз или вторичный (отдаленный) некроз [4].
Ключевыми структурами, связанными с активацией механизмов некроза и апоптоза нервной ткани головного мозга млекопитающих, являются межнейронные синапсы. Это обусловлено тем, что возбуждающие нейромедиаторы после повреждения синаптических пузырьков и выхода в межклеточное пространство запускают кальций-зависимые эксайтотоксические механизмы гибели нейронов [5, 6]. Изменения синапсов гиппокампа происходят быстро в ранние сроки после ишемии и отражают соответствующие патофизиологические нарушения в синаптической передаче. В любом случае, чем активнее выделение глутамата, тем выше вероятность активации кальций-зависимых механизмов деструкции постсинаптических нейронов [7, 8].
Кроме того, синапсы гиппокампа обладают высокой пластичностью, что обеспечивает восстановление и перестройку межнейронных отношений этого отдела головного мозга после ишемии [9]. Последнее также отмечено другими авторами [10]. Максимальные проявления диффузно-очагового разрушения синаптических контактов отмечались через 1–3 суток после острой ишемии, а восстановление межнейронных связей происходило на фоне исчезновения признаков отека-набухания нервной ткани через 7–14 суток [9, 11].
В последнее время активно изучаются неапоптотиче-ские свойства протеолитических ферментов, в частности каспаз. Каспаза-3 рассматривается как один из ключевых адаптивных ферментов [12, 13]. Каспаза-3 может быть активирована в результате Ca2+-зависимого процесса, при этом активация проходит по неизвестному механизму, а активированный фермент обладает меньшей, чем при апоптозе, активностью [14–16].
Кроме того, апоптотические протеиназы имеют функции, непосредственно связанные с работой нервной системы в норме, иными словами, с нормальной нейропластичностью. Например, пластичность и подвижность конусов роста сопровождается локальной активацией каспазы-3 в конусе роста. Ингибирование каспазы-3 в зрелом головном мозге приводит к нарушению пластических процессов и снижению адаптивных возможностей мозга. То есть каспазе-3 присуща плейотропность или мультимодальность, что реализуется в вовлеченности этого фермента во множество различных функций, часто прямо противоположных. При развитии церебральных патологий этот фермент опосредует как гибель нервных клеток, так и компенсаторные процессы, необходимые для выживания нейронов и нормального функционирования мозга в целом. Именно поэтому блокирование каспазы-3 в мозге для лечения церебральных патологий не дало положительных результатов и даже опасно для его восстановления. Кроме того, после ишемии нервной ткани происходит адаптивная активация каспазы-3 без каких-либо признаков апоптоза [12, 13].
Таким образом, все вышесказанное свидетельствует от тесной связи механизмов развития апоптоза и нейропластичности. При этом исход постишемического периода зависит от характера и степени тяжести острой ишемии [12, 13].
Обзор литературы показал, что практически нет работ, посвященных сопоставлению иммуногистохимических проявлений активации синаптической пластичности и уровня экспрессии каспазы-3 после 20-минутной окклюзии общих сонных артерий.
Цель настоящего исследования: иммуногистохимическое изучение коры гиппокампа для выявления связи синаптической пластичности и активности каспазы-3 как одного из плейотропных ферментов защиты головного мозга.
Материал и методы
Исследование выполнено на базе ФГБОУ ВО «Омский государственный медицинский университет» Минздрава России, одобрено этическим комитетом университета (протокол № 83 от 14 октября 2016 г.).
В качестве экспериментальных животных использовали белых крыс ( n =30, самцы) линии Wistar массой 180–200 г. Животные содержались в обычном виварии. Исследования проводили в соответствии с «Правилами проведения работ с использованием экспериментальных животных» (Приложение к приказу Министерства здравоохранения СССР от 12.08.77 № 755), рекомендациями Международного комитета по науке о лабораторных животных, поддержанными ВОЗ, директивой Европейского Парламента № 2010/63/EU от 22.09.10 «О защите животных, используемых для научных целей».
Применен комплексный методологический подход, включающий экспериментальный, анатомический, гистологический, морфометрический и информационноматематический методы, а также методы наблюдения, описания и анализа.
Выход из эксперимента осуществляли путем декапитации в соответствии с директивой Европейского Парламента № 2010/63/EU от 22.09.2010 «О защите животных, используемых для научных целей».
Острую 20-минутную ишемию мозга путем пережатия общих сонных артерий (2-сосудистая модель неполной глобальной ишемии без гипотонии) моделировали на фоне премедикации (сульфат атропина 0,1 мг/кг, подкожно) и общей анестезии (Zoletil 100, 10 мг/кг).
Общее состояние животных в послеоперационном периоде оценивалось по 100-бальной системе [17] с учетом принципов исследования физиологии и патофизиологии головного мозга [18, 19].
Взятие материала проводили через 1 ( n =5), 3 ( n =5), 7 ( n =5), 14 ( n =5) и 30 ( n =5) суток после ишемии. Контролем ( n =5) служили ложнооперированные (без окклюзии артерий) животные того же возраста. Головной мозг фиксировали путем перфузии 4%-го раствора параформа на 0,1 М фосфатном буфере (рН — 7,4) через восходящую часть дуги аорты. Серийные фронтальные срезы (2–4 мкм) на уровне гиппокампа [20] окрашивали гематоксилином и эозином, а также с помощью иммуногистохимического метода против каспазы-3, р-53, bcl-2, синаптофизина (р38) и MAP-2 (производитель: Leica Biosystems Newcastle Ltd, Великобритания). Окраска осуществлялась согласно рекомендациям фирмы производителя реагентов. На микроскопе Leica DM 1000 делались цифровые микрофотографии серийных срезов (по 200 полей зрения с каждого отдела на срок).
Для исследования ультраструктуры клеток, отростков и синапсов материал контрастировали 1– 2 ч в 1%-м неза-буференном растворе четырехокиси осмия, заключали в смесь эпона и аралдита. Ультратонкие срезы окрашивали уранилацетатом и цитратом свинца. Использовали ультрамикротом LKB- 8800 (Швеция), микроскоп Hitachi-600H (Япония). Фотографировали на пленки 9 x 12 см при разном увеличении.
Морфометрический анализ проводили с использованием ранее разработанного подхода к оценке систем межнейронной коммуникации с помощью иммуногистохимических методов и программы ImageJ 1.46 [21]. Определяли общую численную плотность нейронов, относительную площадь р38- и каспаза-позитивного материала. Проверку статистических гипотез осуществляли непараметрическими критериями Манна — Уитни, ANOVA (однофакторный дисперсионный анализ Краскела — Уоллиса) и критерия х 2 (StatSoft STATIS-TICA 8.0) [22]. Результаты представлены как медиана (нижний, верхний квартили) и в процентах (95%-й доверительный интервал — 95% ДИ). В ходе проведения статистического анализа нулевая гипотеза отвергалась при p ≤ 0,05.
Результаты
Для молекулярного слоя поля СА3 гиппокампа характерно наличие уникальной зоны — stratum lucidum ), которая локализуется на уровне апикальных участков дендритов пирамидных нейронов и состоит из дендритов, гигантских синапсов, сформированных аксонами мшистых волокон, и небольшого количества отростков астроцитов. Существенно то, что при окраске гематоксилином и эозином stratum lucidum не контрастировалась с другими уровнями молекулярного слоя СА3 и выглядела как в СА1 (рис. 1 а, б). Совершенно иная картина наблюдалась при иммуногистохимическом выявлении цито-скелетного белка MAP2 (microtubule-associated protein 2; dendrite-specific). Stratum lucidum была представлена в виде светлой полосы, содержащей срезы дендритов с MAP2-позитивным материалом, над слоем тел пирамидных нейронов только в поле СА3. Светлые участки соответствовали синаптическим терминалям, цитоскелет которых не содержит MAP2-позитивный материал (рис. 1 в, г; 3 а, б).
Именно эти синаптические терминали (светлые участки при окраске на МАР2) в stratum radiatum СА1 и особенно гигантские терминали в stratum lucidum СА3 интенсивно окрашивались при иммуногистохимическом выявлении синаптофизина (рис. 2 а, б). В stratum radiatum СА1 крупные терминали практически не выявлялись. Локализация меток была вдоль дендритов и в нейропиле (рис. 2а). В stratum lucidum СА3 метки локализовались вдоль апикальных дендритов (рис. 2б).
Существенно то, что при окраске на каспазу-3 локализация меток (рис. 2 в, г) соответствовала таковой при окраске на синаптофизин (рис. 2 а, б). В телах пирамидных нейронов каспаза-3 не выявлялась, она локализова- лась только в терминалях аксодендритических, аксоши-пиковых и аксосоматических синапсов (рис. 2 в, г).
Таким образом, по своей локализации каспаза-3 соответствовала стратегическим зонам реализации механизмов синаптической пластичности. При этом большее ее количество было в терминалях аксошипиковых и аксо-дендритических синапсов. Это позволяет предположить, что данный плейотропный фермент апоптоза может принимать участие в механизмах защиты, компенсации и восстановления межнейронных связей гиппокампа как в норме, так и после ишемического воздействия. При этом острая ишемия головного мозга может стимулировать защитные свойства каспазы-3.
Для проверки этой гипотезы в настоящей работе оценивали изменения количества нейронов, ультраструктуры синапсов, сравнивали площадь меток при иммуногистохимическом выявлении синаптофизина и каспазы-3 в терминалях. Изучение апоптоза сводилось к определению активности про- (р53), антиапоптозных (bcl-2) белков и каспазы-3 в перикарионе нейронов.
Установлено, что в постишемическом периоде происходило уменьшение общей численной плотности пирамидных нейронов. Через 30 суток после окклюзии дефицит нейронов в поле СА1 составил 33,0%, а в СА3 — 17,4% (табл. 1).
В остром периоде (1, 3 суток) после острой ишемии в гиппокампе появлялись нейроны с признаками тин-кториальных (усиление и уменьшение интенсивности окраски цитоплазмы и ядра — гиперхромные и гипохромные), гидропических (вакуолизация, отек-набухание), дегидратационных (темные клетки с обратимым сморщиванием) и некробиотических (клетки-тени, пик-номорфные, гомогенизированные клетки) изменений,
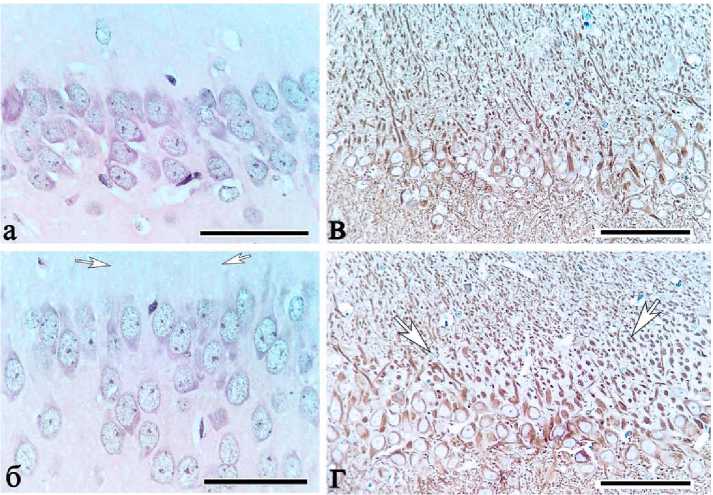
Рис. 1. Поля СА1 (а, в) и СА3 (б, г) гиппокампа контрольных белых крыс: демонстрация выявления и локализации stratum lucidum (стрелки); а, б — окраска гематоксилином и эозином, в, г — иммуногистохимическая окраска против MAP2. Докраска гематоксилином. Объектив х 40, шкала — 100 мкм (в, г); х 100, шкала — 25 мкм (а, б)
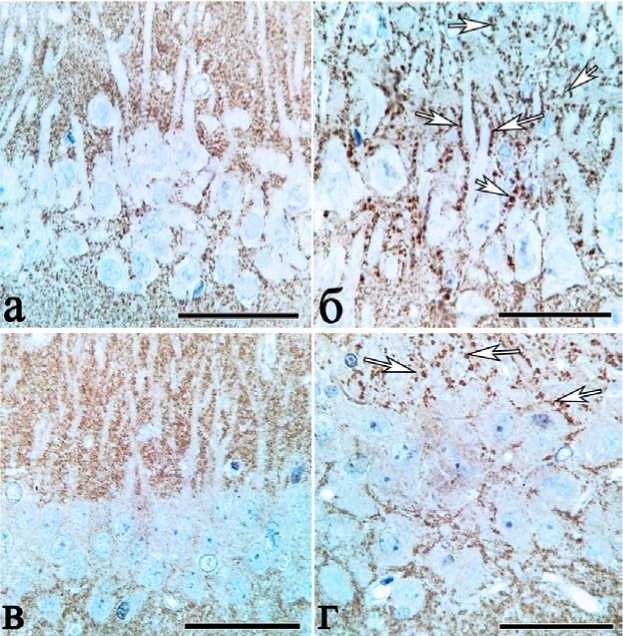
Рис. 2. Поля СА1 (а, в) и СА3 (б, г) гиппокампа контрольных белых крыс: наличие гигантских терминалей в stratum lucidum (стрелки) СА3 и более мелких терминалей в stratum radiatum СА1 при иммуногистохимической окраске против синаптофизина (а, б) и каспазы-3 (в, г). Докраска гематоксилином. Объектив х 100, шкала — 25 мкм
Таблица 1
Общая численная плотность (на 1 мм2площади клеточных слоев) нейронов гиппокампа в постишемическом периоде, Me (QL-QU)
|
Группы животных |
Поля гиппокампа |
|
|
СА1 |
СА3 |
|
|
Ложнооперированные |
2936 (2726-3652) |
1983 (1755-2106) |
|
1 сутки |
2322 (2167–2553)* |
1754 (1541–1902)* |
|
3 суток |
2308 (1960–2456)* |
1699 (1432–1712)*^ |
|
7 суток |
2135 (1865–2301)*^ |
1650 (1415–1720)* |
|
14 суток |
2090 (1767–2115)*^ |
1662 (1398–1734)* |
|
30 суток |
1983 (1764–2213)* |
1638 (1178–1702)* |
|
ANOVA |
df=5; Н=18,5; p =0,02& |
df=5; Н=16,6; p =0,03& |
Примечание: * — сравнение с ложнооперированными животными, ^ — сравнение с предыдущим сроком (критерий Манна — Уитни), & - сравнение между всеми сроками по данному полю гиппокампа (ANOVA); различия статистически значимы при p<0,05. Анализировали по 5 – 10 серийных срезов с каждого животного. Me — медиана, QL- нижний , QU — верхний квартили, ANOVA — однофакторный дисперсионный анализ Краскела — Уоллиса.
фагоцитоза и мелкие поля «выпадения» нейронов. Среди необратимо поврежденных преобладали пикноморфные нейроны с признаками коагуляционно-ишемического некроза (рис. 3 а, б). В молекулярном слое появлялись контуры окрашенных эозином апикальных дендритов (рис. 3 б). Нейроглиальные взаимоотношения характеризовались увеличением количества единичных, групп астроцитов и микроглиоцитов около патологически измененных нейронов.
Через 7 суток в гиппокампе сохранялись проявления кариоцитолизиса, отека-набухания и вакуолизации тел нейронов, пикноморфные нейроны. Выявлялись очаги выпадения нейронов и большое количество микроглиоцитов, которые располагались около ишемически измененных нейронов либо отдельно, либо в составе группы с астроцитами.
Через 14 суток рядом с очагами ишемически поврежденных нейронов часто выявлялись гипертрофи-
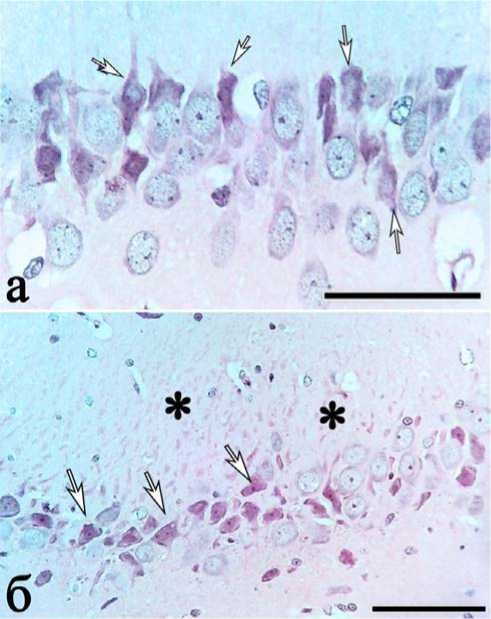
Рис. 3. Поля СА1 (а) и СА3 (б) гиппокампа через 1 сутки после ишемии: преобладают нейроны с признаками коагуляционноишемического некроза (стрелки), в stratum lucidum (*) видны контуры апикальных дендритов. Окраска гематоксилином и эозином. Объектив х 100, шкала — 25 мкм (а); х 40, шкала — 100 мкм (б)
рованные нейроны. Даже через 30 суток после ишемии сохранялись очаги пикноморфных нейронов с большим содержанием глиальных клеток.
Изучение ультраструктуры синапсов гиппокампа показало, что после ишемии появлялось большое количество синаптических терминалей, измененных по светлому типу деструкции. Подобные изменения затрагивали все типы синапсов, включая и гигантские в stratum lucidum (рис. 4б).
Таким образом, после 20-минутной окклюзии общих сонных артерий в гиппокампе происходила необратимая деструкция части пирамидных нейронов и разрушение межнейронных синапсов.
По данным иммуногистохимического исследования, белки регуляции апоптоза (р53, bcl-2) выявлялись в единичных нейронах, основная масса перикарионов была bcl-2-, р53-негативной. Каспаза-3 имела высокую активность только в аксонах и синаптических терминалях, а в перикарионах меток к данному белку, как и в контроле, не выявлялось. Мы полагаем, что это свидетельствовало о том, что при использованной модели острой ишемии не выявлялось функционально значимых объемов элиминации нейронов путем апоптоза. Единичные нейроны с признаками активации белков регуляции апоптоза без наличия активной каспазы-3 в цитоплазме сомы нейронов свидетельствовали только о принципиальной возможности реализации данного процесса. Каспазу-3 в данном случае необходимо рассматривать в аспекте ее плейотропности, участия в адаптационных и восстановительных процессах за счет постишемической активации нейропластичности на уровне аксонов и синапсов.
Для этого в полях СА1 и СА3 проведено сравнение относительной площади меток синаптофизина и каспазы-3 в динамике постишемического периода. Установлено, что в поле СА1 количество меченного синаптофизина снижалось через 1 и 3 суток. Затем этот показатель восстанавливался до контрольного уровня (табл. 2).
В гигантских синапсах поля СА4 содержание синап-тофизина уменьшалось через 1 сутки после ишемии, а уже через 3 суток было на уровне контроля (табл. 3).
Все это свидетельствовало о том, что выявленное нами разрушение терминалей по светлому типу сопровождалось, вероятно, и разрушением синаптофизина, связанного с синаптическими пузырьками во всех синапсах.
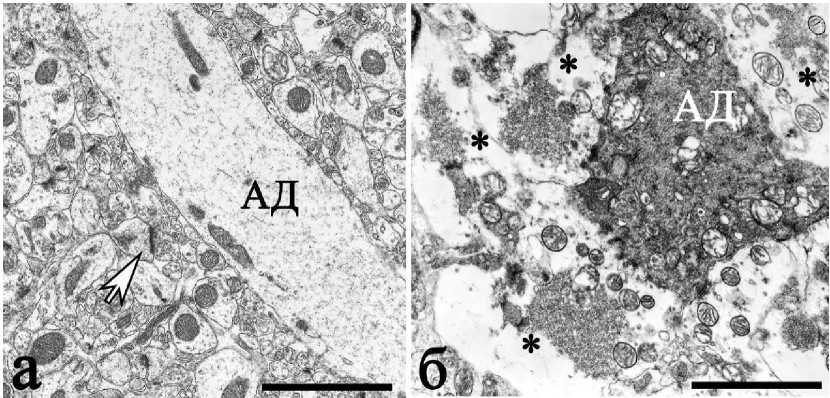
Рис. 4. Ультраструктура зоны апикальных дендритов (АД) поля СА1 (а — норма) и СА3 (б — через 1 сутки после ишемии): вокруг дендритов в СА1 преобладают мелкие и средние синапсы (стрелка), для поля СА3 после ишемии характерно наличие гигантских терминалей с признаками светлого типа деструкции (*). Окраска уранилацетатом и цитратом свинца, шкала — 2 мкм
Таблица 2
Относительная площадь (%) меток синаптофизина и каспазы-3 в stratum radiatum поля СА1 гиппокампа белых крыс в норме и после восстановления кровотока (по 200 полей зрения при каждом сравнении на срок)
|
Группы |
Белки |
|
|
Синаптофизин |
Каспаза-3 |
|
|
Ложнооперированные |
20,2 (14,9–26,4) |
25,4 (19,5–32,0) |
|
1 сутки |
9,1 (5,5–14,0) |
13,6 (9,2–19,1) |
|
χ 2=8,9; p = 0,003 * |
χ 2=8,1; p=0,004 * |
|
|
3 суток |
11,4 (7,4–16,6) χ 2=5,2; p = 0,02 * |
22,1 (16,5–28,5) χ 2=4,4; p=0,037 ^ χ 2=7,5; p = 0,01# |
|
34,3 (27,8–41,3) |
||
|
7 суток |
16,7 (11,8–22,6) |
χ 2=6,8; p =0 ,01^ χ 215,4; p = 0,0001*# |
|
38,5,2 (31,7–45,6) |
||
|
14 суток |
12,8 (8,1–17,7) |
χ 2=7,3; p = 0,007* χ 2=33,2; p = 0,0001# |
|
30 суток |
17,5 (12,5–23,5) |
20,1(14,8–26,3) χ 2=15,5; p = 0,0001 ^ |
Примечание: различия статистически значимы в cравнении с контролем (*), с предыдущим сроком (^) и между белками (#) при p≤0,05 (критерий χ 2, n=200). Данные представлены как среднее и 95% ДИ.
Таблица 3
Относительная площадь (%) меток синаптофизина и каспазы-3 в stratum lucidum поля СА3 гиппокампа белых крыс в норме и после восстановления кровотока (по 200 полей зрения на каждый срок)
|
Группы |
Белки |
|
|
Синаптофизин |
Каспаза-3 |
|
|
Ложнооперированные |
24,6 (18,8–31,2) |
21,1 (15,7–27,4) |
|
1 сутки |
15,8 (11,0–21,6) χ 2=4,3; p=0,038 * |
19,5 (14,3–25,7) |
|
3 суток |
22,0 (16,5–28,4) |
27,8 (21,7–34,6) |
|
7 суток |
29,2 (23,0–36,0) |
23,3 (17,6–29,8) |
|
14 суток |
22,3 (16,7–28,7) |
19,7 (14,4–25,9) |
|
30 суток |
28,2 (22,1–35,0) |
17,6 (12,6–23,6) |
Примечание: * — в cравнении с контролем различия статистически значимы при p≤0,05 (критерий χ 2). Данные представлены как среднее и 95% ДИ.
В норме относительная площадь меток каспазы-3 в поле СА1 статистически значимо не отличалась от таковой синаптофизина (табл. 1 и 2). В постишемическом периоде наиболее выраженные изменения содержания каспазы-3 отмечались в stratum radiatum поля СА1. Здесь содержание каспазы-3 в остром периоде по сравнению с контролем сначала (1 сутки) уменьшалось, а через 3 суток восстанавливалось. Затем (7 и 14 суток) сохранялось высокое содержание этого белка. При этом через 3, 7 и 14 суток относительная площадь меток каспазвы-3 была больше, чем меток синаптофизина (табл. 1). В stratum lu- cidum поля СА3, содержащем большое количество гигантских терминалей, статистически значимых изменений содержания каспазы-3 выявить не удалось (табл. 2).
Обсуждение
Ранее при использовании 2-сосудистой модели неполной глобальной ишемии без гипотонии (ООСА) нами была показана смешанная (парапоптоз) природа гибели нейронов неокортекса — гидропическая дистрофия, сочетание процессов некроза (колликвационного и коагуляционного) и апоптоза. Однако основную роль в гибели нейронов все же играли процессы быстрого (аутолитического колликвационного) и отдаленного постишемического некроза (коагуляционного на фоне истощения лизосомальных ферментов) [9, 11]. По данным литературы, каспаза-3 рассматривается не только как ключевой фермент конечной стадии апоптоза, но и как важный химический компонент ишемического прекондиционирования и синаптической пластичности [12, 13]. В результате возникла необходимость выявления связи синаптической пластичности и активности каспазы-3 как одного из ключевых ферментов защиты нейронов.
Наиболее подходящей моделью острой ишемии для решения поставленных задач была 20-минутная окклюзия общих сонных артерий, после которой, как правило, возникают диффузно-очаговые изменения нервной ткани головного мозга экспериментальных животных [9, 11]. Выбор отдела головного мозга был обусловлен тем, что в stratum lucidum поля СА3 содержится большое количество гигантских синапсов, которые можно изучать с помощью методов световой иммуногистохимии.
При сравнении серийных фронтальных срезов гиппокампа мы показали, что места локализации синапто-физина в stratum lucidum совпадали с локализацией каспазы-3 — гигантские синаптические терминали. Метки обоих белков четко выявлялись в виде округлых гранул, локализацию и размеры которых можно было оценить с помощью программы ImageJ. Установлено, что максимальное содержание каспазы-3 было в поле СА1 через 3–14 суток после острой ишемии.
Таким образом, мы показали, что каспаза-3 в норме и после ишемии локализовалась в синаптических терминалях, а в перикарионах этот белок отсутствовал. Кроме того, с помощью иммуногистохимии нам не удалось показать активации белков р53 и bcl-2. То есть подтверждались неапоптотические свойства каспазы-3, которая, вероятно, участвовала в постишемической активации нейропластичности. По нашим данным, в постишемическом периоде на фоне активации синапсов и компенсаторного неосинаптогенеза происходила активация каспазы-3 в терминалях и аксонах без каких-либо признаков ее активации в перикарионе, свойственной для апоптоза.
Заключение
-
1. Плейотропный фермент апоптоза каспаза-3 в норме и после острой ишемии локализуется в аксонах и си-
- наптических терминалях, имеет связь с механизмами синаптической пластичности гиппокампа белых крыс.
-
2. Максимальная активация каспазы-3 происходит в stratum radiatum поля СА1 гиппокампа через 3–14 суток после восстановления кровотока.
Список литературы Плейотропные ферменты апоптоза и синаптическая пластичность гиппокампа белых крыс после окклюзии общих сонных артерий
- Zeng Y. S., Xu Z. C. Co-existence of necrosis and apoptosis in rat hippocampus following transient forebrain ischemia. Neurosci. Res. 2000; 37: 113-125. DOI: 10.1016/s0168-0102(00)00107-3
- Winkelmann E. R., Charcansky A., Faccioni-Heuser M. C., Netto C. A., Achaval M. An ultrastructural analysis of cellular death in the CA1 field in the rat hippocampus after transient forebrain ischemia followed by 2, 4 and 10 days of reperfusion. Anat. Embryol. 2006; 211: 423-434. DOI: 10.1007/s00429-006-0095-z
- Wirth C., Brandt U., Hunte C., Zickermann V. Structure and function of mitochondrial complex I. Biochim Biophys Acta. 2016; 1857(7): 902-914. DOI: 10.1016/j.bbabio.2016.02.013
- Muller G. J., Stadelmann C., Bastholm L., Elling F., Lassmann H., Johansen F. F. Ischemia leads to apoptosis-and necrosis-like neuron death in the ischemic rat hippocampus. Brain Pathol. 2004; 14(4): 415-424. DOI: 10.1111/j.1750-3639.2004.tb00085.x
- Baron J.-C., Yamauchi H., Fujioka M., Endres M. Selective neuronal loss in ischemic stroke and cerebrovascular disease. J. Cereb. Blood Flow Metab. 2014; 34: 2-18. DOI: 10.1038/jcbfm.2013.188
- Maurer L. L., Philbert M. A. The mechanisms of neurotoxicity and the selective vulnerability of nervous system sites. Handb. Clin. Neurol. 2015; 131: 61-70.
- DOI: 10.1016/B978-0-444-62627-1.00005-6
- Nikonenko A. G., Radenovic L., Andjus P. R., Skibo G. G. Structural features of ischemic damage in the hippocampus. The anatomical record. 2009; 292: 1914-1921.
- DOI: 10.1002/ar.20969
- Shetty A. K. Hippocampal injury-induced cognitive and mood dysfunction, altered neurogenesis, and epilepsy: can early neural stem cell grafting intervention provide protection? Epilepsy Behav. 2014; 38: 117-124.
- DOI: 10.1016/j.yebeh.2013.12.001
- Степанов А. С., Акулинин В. А., Степанов С. С., Авдеев Д. Б. Клеточные системы восстановления и утилизации поврежденных нейронов головного мозга белых крыс после 20-минутной окклюзии общих сонных артерий. Российский физиологический журнал им. И. М. Сеченова. 2017; 103(10): 1135-1147.
- Семченко В. В., Степанов С. С., Боголепов Н. Н. Синаптическая пластичность головного мозга (фундаментальные и прикладные аспекты). 2-е изд. М.: 2014: 408.
- Степанов А. С., Авдеев Д. Б., Акулинин В. А., Степанов С. С. Структурно-функциональные изменения нейронов неокортекса белых крыс после 20-минутной окклюзии общих сонных артерий. Патологическая физиология и экспериментальная терапия. 2018; 62(2): 30-38.
- Яковлев А. А., Гуляева Н. В. Плейотропные функции протеиназ мозга: методические подходы к исследованию и поиск субстратов каспазы. Биохимия. 2011; 76(10): 1325-1334.
- DOI: 10.1134/s0006297911100014
- Яковлев А. А., Гуляева Н. В. Прекондиционирование клеток мозга к патологическим воздействиям: вовлеченность протеаз (обзор). Биохимия. 2015; 80(2): 204-213.
- McLaughlin B., Hartnett K. A., Erhardt J. A., Legos J. J., White R. F., Barone F. C., Aizenman E. Caspase 3 activation is essential for neuroprotection in preconditioning. Proc. Natl. Acad. Sci. USA. 2003; 100: 715-720.
- DOI: 10.1073/pnas.0232966100
- Launay S., Hermine O., Fontenay M., Kroemer G., Solary E., Garrido C. Vital functions for lethal caspases. Oncogene. 2005; 24: 5137-5148.
- DOI: 10.1038/sj.onc.1208524
- Khalil H., Peltzer N., Walicki J., Yang J.-Y., Dubuis G., Gardiol N., Held W., Bigliardi P., Marsland B., Liaudet L., Widmann Ch. Caspase 3 protects stressed organs against cell death. Mol. Cell Biol. 2012; 32: 4523-4533.
- DOI: 10.1128/mcb.00774-12
- Корпачев В. Г., Лысенков С. П., Тель Л. З. Моделирование клинической смерти и постреанимационной болезни у крыс. Патологическая физиология и экспериментальная терапия. 1982; 3:78-80.
- Буреш Я., Бурешова О., Хьюстон Д. П. Методики и основные эксперименты по изучению мозга и поведения. М.: Высшая школа; 1991: 399.
- Васильев Ю. Г., Вольхин И. А., Данилова Т. Г., Берестов Д. С. Оценка неврологического статуса домашних и лабораторных животных. Международный вестник ветеринарии. 2013; 3: 52-55.
- Paxinos G., Watson C. The Rat Brain in Stereotaxic Coordinates. 5th ed. San Diego: Elsevier Academic Press; 2005: 367. 21. Акулинин В. А., Степанов С. С., Авдеев Д. Б., Степанов А. С., Разумовский В. С., Артюхов А. В., Горбунова А. В. Особенности изменений неокортекса, архикортекса и миндалевидного тела белых крыс после острой ишемии. Журнал анатомия и гистопатология. 2018; 7(2): 9-17.
- DOI: 10.18499/2225-7357-2018-7-2-9-17
- Боровиков В. Statistica. Искусство анализа данных на компьютере. 2-е изд. СПб: Питер; 2003: 688.
