Подходы к анализу структуры растительного покрова ландшафтов с пересеченным рельефом

Автор: Королюк Андрей Юрьевич
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Структура и динамика растительных сообществ
Статья в выпуске: 1-5 т.14, 2012 года.
Бесплатный доступ
В статье продемонстрированы различные подходы анализа ландшафтов, в которых распределение растительных сообществ определяется экспозицией, крутизной и формой склона.
Растительный покров, пространственная структура, экологические факторы, модель сопки
Короткий адрес: https://sciup.org/148200805
IDR: 148200805 | УДК: 581.524
Analysis of spatial patterns in vegetation of landscapes with complicated relief
For landscapes, where the distribution on plant communities is determined with exposition, steepness and shape of their slopes, the different methods of landscape analysis are proposed.
Текст научной статьи Подходы к анализу структуры растительного покрова ландшафтов с пересеченным рельефом
Роль рельефа в организации растительного покрова общеизвестна. Во многих ландшафтах распределение типов растительных сообществ по склонам с различными характеристиками определяет основные черты пространственной структуры растительности. В то же время показатели крутизны, экспозиции и формы склона являются одними из наиболее легко определяемых характеристик экотопов. Эти же показатели, в виде усредненных величин, доступны из цифровых моделей рельефа. Одним из наиболее интересных объектов с точки зрения разработки и тестирования методов анализа распределения сообществ в ландшафте являются районы с пересеченным рельефом: балочные системы, мелкосопочники и горные системы.
На территории Южной Сибири большим разнообразием и высокой пространственной неоднородностью растительности выделяются лесостепные ландшафты. В них количество осадков примерно соответствует потенциальной испаряемости. При этом южные инсолируемые склоны испаряют большее количество воды, чем северные теневые, а соответственно растите льные сообщества здесь находятся в дефиците влаги, в то время как теневой склон может иметь достаточное увлажнение. Этот контраст возрастает с увеличением широты, так как при движении на север различия в инсоляции и те-плообеспеченности между северными и южными склонами увеличиваются. Так в верховьях Колымы (около 62 0 с.ш.) средняя температура III декады июня на поверхности почвы в степном сообществе на южном пологом склоне составляет 18,10, а на северном – 3,10 [1]. Сумма положительных температур составляет соответственно 24000 и 6600. Разница в теплообеспеченности еще более вырастает на крутых склонах. Контрастность термических условий на склонах разной экспозиции свойственна всем лесостепным ландшафтам умеренного пояса. Это определяет важность учета экспозиции и крутизны склона при исследовании взаимосвязей между растительными сообществами и экологическими факторами.
Целью нашей работы является разработка методик единообразного и формализованного описания
структуры растительности, базирующихся на совокупности конкретных геоботанических описаний для ландшафтов, структура растительного покрова которых определяется распределением типов сообществ по склонам различной экспозиции, крутизны и формы.
Перед собой мы поставили три вопроса:
-
1. Как оценить различия растительных сообществ на склонах разной экспозиции и как определить, какие экспозиции наиболее контрастны по растительности?
-
2. Как определить ведущие экологические факторы, отвечающие за различия в растительных сообществах на разных склонах?
-
3. Насколько эффективна предложенная нами ранее модели сопки [4] в применении к различным регионам?
В качестве основных тестовых объектов мы выбрали два ключевых участка, представляющих низкогорные лесостепные ландшафты Южной Бурятии – лиственничную и сосновую лесостепь бассейна р.Джида. Для построения моделей сопки были дополнительно использованы данные из различных регионов Алтае-Саянской горной области, Забайкалья и провинции Внутренняя Монголия Китая. Хранение и анализ данных проводились в программах IBIS 6.2 [2] и PAST 2.14 [6]. Анализировался фактор увлажненности почв, для определения положения геоботанических описаний на этом градиенте (статусы увлажнения сообществ) использовались оптимумы растений для Южной Сибири [3].
Для анализа контрастности растительных сообществ на склонах различной экспозиции был использован массив из 115 геоботанических описаний, выполненных в низкогорной лиственничной лесостепи в западной части Джидинского района Республики Бурятия. Описания выполнялись на площади 100 кв.м. и закладывались таким образом, чтобы охватить склоны различной крутизны и экспозиции. На каждой площадке выявлялся состав видов и проективное покрытие каждого из них в процентах, а также определялась экспозиция и крутизна склона в градусах.
Все описания были разнесены по восьми 45градусным секторам. Учитывались описания, выполненные на склонах 5 и более градусов. Это свя- зано с тем, что пологие склоны исследованного участка соответствуют подгорным шлейфам, на которых независимо от экспозиции развиваются зональные степи, близкие по флористическому составу. Единство экологических условий здесь определяется близкими показателями инсоляции. Включение в анализ пологих склонов нивелирует различия растительных сообществ склонов различной экспозиции, и степень нивелировки тем боль- ше, чем больше описаний пологих склонов мы используем в обработке. Для всех пар секторов были вычислены средние коэффициенты сходства по Съеренсену (таблица) между всеми парами описаний. Анализ таблицы показывает резкое отличие северных склонов от всех остальных. В целом наибольшие отличия наблюдаются между северными и южными (с включением ЮЗ и ЮВ) секторами.
Таблица. Средние коэффициенты Съеренсена между описаниями на склонах различной экспозиции в лиственничной лесостепи.
|
экспозиции |
00 –440 |
450 –890 |
90 0 – 1240 |
1250 – 1790 |
1800 – 2240 |
225 0 – 2690 |
2700 – 3140 |
3150 – 2590 |
|
00–440 |
0.40 |
0.25 |
0.17 |
0.14 |
0.15 |
0.15 |
0.22 |
0.36 |
|
450–890 |
0.25 |
0.38 |
0.31 |
0.27 |
0.30 |
0.31 |
0.35 |
0.22 |
|
900–1240 |
0.17 |
0.31 |
0.46 |
0.36 |
0.38 |
0.39 |
0.39 |
0.15 |
|
1250–1790 |
0.14 |
0.27 |
0.36 |
0.38 |
0.39 |
0.37 |
0.35 |
0.11 |
|
1250–1790 |
0.15 |
0.30 |
0.38 |
0.39 |
0.48 |
0.43 |
0.42 |
0.10 |
|
1800–2240 |
0.15 |
0.31 |
0.39 |
0.37 |
0.43 |
0.46 |
0.41 |
0.12 |
|
2250–2690 |
0.22 |
0.35 |
0.39 |
0.35 |
0.42 |
0.41 |
0.48 |
0.18 |
|
2700–3140 |
0.36 |
0.22 |
0.15 |
0.11 |
0.10 |
0.12 |
0.18 |
0.42 |
|
средний коэффициент сходства |
0.23 |
0.30 |
0.33 |
0.30 |
0.33 |
0.33 |
0.35 |
0.21 |
Для экспозиций от 3150 до 450 (теневые склоны) и от 1250 до 2250 (световые склоны) были выбраны описания для дальнейшего анализа. Аналогичные выборки были сделаны и для ключевого участка, представляющего сосновую лесостепь в правобережной части нижнего течения р.Джида (восточная часть Джидинского района).
На первом этапе было оценено сходство парциальных флор представляющих южные и северные склоны. Был вычислен коэффициент сходства Съе-ренсена для пары сводных описаний (флористические списки). Уровень сходства в лиственничной лесостепи (0.53) и сосновой лесостепи (0.58) близок, что позволяет говорить близком уровне контрастности флористического состава для двух ключевых участков. В обоих случаях более половины видов от общего состава флоры встречается и на южных, и на северных склонах.
Подсчет коэффициентов Брея-Кертиса между сводными описаниями с учетом встречаемости показал более значительные отличия между ключевыми участками: сосновая лесостепь - 0.47, лиственничная лесостепь - 0.32. Учет встречаемости позволяет снизить шум, который вносят единично встреченные виды, находки которых зачастую случаях носят случайный характер. Так как площадки под геоботанические описания закладывались таким образом, чтобы охватить склоны всех экспозиций и крутизны, то можно говорить о том, что встречаемость в этом случае отражает степень распространенности видов в совокупности растительных сообществ. Коэффициенты сходства Брея-Кертиса показывают нам, что, несмотря на близкую флористическую контрастность, контрастность растительности с учетом встречаемости в листвен- ничной лесостепи значительно выше, чем в сосновой.
Подсчет коэффициентов Брея-Кертиса с учетом активности видов, которая вслед за Л.И. Малышевым [5] высчитывалась как корень из произведения встречаемости вида на его среднее проективное покрытие, показал двукратные различия между ключевыми участками по фитоценотической контрастности северных и южных склонов: в сосновой лесостепи - 0.43, в лиственничной лесостепи - 0.23.
Был проведен анализ флористического состава склонов по сводным описаниям. Растения, встречаемость которых различается на 30 % и более мы рассматривали как дифференцирующие. В лиственничной лесостепи их количество для северных склонов составило 28 видов, имеющих преимущественно лесную экологию. Южные склоны отличают 32 степных вида. Число дифференцирующих видов значительно превышает число видов с высоким постоянством встречающихся на склонах обоих экспозиций (со средней встречаемостью более 15 %). Общих видов выявилось лишь 17 – это растения, обычные в луговых степях, сухих светлых лесах и в опушечных сообществах. В ландшафте сосновой лесостепи ситуация противоположна. Для северных склонов характерны 17 растений, для южных всего 7. Число общих видов составляет 41. Как дифференцирующие, так и общие виды в этих условиях представляют многочисленную группу степных ксерофитных растений.
Анализ экологических статусов описаний во многом согласуется с полученными результатами. В лиственничной лесостепи средняя разница увлажнения между северными и южными склонами достоверна и составляет почти 7 ступеней: 57 (ув- лажнение соответствует травяным лесам и настоящим лугам) и 50 (соответствует богаторазнотравным дерновиннозлаковым степям). Различия уровня увлажнения между склонами различной сосновой лесостепи составляют менее одной ступени различия и не достоверны. Здесь среднее увлажнение и северных, и южных склонов (50 ступень), соответствует богаторазнотравным дерновиннозла-ковым степям.
Для анализа связей флористического состава и фактора увлажнения на каждом участке с использованием теста Мантеля была вычислена корреляция между сходством флористического состава и разницей статусов увлажнения. Результаты показали, что в ландшафтах лиственничной лесостепи различия во флористическом составе описаний жестко связаны с различиями в степени увлажнения местообитаний – корреляция составляет 0.8194 (p=0.0000). Таким образом, можно утверждать, что на анализируемом участке увлажнение выступает основным лимитирующим фактором. Экспозиция склонов в условиях равновесия между количеством осадков и потенциальной испаряемостью вносит коренной вклад в пространственное перераспределение теполообеспеченности и увлажнения. Растительность на инсолируемых склонах развивается в условиях дефицита влаги, а увлажнение северных склонов достаточно для развития здесь сомкнутых лиственничных лесов с развитыми травяным и моховым ярусами.
Аналогичный анализ для сосновой лесостепи показал отсутствие значимой связи между флористическим сходством сообществ и разницей в степени увлажнения (r = 0.1549, p=0.0225). На ключевом участке и южные, и северные склоны находятся в условиях дефицита влаги. В силу этого на склонах всех экспозиций в напочвенном покрове доминируют степные многолетние травы.
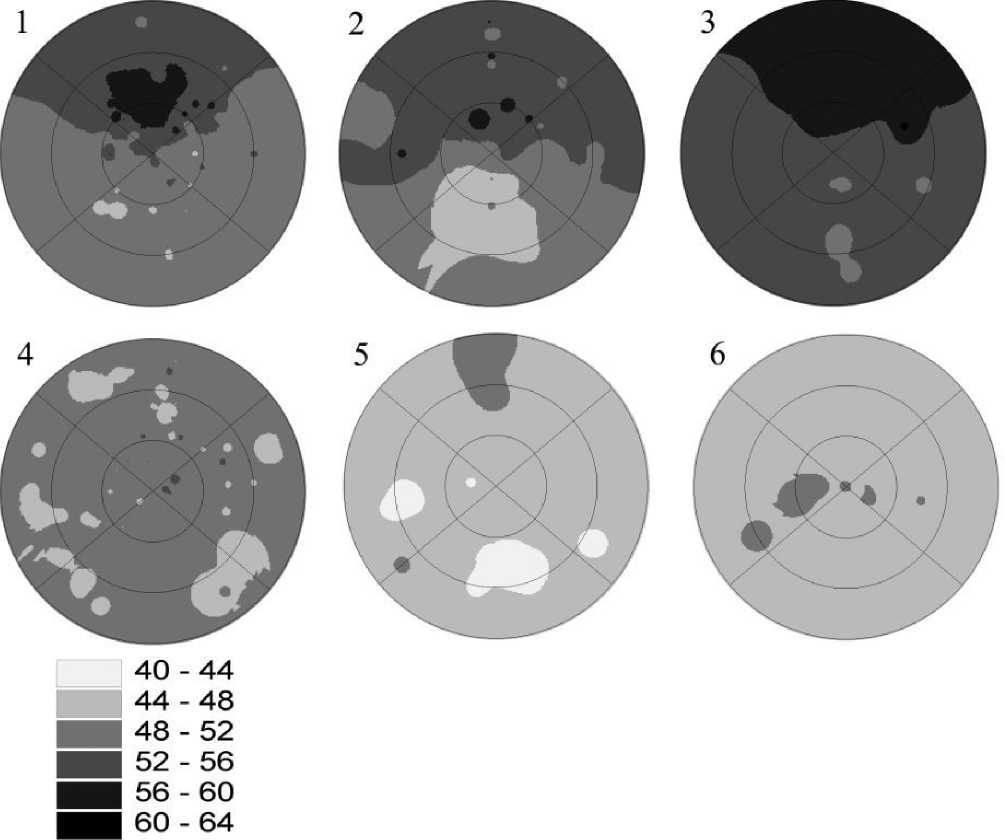
Рис . Модели сопки. Ландшафты с резкими экспозиционными различиями в растительности : 1. Лиственничная лесостепь Бурятии. 2. Березовая лесостепь Западного Алтая. 3. Березовая лесостепь Внутренней Монголии. Ландшафты со слабыми экспозиционными различиями в растительности : 4. Сосновая лесостепь Бурятии. 5. Степные мелкосопочники Западного Алтая. 6. Степные мелкосопочники Внутренней Монголии.
40-64 – ступени увлажнения. Внешнее кольце – склоны до 5 градусов крутизны, внутренний круг – более 15 градусов. Вертикально направление сверху вниз – С-Ю, горизонтальное слева направо – З-В.
Полученные результаты хорошо подтверждаются моделями сопки (рисунок). Для лиственничной лесостепи характерна четко выраженная эспозици-онная структура, в то время как в сосновой лесостепи склоны всех экспозиций соответствуют степному уровню увлажненности. Аналогичные закономерности мы можем наблюдать и в других регионах Внутренней Азии. Так, две модели сопки в Западном Алтае, располагающиеся на расстоянии около 50 км, кардинально различаются по выраженности экспозиционной структуры растительного покрова. Аналогичная пара моделей была построена для мелкосопочных ландшафтов Внутренней Монголии. Здесь также наблюдается рост экологической контрастности в ряду аридизации климата. Таким образом, в различных географических регионах прослеживаются очевидные аналогии в пространственно-структурной организации растительного покрова, хотя состав флоры может в значительной степени различаться.
Таким образом, использование простых показателей, характеризующих положение растительных сообществ в рельефе, дает возможности обоснованно говорить о существовании или отсутствии значимых связей между тем или иным экологическим фактором, с одной стороны, составом фитоценозов и структурой растительного покрова, с другой. Использование экологических шкал растений дает возможность, хотя и грубой, но в то же время быстрой оценки роли экологических факторов. Это может быть полезно для принятия решений о дальнейшей направленности экологических, как правило трудоемких и затратных исследований.
Как показывает наш опыт из различных областей Внутренней Азии, модель сопки является универсальным методом, позволяющим формализовано подойти к анализу пространственно-структурной организации растительного покрова ландшафтов с пересеченным рельефом.
Выражаю огромную признательность Аненхо-нову О.А., Бадмаевой Н.К., Санданову Д.В., Чепиноге В.В., Дулеповой Н.А. за организацию и проведение полевых исследований на территории Республики Бурятии, а также А.А. Звереву за помощь в обработке данных. Исследования проводились при поддержке Российского фонда фундаментальных исследований (грант 10-04-91159).
Список литературы Подходы к анализу структуры растительного покрова ландшафтов с пересеченным рельефом
- Берман Д.И., Алфимов А.В. Микроклиматическая обусловленность существования степных экосистем в субарктике северо-востока Азии//Бюлл. МОИП. Отд. биол. 1993. Т. 98, Вып. 3. С. 118-128.
- Зверев А.А. Информационные технологии в исследованиях растительного покрова. Томск: ТМЛ-Пресс, 2007. 304 с.
- Королюк А.Ю. Экологические оптимумы растений юга Сибири//Ботанические исследования Сибири и Казахстана. Вып.12. 2006, Барнаул-Кемерово, с. 3-38.
- Королюк А.Ю. Модель сопки -метод анализа структуры растительного покрова//Растительность России. 2008. № 13. С.117-122.
- Малышев Л.И. Флористическое районирование на основе количественных признаков//Бот. журн. 1973. Т. 58, № 11. С. 1581-1588.
- Hammer Ø., Harpe D.A.T., Ryan P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis//Palaeontologia Electronica. 2001. 4(1). 9 p.
