Плейстоценовый морж на реке Печоре: минералого-геохимические данные и палеоэкологические реконструкции

Автор: Силаев В.И., Смолева И.В., Филиппов В.Н., Шанина С.Н., Хазов А.Ф., Макеев Б.А., Киселева Д.В., Фокина А.К., Йоган Ван дер П.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Научные статьи
Статья в выпуске: 2 (60), 2023 года.
Бесплатный доступ
Исследованы фрагменты левого бивня плейстоценового моржа, ископаемые остатки которого были обнаружены в 2009 г. на берегу р. Печоры. Проанализированы гранулометрический, химический и нормативно-минеральный составы вмещающих кости грунтов; термические свойства, химический и микроэлементный составы бивня; рентгеноструктурные параметры и химический составы костного биоапатита; макростроение, элементный и аминокислотный составы костного органического вещества; изотопный состав углерода, кислорода в биоапатите и углерода, азота - в костном коллагене. Для биоапатита установлен умеренно изотопно-легкий углерод, свойственный внепещерным ископаемым костям плейстоценовых животных, и изотопно-тяжелый кислород, типичный для гидрокарбоната морской воды. Изотопные данные для органического вещества печорского моржа коррелируются с аналогичными характеристиками морских животных, но при этом указывают не на моллюсковую диету, свойственную современным моржам, а на рыбную. Последнее свидетельствует о необычной для морских хищников среде обитания печорского моржа и необычном рационе его питания.
Морж, р. печора, бивень, минералого-геохимические и изотопно-геохимические свойства
Короткий адрес: https://sciup.org/149143575
IDR: 149143575 | УДК: 548+549+902+904 | DOI: 10.19110/1994-5655-2023-2-57-67
Pleistocene walrus on the Pechora river: mineralogical and geochemical data and paleoecological reconstructions
Left tusk’s fragments of the Pleistocene walrus were studied. Its fossil remains were found on the bank of the Pechora River in 2009. The analyses covered granulometric, chemical and normative-mineral composition of grounds inside the bones; thermal properties, chemical and microelemental composition of the tusk; X-ray diffraction parameters and chemical composition of bone bioapatite; macrostructure, elemental and amino acid composition of bone organic matter; isotopic composition of carbon, oxygen in bioapatite and carbon, nitrogen in bone collagen. Bioapatite was identified for moderately isotopically light carbon, characteristic of extracave fossil bones of the Pleistocene animals, and isotopically heavy oxygen, typical of seawater bicarbonate. The isotopic data for the organic matter of the Pechora walrus correlated with the similar characteristics of marine animals but simultaneously indicated not a mollusk diet, typical of modern walruses, but a fish diet. The latter fact evidenced the habitat and the diet of the Pechora walrus being untypical for marine predators.
Текст научной статьи Плейстоценовый морж на реке Печоре: минералого-геохимические данные и палеоэкологические реконструкции
В 2009 г. на бечевнике на правом берегу р. Печоры (N65°25.750’ E52°23.852’), примерно в 2 км выше дер. Гарево Усть-Цилемского района Республики Коми (рис. 1), местный житель А.А. Глухенький нашел крупный костный фрагмент, который он передал в Геологический музей им. А.А. Чернова
Института геологии ФИЦ Коми НЦ УрО РАН (инвентарный номер 548/317).
Обнаруженный фрагмент является частью рострального отдела черепа моржа Odobenus rosmarus Linnaeus, 1758 (рис. 2, а, б), включающей верхнечелюстные кости со
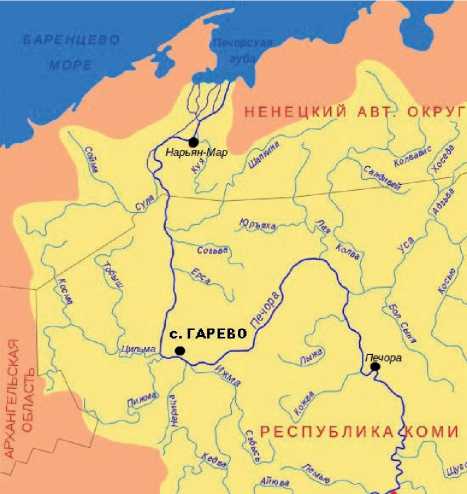
Рисунок. 1. Место находки костного фрагмента на реке Печоре.
Figure 1. Location area of the bone fragment’s finding on the Pechora River.
запредельный для 14С-метода возраст – более 45 тыс. лет.
Следует отметить, что это не единственная находка морского млекопитающего на р. Печоре. Б.И. Гуслицером в 1973 и 1981 гг. упоминались находки здесь в отложениях верхнего плейстоцена кости нерпы (определение И.Е. Кузьминой и Н.К. Верещагиной). Известен также случай захода в этот район белого медведя, сильно истощенную особь которого местные жители добыли в 1980-е гг. у дер. Мыла, расположенной в 65 км к западу от дер. Гарево.
В целом хорошая сохранность кости, отсутствие на ней следов окатанности и обработки человеком, а также то, что фрагмент черепа слишком крупный и тяжелый для дальнего переноса рекой к месту находки свидетельствуют, вероятно, о его местном захоронении. Исходя из имеющихся на настоящий момент геологических и геохронометрических данных, можно предположить, что появление моржа в районе широтного отрезка р. Печоры в 270 км по прямой от Печорского моря и в 400 км от устья р. Печоры произошло во времена или родионовского (стадия MIS 7, пик 243 тыс. л. н.), или микулинского (подстадия MIS 5e, пик 123 тыс. л. н.) интерстадиалов. На последний из них приходится сулинская бореальная трансгрессия.

Рисунок 2. Внешний вид современного моржа (а), лицевой отдел черепа найденного на р. Печоре ископаемого моржа атлантического подвида (б) и фрагменты левого бивня, послужившие объектом исследований (в).
Figure 2. The modern walrus specimen (a), the facial skull of fossil walrus of the Atlantic subspecies found on the Pechora River (б), and left tusk’s fragments under study (в).
Основная часть сводного разреза четвертичных отложений в районе находки костных остатков печорского моржа состоит из трех горизонтов морен: одного нижне-неоплейстоценового (?) и двух средненеоплейстоценовых – печорского (MIS 8) и вычегодского (MIS 6) с разделяющими их песчаными и глинистыми осадками, предположительно, родионовского возраста. Верхняя часть разреза представлена осадками, относи- скуловыми отростками, а также предчелюстные и носовые кости. Альвеола правого клыка (бивня) частично разрушена в дистальной части. Левый бивень (С1) обломан, оставшаяся часть коронки составляет, мм: по высоте – 20, по ширине – 37, по толщине – 52. Из постоянных зубов присутствуют по одному резцу (I3) и по три премоляра (Pm1– Pm3) с каждой стороны. Жевательные зубы характеризуются хорошо выраженными поверхностями стирания. По данным сравнительного морфологического исследования, проведенного Н.В. Крюковой (Институт проблем экологии и эволюции им. А.Н. Северцова РАН, г. Москва), поверхность стачивания функциональных зубов (I3 Pm1 Pm2 Pm3) и их высота над десной соответствуют типу моржей с традиционным питанием, где основной пищей является бентос. Сравнение найденного образца с черепами современных моржей известного возраста показывает, что находка отвечает половозрелому животному атлантического подвида в возрасте 13–14 лет.
В Центре изотопных исследований Гронингенского университета (г. Гронинген, Нидерланды) была сделана УМС радиоуглеродная датировка (GrA 66467), показавшая мыми к отложениям ранне-средневалдайского подпрудного озера и покровному комплексу, сформировавшемуся, скорее всего, во время последнего ледникового максимума. Хотя ближайшие разрезы с отложениями морской бореальной трансгрессии расположены на удалении 150– 200 км от места находки моржа, мы предполагаем, что эта находка связана именно с сулинской бореальной трансгрессией, случившейся 130–115 тыс. л. н. (рис. 3).
Во внутренней полости бивня ископаемого моржа был обнаружен терригенный материал, имеющий, вероятно, отношение к вмещающим ископаемого моржа грунтам. По гранулометрическому составу этот материал – гравийно-песчаный, несортированный, с практически полным набором песчаных фракций (табл. 1). В их составе рентгенофазовым анализом установлены кварц, альбит, мусковитоподобная слюда, хлориты, кальцит и апатит (рис. 4). Последний, судя по уширенности соответствующих рентгеновских пиков, обусловлен мелким детритом костного материала самого печорского моржа. По пропорциям между нормативными содержаниями кварца+альбита, слюды и хлоритов грунты отвечают хлорит-слюдистым супесям,
|
Средний плейстоцен |
Верхний плейстоцен |
Рисунок 3. Положение костных остатков печорского моржа на шкале времени AICC2012 – показано точкой в рамке интервала микулинского межледниковья и сулинской бореальной морской трансгрессии.
Figure 3. The position of the Pechora walrus bone remains on the AICC2012 time scale. It is marked by a dot in the interval frame of the Mikulin interglacial and the Sulina boreal marine transgression.
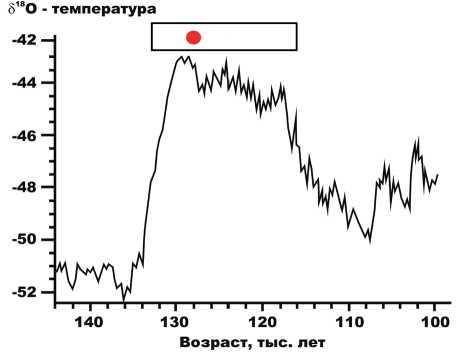
Таблица 1 Гранулометрический и нормативно-минеральный составы грунтов, обнаруженных во внутренней части бивня
Table 1 Granulometric and normative-mineral composition of grounds found in the inner part of the tusk
|
Гранулометрические фракции |
Содержание, мас. % |
Нормативно-минеральный состав, мол. % |
|||||
|
Кварц |
Альбит |
Слюда |
Хлорит |
Апатит |
Гетит |
||
|
Гравий (+2 мм) |
38.81 |
36.37 |
6.53 |
29.24 |
22.76 |
3.82 |
1.28 |
|
Песок гр/з (–2+1 мм) |
18.46 |
25.02 |
13.0 |
26.25 |
27.82 |
3.96 |
3.95 |
|
Песок кр/з (–1+0.5 мм) |
23.49 |
24.0 |
8.77 |
27.82 |
32.18 |
7.23 |
0 |
|
Песок ср/з (–0.5+0.25 мм) |
18.46 |
||||||
|
Песок м-т/з (–0.25+ 0.01 мм) |
0.78 |
||||||
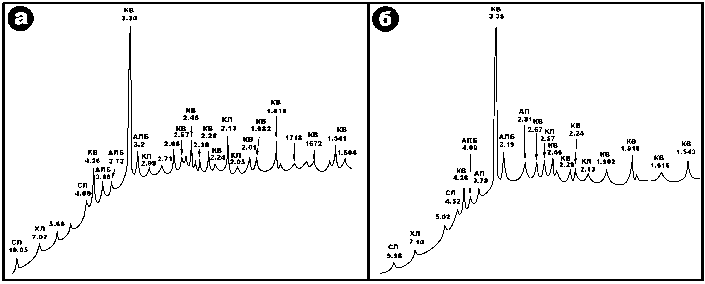
Рисунок 4. Рентгенофазовый состав вмещающих грунтов, извлеченных из бивня ископаемого моржа. Минералы: Кв – кварц, АЛб – альбит, СЛ – слюда, ХЛ – хлорит, КЛ – кальцит, АП – апатит.
Figure 4. X-ray phase composition of grounds extracted from the tusk of fossil walrus. Minerals: КВ – quartz, АЛБ – albite, СЛ – mica, ХЛ – chlorite, КЛ – calcite, АП – apatite.
соответствуя не тиллам, а, скорее, терригенным морским или аллювиальным осадкам Русской плиты.
Материалы и методы
Предметом непосредственного исследования послужили небольшие фрагменты левого бивня (рис. 2, в). В ходе работы использован широкий комплекс современных минералогических методов, хорошо себя зарекомен- довавших в приложении ко многим палеонтологическим объектам [1–5]: оптическая микроскопия (компьютеризированный комплекс OLYMPUS BX51); термический анализ (DTG-60А/60 АН, Shimadzu); определение содержания Сорг методом кулонометрического титрования; рентгенофлюоресцентный анализ (XRD-1800 Shimadzu); рентгеновская дифрактометрия (XRD-6000); аналитическая растровая электронная микроскопия (JSM-6400 Jeol; Tescan Vega); газовая хроматография – анализ элементного состава коллагена (EA 1110 (CHNS–O)); газовая хроматография – анализ состава аминокислот в органическом веществе (GC-17A Shimadzu с пламенно-ионизационным детектором); масс-спектрометрия с индуктивно связанной плазмой (NexION 300S Perkin Elmer); масс-спектрометрический анализ изотопного состава С, О в биоапатите и С, N в костном коллагене (Delta V. Avantage с аналитическим комплексом Thermo Fisher Scientific).
Результаты и их обсуждение
Исследованные фрагменты бивня характеризуются относительной прочностью и буроватым цветом. Валовый химический состав (мас. %): SiO2 – 2.55; Al2O3 – 0.96; Fe2O3 – 3.81; MnO – 0.9; MgO – 0.81; CaO – 58.71; SrO – 0.30; Na2O – 0.87; K2O – 0.14; P2O5 – 29.69; SO3 – 1.26. Около 12 мас. % здесь составляют иллювиированные в кость минеральные примеси, обусловленные средой захоронения. Пересчеты приводят к следующему фазовому составу и нормативному содержанию минеральных примесей (мол. %): кварц – 0.99; альбит – 0.52; слюда – 1.29; хлориты – 2.49; пирит – 1.45; гетит – 3.21. Из приведенных данных следует, что иллювиированная примесь в исследованной кости, по сравнению с вмещающими ее супесями отличается гораздо более глинистым (слюдисто-хлоритовым) составом. Кроме того, кость обогащена бактериогенным пиритом и гетитом – продуктом гидролиза пирита.
В составе исследованного бивня обнаружены 46 микроэлементов, общее содержание ко торых превышает 3 мас. % (табл. 2). По этому показателю бивень превосходит практически все исследованные нами ископаемые кости плейстоценовых животных (рис. 5), что подтверждает относительную древность печорского моржа. В состав микроэлементов, находящихся в бивне, входят девять эссенциальных (жизненно-необходимых), 18 физиогенно-активных (разного происхождения) и 19 абиогенных (элементы, обусловленные фоссилизацией). От-
Таблица 2
Содержание микроэлементов, г/т
Content of trace elements, ppm
Table 2
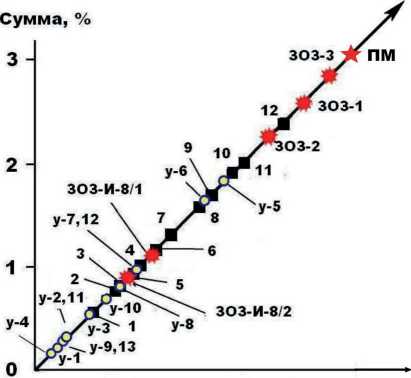
Продолжительность фоссилизации
Рисунок 5. Суммарное содержание микроэлементов в бивне ископаемого моржа – ПМ; в сравнении с костным детритом лошадей со стоянки Заозерной – ЗОЗ [4], мамонтовой фауны из Печорского Приуралья – 1–12 [5], плейстоценовых млекопитающих со стоянки Ушбулак в Казахстане – У-1-13 [1].
Figure 5. The total content of trace elements in the tusk of fossil walrus – ПМ – compared with bone detritus of horses from the Zaozernaya site – ЗОЗ [4], mammoth fauna from the Pechora Cis-Urals, 1–12 [5], fossil bones of Pleistocene mammals from the Ushbulak site in Kazakhstan – у–1–13 [1].
Биоапатит
Биоапатит в бивне печорского моржа характеризуется типичной рентгеновской дифрактограммой, в которой сильно преобладает несколько уширенный и слабо расщепленный пик основного отражения (рис. 6). Упомянутые выше минеральные примеси проявились на рентгенограмме незначительно. Величина рентгеновского индекса кристалличности биоапатита, оцененная по отношению интенсивностей отражений I300/I121, составила для исследуемого бивня 0.06, что достаточно низко для столь древних ископаемых костей скелета, не говоря уже о биоапатите в зубах млекопитающих, в котором индекс кристалличности варьируется в диапазоне 0.35–0.5.
По составу биоапатит в исследуемом бивне является фоссилизационно-измененным как в катионной, так и анионной подрешетках (табл. 3). В качестве наиболее существенных эпигенетических примесей к кальцию выступают железо и марганец, фосфор частично замещен серой и углеродом. Кристаллохимическая формула минерала имеет вид (Ca9.57–9.81Fe0.13–0.32Mn0–0.13)10[P4.92–5.05S0.17–0.26C0.7–0.91O24] (OH)1.26–1.53. Значение апатитового модуля (Ca/Pат) колеблет- ся в пределах 1.89–1.97, что соответствует существенно измененному биоапатиту в плейстоценовых костях, захороненных в открытых грунтах.
В режиме аналитической СЭМ в исследованных образцах выявлена характерная для ископаемых костей ассоциация микроминералов, в которую входят вторичные фосфаты, карбонаты, фрамбоидальный пирит, множество самородных металлов и сплавов, представленных инди-
Таблица 3
Химический состав (мас., %) и кристаллохимические формулы биоапатита в бивне печорского моржа
Table 3
Chemical composition (wt., %) and crystal chemical formulas of bioapatite in the tusk of the Pechora walrus
|
P 2 O 5 |
SO3 |
CaO |
MnO |
Fe2O3 |
Эмпирические формулы |
Са/Pat |
|
36.85 |
1.51 |
57.23 |
н.о. |
4.41 |
(Ca 9.81 Fe 0.19 ) 10 [(P 4.98 S 0.18 C 0.84 ) 6 O 24 ](OH) 1.34 |
1.97 |
|
37.43 |
1.99 |
57.03 |
1.00 |
2.55 |
(Ca 9.57 Fe 0.30 Mn 0.13 ) 10 [(P 4.95 S 0.23 C 0.82 ) 6 O 24 ](OH) 1.41 |
1.93 |
|
37.86 |
1.91 |
57.07 |
0.61 |
2.55 |
(Ca 9.58 Mn 0.12 Fe 0.30 ) 10 [(P 5.01 S 0.22 C 0.77 ) 6 O 24 ](OH) 1.45 |
1.91 |
|
37.80 |
2.15 |
56.54 |
0.77 |
2.74 |
(Ca9.57Mn0.11Fe0.32)10[(P5.05S0.25C0.70)6O24](OH)1.45 |
1.89 |
|
37.79 |
2.19 |
57.99 |
0.91 |
1.12 |
(Ca 9.75 Mn 0.12 Fe 0.13 ) 10 [(P 5.01 S 0.26 C 0.73 ) 6 O 24 ](OH) 1.53 |
1.95 |
|
37.58 |
1.44 |
58.50 |
0.63 |
1.85 |
(Ca 9.70 Mn 0.09 Fe 0.21 ) 10 [(P 4.92 S 0.17 C 0.91 ) 6 O 24 ](OH) 1.26 |
1.97 |

Рисунок 7. Биоапатит (а) и микроминеральные примеси в нем (б–ж). Микроминералы: Ф – Ca-Fe фосфат; К – кальцит; Cu-Fe-Sn – железистая бронза; Cr – самородный хром; Сu-Fe-Ni – никеле-во-железо-медный сплав. СЭМ-изображения в режимах вторичных (а, г) и упруго-отраженных (б, в, д–ж) электронов.
Figure 7. Bioapatite (a) and micromineral impurities in it (б–ж). Microminerals: Ф – Ca-Fe phosphate; K – calcite; Cu-Fe-Sn – iron bronze; Cr-Fe – native chromium; Сu-Fe-Ni – nickel-iron-copper alloy. SEM images in the modes of secondary (a, г) and elastically reflected (б, в, д–ж) electrons.
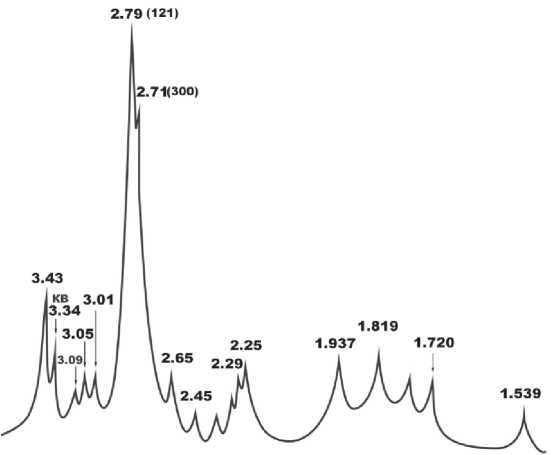
Рисунок 6. Ретгеновская дифрактограмма биоапатита в бивне печорского моржа.
Figure 6. X-ray diffraction pattern of bioapatite in the tusk of the Pechora walrus.
видами размером от субмикронного до 5–10 мкм (рис. 7). Их фазовая диагностика производилась по энергодисперсионным спектрам (рис. 8).
Химический состав вторичных фосфатов (мас. %): P2O5 – 31.03–32.71; CaO – 26.98–28.77; MnO – 0–0.65; Fe2O3 – 38.91–39.92; K2O – 0.39–0.64. Кристаллохимическая формула – (Ca 1.05–1.18 Fe 1.08–1.11 K 0.02–0.03 )[PO 4 ](OH) 1.28–1.65 . Наложенные на биоапатит карбонаты представлены железосодержащим кальцитом с формулой (Ca0.98–1Fe0–0.02)[CO3]. Самородно-металлические фазы: Ni-Fe-Cu cплавы состава Cu 0.39–0.4 Fe 0.29–0.34 Ni 0.25–0.29 Zn 0.01–0.02 ; железистая бронза Cu 0.6– 0.61 Sn 0.15–0.16 Fe 0.23–0.25 ; самородный хром Cr 0.98–1 Fe 0–0.02 .
Органическое вещество
На кривых нагревания материала исследуемого бивня (рис. 9) зарегистрированы три эффекта: эндотермический с максимумом при 106 °С, отвечающий потере сорбционной воды, и два экзотермических с максимумами при 314 и 405 °С, обусловленных выгоранием соответственно низко- и высокомолекулярного органического вещества [6]. Последние два пика весьма сильно различаются по интенсивности, что характерно именно для плейстоценовых костей, в которых органическое вещество со временем молекулярно деградирует.
Выделенное из бивня методом химической деминерализации органическое вещество характеризуется преимущественно светло-бурым цветом и хорошо сохранившейся фибриллярной структурой (рис. 10). Его элементный состав определялся пирохроматографическим методом (мас. %): C = 44.5: N = 16.59: H = 6.8. Атомное отношение C/N в нем находится на уровне 3.35, что свидетельствует о хорошей химической сохранности органического матрикса в кости печорского моржа.
В составе исследуемого органического вещества определено 14 аминокислот (далее – АК), общее содержание которых оказалось выше, чем в ранее изученных нами ископаемых костях плейстоценовых млекопитающих (табл. 4). Высокие содержания глицина, аланина, пролина и гидроксипролина свидетельствуют о том, что основная масса исследованного материала является коллагеном. Большая часть АК (9 из 14) представлена одним энантиомером – L, в некоторых АК (аланин, аcпарагиновая и глутаминовая кислоты) обнаружены оба энантиомера – L и D – при соотношении содержаний D/L в диапазоне 0.01–0.05, что говорит о незначительной бактериальной переработке материала.
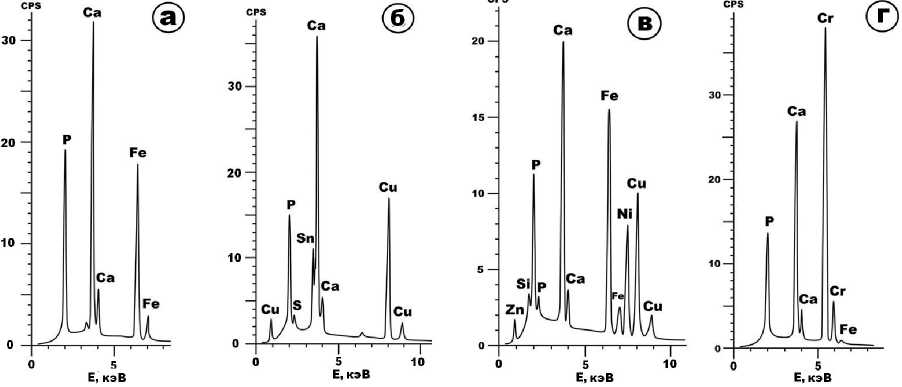
Рисунок 8. Энергодисперсионные спектры микроминералов в биоапатите печорского моржа: Ca-Fe-фосфат (а); железистая бронза (б); никель-желе-зо-медный сплав (в); железосодержащий хром (г).
Figure 8. Energy-dispersive spectra of microminerals in bioapatite of the Pechora walrus: Ca-Fe-phosphate (a); iron bronze (б); nickel-iron-copper alloy (в); iron-containing chromium (г).
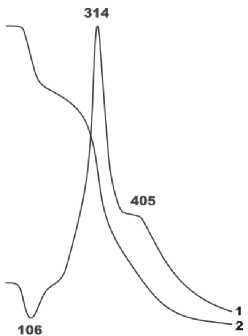
Рисунок 9. Данные термографического анализа материала ископаемого моржа, кривые: 1 – нагревания (температура, °C), 2 – потеря веса. Figure 9. The thermographic analysis data of fossil walrus’ material: 1 – heating (temperature, °C), 2 – weight losses.

Рисунок 10. Органическое вещество, выделенное из бивня печорского моржа.
Figure 10. Organic matter isolated from the tusk of the Pechora walrus.
Все полученные по АК дан- ные свидетельствуют о хорошей сохранности коллагена и соответствии с приведенной выше оценкой его биологического возраста. Последовательность сокращения групповых содержаний АК в исследованном образце имеет вид: алифатиче- ские > имино (особенно характерные для костного коллагена животных) > ароматические > кислые > гидроксильные > основные. Для плейстоценовых лошадей со стоянки Заозерной, плейстоценовых растительноядных млекопи- тающих со стоянки Ушбулак и медведей из пещеры Има-най аналогичная последовательность несколько отличается: алифатические > имино > кислые > гидроксильные > ароматические > основные. Из сопоставления следует, что органический матрикс в бивне печорского моржа (рис. 11) характеризуется не только повышенной концентрацией ароматических АК (за счет тирозина), но и аномально высокими содержаниями лейцина (алифатические АК) и лизина (основные АК). Это свидетельствует о возможном присутствии в его составе примесей нерегулярных полимерных органических соединений типа меланоидинов, содержащих аминокислоты. Образование таких соединений на начальных этапах преобразования органического вещества было показано на примере костного детрита рыб [7]. Меланоидины обычно окрашены в золотистый или коричневый цвет, устойчивы к воздействию слабых кислот, а для их разрушения необходимы жесткие условия кислотного гидролиза.
Изотопия и палеоэкологические
реконструкции
Изотопный состав С, О в биоапатите и С, N в костном коллагене исследованного образца анализировался в Институте геологии ФИЦ Коми НЦ УрО РАН [8]. Дополнительно к этому изотопный анализ органического вещества в том же образце был осуществлен в Центре изотопных исследований Гронингенского университета (Нидерланды). В случае костного биоапатита его разложение производилось в ортофосфорной кислоте. Значения δ13С и δ18О рассчитывались относительно стандартов PDB (для углерода) и SMOW (для кислорода). Изотопный состав коллагена анализиро- вался в режиме постоянного тока гелия, при этом использовались международный стандарт USGS-40 (L-Glutamic acid) и лабораторный стандарт Acetanilide (C8H9NO). Погрешность определения значений изотопных коэффициентов составляла ± 0.1 ‰ (1σ).
На диаграмме изотопного состава биоапатита (рис. 12) показаны две серии данных, полученных для костей плейстоценовых животных. К первой серии относятся ископаемые кости, фоссилизированные во внепещерных условиях: мамонтовая фауна, плейстоценовые лошади и другие растительноядные млекопитающие. В таких костях вариации значений изотопных коэффициентов для биоапатита лежат в пределах (‰): δ13С = –9 …–15; δ18О = 16–23. Во вторую серию включены кости плейстоценовых млекопитающих, претерпевшие пещерную фоссилизацию и вследствие этого получившие изотопное утяжеление как по углероду, так и кислороду (‰): δ13С = –2 …–10; δ18О = 19–27. Понятно, что такое утяжеление обусловлено изотопным влиянием на
Таблица 4
Аминокислотный состав органического вещества в бивне печорского моржа, мг/г
Table 4
Amino acid composition of organic matter in the tusk of the Pechora walrus, ppm
|
Аминокислоты (АК) |
Морж |
Плейстоценовые лошади [8] |
Плейстоценовые млекопитающие [7] |
Пещерные медведи (Иманай) |
|
Глицин C2H5O2N / Gly |
173.17 |
183.3 ± 96.36 |
180.03 ± 96.3 |
125.6 ± 24.18 |
|
Аланин C3H7O2N / Ala |
78.35 |
52.9 ± 43.45 |
84.48 ± 45.82 |
55.16 ± 10.07 |
|
Валин C5H11O2N / Val |
17.6 |
15.29 ± 12.18 |
22.19 ± 14.67 |
15.46 ± 3.44 |
|
Лейцин C6H13O2N / Leu |
40.43 |
20.01 ± 15.4 |
20.6 ± 19.1 |
20.28 ± 4.32 |
|
Изолейцин C6H13O2N |
5.99 |
21.79 ± 6.13 |
10.57 ± 7.16 |
5.36 ± 1.19 |
|
Алифатические АК |
315.54 |
213.69 ± 173.3 |
317.88 ± 168.86 |
181.38 ± 91.29 |
|
Фенилаланин C9H11O2N / Phe |
20.92 |
13.28 ± 9.86 |
19.02 ± 10.25 |
11.51 ± 2.55 |
|
Тирозин C9H11O3N / Tyr |
102.42 |
12.93 ± 13.07 |
31.94 ± 22.88 |
5.65 ± 7.22 |
|
Ароматические АК |
123.34 |
26.21 ± 22.91 |
57.07 ± 32.83 |
17.16 ± 8.88 |
|
Лизин C9H15O2N /Lys |
44.6 |
17.63 ± 15.32 |
27.98 ± 16.61 |
15.13 ± 2.87 |
|
Основные АК |
44.6 |
17.63 ± 15.32 |
27.98 ± 16.61 |
15.13 ± 2.87 |
|
Аспарагиновая кислота C4H7O4N / Asp |
33.98 |
28.57 ± 23.72 |
37.96 ± 21.31 |
21.1 ± 4.34 |
|
Глутаминовая кислота C5H9O4N Glu |
34.45 |
51.06 ± 42.69 |
68.58 ± 37.35 |
40.51 ± 7.75 |
|
Кислые АК |
73.43 |
79.63 ± 66.4 |
106.54 ± 58.59 |
61.62 ± 12.03 |
|
Серин C3H7O3N / Ser |
23.56 |
23.95 ± 20.13 |
53.8 ± 59.61 |
17.19 ± 9.8 |
|
Треонин C4H9O3N / Thr |
37.18 |
34 ± 39.08 |
32.51 ± 24.25 |
13.24 ± 2.85 |
|
Гидроксильные АК |
60.74 |
57.93 ± 58.78 |
86.31 ± 82.37 |
30.43 ± 12.15 |
|
Пролин C5H9O2N / Pro |
117.41 |
85.64 ± 73.79 |
123 ± 9 |
77.73 ± 14.86 |
|
Гидроксипролин C5H9O3N / Hyp |
69.41 |
51.82 ± 44.02 |
80.85 ± 45.28 |
43.28 ± 8.27 |
|
ИМИНО |
186.82 |
137.46 ± 117.25 |
204.75 ± 115.66 |
121.01 ± 22.89 |
|
Общее содержание |
804.47 |
533.64 ± 454.18 |
800.48 ± 463.17 |
476.88 ± 95.15 |
Содержание,
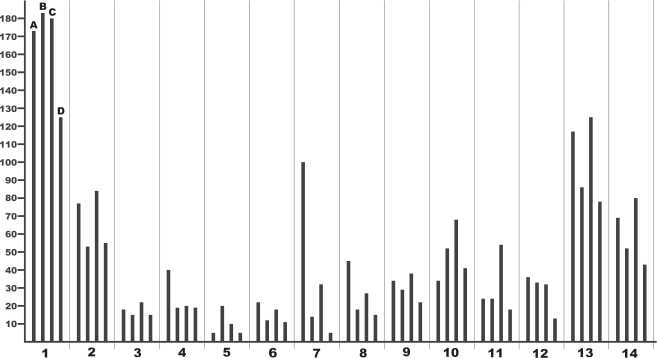
Рисунок 11. Пропорции между белковыми аминокислотами в коллагене печорского моржа (A), плейстоценовых лошадей со стоянки Заозерной (В), плейстоценовых млекопитающих со стоянки Ушбулак (С) и медведей из пещеры Иманай (D). По горизонтали – аминокислоты: 1 – глицин; 2 – аланин; 3 – валин; 4 – лейцин; 5 – изолейцин; 6 – фенилаланин; 7 – тирозин; 8 – лизин; 9 – аспарагиновая кислота; 10 – глутаминовая кислота; 11 – серин; 12 – треонин; 13 – гидроксипролин; 14 – пролин.
Figure 11. Proportions between protein amino acids in collagen of the Pechora walrus (A), Pleistocene horses from the Zaozernaya site (B), Pleistocene mammals from the Ushbulak site (C), and bears from the Imanay cave (D). Horizontally – amino acids: 1 – glycine, 2 – alanine, 3 – valine, 4 – leucine, 5 – isoleucine, 6 – phenylalanine, 7 – tyrosine, 8 – lysine, 9 – aspartic acid, 10 – glutamic acid, 11 – serine, 12 – threonine, 13 – hydroxyproline, 14 – proline.
специфичностью. Он содержит умеренно-легкий по изотопному составу углерод, не свойственный скелетам сугубо морских животных, например, раковинам фораминифер [9], но типичный для внепещерных костей плейстоценовых сухопутных животных, и изотопно-тяжелый кислород, характерный для гидрокарбоната морской воды. Такое изотопное сочетание можно объяснить маргинальной – сухопутно-морской средой обитания животного.
На диаграмме изотопного состава костного коллагена также выделяются две серии изотопных полей (рис. 13). Первая серия отвечает данным, полученным для континентальных животных – плейстоценовых лошадей, шерстистых носорогов и других растительноядных животных, бурых медведей, пещерных львов, медведей и гиен. Для коллагена этих животных характерны следующие диапазоны варьирования изотопных коэффициентов (‰): δ13С = –27 …–18; δ15N = 1–12. Вторая серия данных соответствует морским животным, костный коллаген которых по изотопным данным колеблется в принципиально иных диапазонах (‰): δ13С = –18 …–6; δ15N = 6–19. Данные для коллагена печорского моржа вполне вписываются в серию данных для морских животных, но при этом соответствующие точки оказались сдвинутыми относительно поля моллюсков – наиболее свойственной современным моржам пище – в сторону более изотопно-легкого углерода, свойственного морской рыбе. Это можно объяснить необычными для печорского моржа средой обитания и диетой, а именно его миграцией по р. Печоре вслед за поднимающейся по ней лососевой рыбой, например семгой. Сделанный вывод вполне согласуется с тем фактом, что изотопные данные для современных морских хищников соответствуют на рассматриваемой диаграмме полю с примерно теми же значениями изотопного состава углерода, но с более изотопно-тяжелым азотом.
биоапатит пещерной углекислоты, образующейся за счет морских карбонатолитов.
На фоне данных по внепещерным и пещерным ископаемым костям результаты изотопного анализа биоапатита в бивне печорского моржа характеризуются очевидной
Заключение
Исследованы фрагменты левого бивня плейстоценового моржа, ископаемые остатки которого были обнаружены в 2009 г. на эрозионном берегу р. Печоры. Проанализиро-
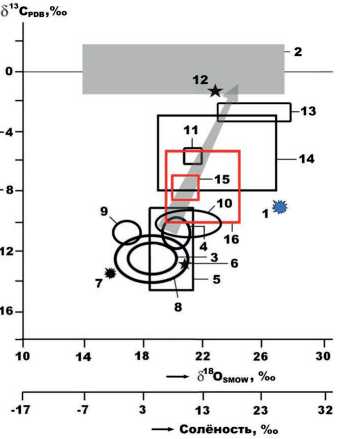
Рисунок 12. Изотопный состав биоапатита в ископаемых костях плейстоценовых млекопитающих на фоне морских карбонатных пород: 1 – бивень печорского моржа; 2 – карбонатолиты; 3 – мамонтовая фауна Западной Европы; 4 – мамонтовая фауна Печорского Приуралья; 5, 6 – соответственно мамонтовая фауна и неоплейстоценовый бурый медведь Омского Прииртышья (коллекция А.А. Бондарева); 7 – пещерная гиена с Южного Прибайкалья (образец от Д.В. Кобылкина); 8 – предголоценовые таймырские мамонты (образцы от Г.В. Шнейдера и Д.Н. Костина); 9 – плейстоценовые лошади со стоянки Заозерной; 10 – плейстоценовые млекопитающие со стоянки Ушбулак (Северо-Восточный Казахстан); 11, 12 – кости пещерного медведя с Печорского Приуралья с разной степенью каль-цитизации; 13, 14 – соответственно пещерные львы и медведи Западной Европы; 15, 16 – костный детрит соответственно львов и малых медведей ( Ursus «savini» ) из пещеры Иманай. Шкала солености воды построена по данным, вычисленным по уравнению В.Н. Кулешова. Стрелкой показан генеральный тренд изотопного утяжеления углерода и кислорода при переходе от ископаемых костей внепещерного захоронения к пещерным костным остаткам.
Figure 12. Isotope composition of bioapatite in fossil bones of Pleistocene mammals against marine carbonate rocks: 1 – tusk of the Pechora walrus; 2 – carbonatoliths; 3 – mammoth fauna of Western Europe; 4 – mammoth fauna of the Pechora Urals; 5, 6 – mammoth fauna and Neopleistocene brown bear of the Omsk Irtysh Region, respectively (collection of A.A. Bondarev); 7 – cave hyena from the Southern Baikal Region (sample from D.V. Kobylkin); 8 – pre-Holocene Taimyr mammoths (samples from G.V. Shneider and D.N. Kostin); 9 – Pleistocene horses from the Zaozernaya site; 10 – Pleistocene mammals from the Ushbulak site (North-East Kazakhstan); 11, 12 – bones of a cave bear from the Pechora Cis-Urals with different calcitization degree; 13, 14 – cave lions and bears of Western Europe, respectively; 15, 16 – bone detritus of lions and small bears ( Ursus “savini” ), respectively, from the Im-anay cave. The water salinity scale was built according to the data calculated by the V.N. Kuleshov’s equation. The arrow shows the general trend of isotope carbon and oxygen weighting in passing from fossil bones of an extracave burial to cave bone remains.
ваны гранулометрический, химический и нормативно-минеральный составы сохранившихся внутри бивня грунтов; термические свойства, химический и микроэлементый составы самого бивня; рентгеноструктурные параметры и химический состав костного биоапатита в нем; внешние свойства, строение, элементный и аминокислотный составы костного органического вещества. Все данные более или менее согласуются с геологическим возрастом и условиями захоронения костных остатков печорского моржа.
Особый акцент был сделан на изучении изотопного состава костного биоапатита и органического матрикса. На фоне полученных нами ранее данных по внепещерным
__ Морские
--животные
Морская
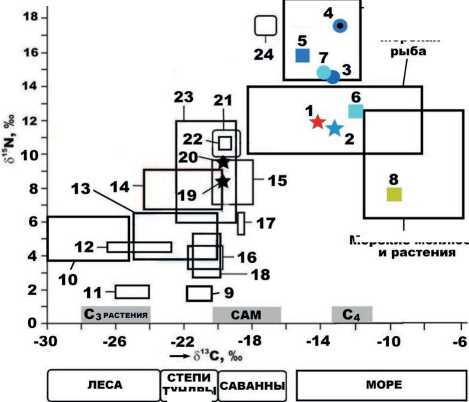
ТУНДРЫ
Морские моллюски
Рисунок 13. Изотопный состав костного коллагена, волос и кожи плейстоценовых и современных млекопитающих: 1 – бивень печорского моржа, определение в ИГ ФИЦ Коми НЦ УрО РАН; 2 – то же, определение в Гронингенском университете; 3–8 – зубы современных морских животных [10, 11], соответственно морж, касатка, гренландский тюлень, карликовый кашалот, дельфин-афалина, морская корова; 9 – плейстоценовые лошади со стоянки Заозерной; 10 – плейстоценовые млекопитающие со стоянки Ушбулак; 11 – носорог шерстистый; 12 – современные таежные бурые медведи (волосы и кожа); 13, 14 – соответственно медведи и львы из пещеры Иманай; 15, 16 и 17, 18 – соответственно до-LGM и пост-LGM популяции пещерных львов и медведей Западной Европы; 19–21 – соответственно ископаемые пещерная гиена, волк и росомаха с Западной Европы; 22 – ископаемая пещерная гиена с Южного Прибайкалья; 23 – современные бурые камчатские медведи (волосы и кожа); 24 – современные белые медведи с о. Вайгач (волосы и кожа). Шкала континентальных ландшафтов показана по Г. Бошрену и Д. Дракеру.
Figure 13. Isotopic composition of bone collagen, hair, and skin of Pleistocene and modern mammals: 1 – tusk of the Pechora walrus identified at the Institute of Geology, Federal Research Centre Komi Science Center of the Ural Branch of the Russian Academy of Sciences; 2 – the same, identified at the University of Groningen; 3–8 – teeth of modern marine animals [10, 11], walrus, killer whale, harp seal, pygmy sperm whale, bottlenose dolphin, sea cow; 9 – Pleistocene horses from the Zaozernaya site; 10 – Pleistocene mammals from the Ushbulak site; 11 – woolly rhinoceros; 12 – modern taiga brown bears (hair and skin); 13, 14 – bears and lions, respectively, from the Imanay cave; 15, 16 and 17, 18 – pre-LGM and post-LGM populations of West-European cave lions and bears, respectively; 19–21 – fossil cave hyena, wolf, and wolverine from Western Europe, respectively; 22 – fossil cave hyena from the Southern Baikal Region; 23 – modern brown Kamchatka bears (hair and skin); 24 – modern polar bears from the Vaigach Ireland (hair and skin). The scale of continental landscapes is shown according to G. Boshren and D. Dracker.
и пещерным ископаемым костям плейстоценовых млекопитающих, обитавших на суше, результаты изотопного анализа биоапатита печорского моржа характеризуются очевидной специфичностью. Он содержит умеренно изотопно-легкий углерод, свойственный внепещерным ископаемым костям плейстоценовых животных, сочетающийся с изотопно-тяжелым кислородом, типичным для гидрокарбоната морской воды и характерным именно для морских животных. Изотопные данные для коллагена печорского моржа коррелируются с аналогичными характеристиками морских животных, но при этом указывают на рыбно-морскую диету. Последнее свидетельствует о необычной для морских хищников среде обитания и необычном рационе питания печорского моржа, который осуществлял, вероятно, длительные миграции по реке, вслед за поднимающейся по ней лососевой рыбой.
Список литературы Плейстоценовый морж на реке Печоре: минералого-геохимические данные и палеоэкологические реконструкции
- Силаев, В.И. Костные остатки млекопитающих с палеонтологической стоянки Ушбулакверо-Восточный Казахстан): археологический контекст, минералого-геохимические свойства и палеоэкологические реконструкции / В.И. Силаев, А.А. Анойкин, Т.Д. Павленок, А.Ф. Хазов, В.Н. Филиппов [и др.] // Вестник геонаук. -2022. - № 4. - С. 3-37.
- Силаев, В.И. Минералого-геохимические особенности пещерной фоссилизации ископаемых костей на примере пещеры Иманай (Южный Урал) / В.И. Силаев, М.Н. Паршукова, Д.О. Гимранов, В.Н. Филиппов, Д.В. Киселева [и др.] // Вестник Пермского университета. Геология. -2020. - Т. 19 (4). - С. 323-358.
- Силаев, В.И. Современные исследования ископаемого костного детрита: палеонтология, минералогия, геохимия / В.И. Силаев, Д.В. Пономарев, Ю.С. Симакова, С.Н. Шанина, И.В. Смолева [и др.] // Вестник Института геологии Коми НЦ УрО РАН. - 2016. - № 5. - С. 19-31.
- Силаев, В.И. Опыт использования минералого-геохими-ческих свойств костных остатков на палеолитической стоянке Заозерье (Средний Урал) / В.И. Силаев, С.Н. Шанина, И.В. Смолева, А.Ф. Хазов, Е.А. Туманова // ПАЖМИ, - 2019. - № 2. - С. 35-77.
- Silaev, V.I. Mineralogical-geochemical characteristics of bone detritus of Pleistocene mammals as a source of paleontological information / V.I. Silaev, D.V. Ponomarev, D.V. Kiseleva, I.V. Smoleva, Yu. S. Simakova // Paleontological Journal. - 2017. - Vol. 51. - № 13. - P. 21-47.
- Votyakov, S.L. Thermal properties of fossilized mammal bone remnants of the Urals / S.L. Votyakov, D.V. Kisileva, Yu.V. Shcapova, N.G. Smirnov, N.O. Sadykova // Journal Thermal Analysis and Calorimetry. - 2010. - Vol. 101. - P. 63-73.
- Дроздова, Т.В. Геохимия аминокислот / Т.В. Дроздова. -Москва: Наука, 1977. - 199 с.
- Силаев, В.И. Ископаемый морж на р. Печоре: изотопно-геохимические данные и экологические реконструкции / В.И. Силаев, Д.В. Пономарев, И.В. Смолева, В.Н. Филиппов, А.Ф. Хазов [и др.] // Современные проблемы теоретической, экспериментальной и прикладной минералогии (Юшкинские чтения-2022): материалы российской конференции с международным участием. Сыктывкар: Геопринт, 2022. - С. 115-117.
- Ветошкина, О.С. Углерод-кислородная изотопия верхнеюрских фораминифер Saracenaria pravoslavlevi как показатель условий седиментации / О.С. Ветошкина, С.В. Лыюров, Д.А. Бушнев // Вестник Института геологии Коми НЦ УрО РАН. - 2013. - № 10. - С. 7-11.
- Fay F. H. Ecology and biology of the pacific walrus, Odobe-nus rosmarus divergens, Illiger // North American Fauna. 1982. No. 74. 279 p.
- Walker, J.L. Dietary studies of marine mammals using stable carbon and nitrogen isotopic rations of teeth / J.L. Walker, SA Mack // Marine Mammals Science. - 1999. -Vol. 15(2). - P. 3214-334.
