Поиск новых биоактивных соединений на основе микробной трансформации пентациклических тритерпеноидов олеананового ряда

Автор: Плотницкая Н.А., Гришко В.В., Ившина И.Б.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 2, 2024 года.
Бесплатный доступ
На фоне дефицита высокоэффективных фармакологических средств для лечения социально значимых заболеваний чрезвычайно актуален поиск новых химических соединений с биологической активностью на основе растительных пентациклических тритерпеноидов, в том числе олеаноловой (ОК) и глицирретовой (ГК) кислот. Данные тритерпеновые кислоты используются для получения производных с выраженной противовирусной, противомикробной, противовоспалительной, противоопухолевой и гепатопротекторной активностью. Преобразование тритерпеноидов в основном осуществляется с использованием химических методов, которые часто реализуются в условиях экстремальных показателей кислотности, температуры, применения дорогостоящих катализаторов или требуют введения защитных групп реакционно-активных центров молекулы. В настоящем обзоре изложены результаты собственных исследований и данные научной литературы последних лет, касающиеся оценки возможности применения биологической трансформации ОК и ГК на основе ферментативной активности микроорганизмов.
Олеаноловая кислота, глицирретовая кислота, микробная трансформация, биологически активные соединения
Короткий адрес: https://sciup.org/147244921
IDR: 147244921 | УДК: 579.222.4 | DOI: 10.17072/1994-9952-2024-2-176-189
Search for new bioactive compounds based on microbial transformation of oleanane type pentacyclic triterpenoids
Against the backdrop of a lack of highly effective pharmacological agents for the treatment of socially significant diseases, the search for new chemical compounds with biological activity based on plant pentacyclic triterpenoids, including oleanolic (OA) and glycyrrhetic (GA) acids, is urgent. These triterpene acids are used to obtain derivatives with pronounced antiviral, antimicrobial, anti-inflammatory, antitumor, and hepatoprotective activities. The transformation of triterpenoids is mainly carried out using chemical methods, requiring extreme acidity, temperature, the use of expensive catalysts or protective groups of the reactive centers of the molecule. This review presents the results of our own research and data from the scientific literature of recent years regarding the assessment of the possibility of using the biological transformation of OA and GA based on the enzymatic activity of microorganisms.
Текст обзорной статьи Поиск новых биоактивных соединений на основе микробной трансформации пентациклических тритерпеноидов олеананового ряда
Вторичные метаболиты растений - неисчерпаемый источник биологически активных веществ благодаря их доступности и большому структурному разнообразию [Atanasov et al., 2021]. Одной из интенсивно разрабатываемых сегодня групп растительных производных являются пентациклические тритерпеноиды, представленные соединениями ряда олеанана, урсана, фриделана, гопана и лупана. Тритерпеновые соединения олеананового ряда, в частности олеаноловая (ОК, 1 ) и глицирретовая (ГК, 2 ) кислоты (рис. 1), широко распространены в природе и характеризуются обширным спектром биоактивности. Однако молекулы тритерпеноидов отличаются повышенной гидрофобностью, что ограничивает их применение в качестве эффективных фармакологических агентов. Химическая трансформация ОК и ГК позволяет включать в структуру их молекул азот-, бром-, хлорсодержащие заместители, что значительно усиливает фармакологический потенциал производных. Полусинтетические производные ОК и ГК известны своим антиоксидантным [Sattarova et al., 2023], иммуномодулирующим [Rakhimova et al., 2023], противоопухолевым [Lisiak et al., 2023], антибактериальным [Blanco-Cabra et al., 2019], антитуберкулезным [Vishwakarma et al., 2022] и др. действием. Однако классический органический синтез биоактивных соединений протекает чаще всего в экстремальных условиях среды или требует применения дорогостоящих катализаторов, а также агентов защиты реакционно-активных центров молекулы. Перспективной альтернативой химическому синтезу являются реакции биотрансформации тритерпеновых соединений с использованием ферментативной активности целых клеток микроорганизмов, которые реализуются в нормальных и экологически безопасных условиях среды, с высоким уровнем регио- и стереоселективности, в одну технологическую стадию. Кроме того, микробная конверсия позволяет осуществлять специфическую модификацию недоступных для химических методов сайтов тритерпеновых молекул [Shah et al., 2014]. Опубликованные ранее специализированные обзоры, посвященные анализу процессов микробной трансформации терпеноидов, включают информацию за период до 2014 года [Гришко, Ногови-цина, Ившина, 2014; Azerad, 2016]. С учетом активного развития данной области исследований в последнее десятилетие обобщение и обсуждение сведений о процессах микробной трансформации пентацикли-ческих тритерпеноидов олеананового ряда и оценка биоактивности их производных представляется актуальным. В настоящей обзорной работе с привлечением собственных экспериментальных данных и литературных сведений последних лет отмечены целесообразность и перспективность применения метода микробной трансформации ОК и ГК для получения новых эффективных лекарственных агентов.
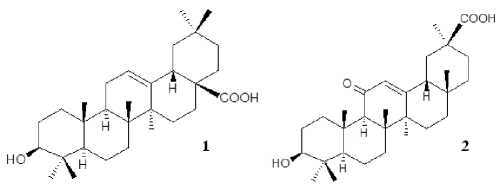
Рис. 1. Структурные формулы ОК ( 1 ) и ГК ( 2 ) [Structural formulas of OA (1) and GA (2) ]
Получение производных ОК и ГК с противовоспалительными свойствами
Подавляющее большинство известных метаболитов ОК и ГК, полученных в процессе микробной трансформации, характеризуется противовоспалительной активностью. Процессы трансформации ОК и ГК мицелиальными грибами, как правило, сопровождаются образованием широкого спектра метаболитов в результате введения гидроксильных или оксогрупп в положения С3, С7 и С15. Реже встречаются реакции карбоксилирования, ацетилирования и гликозилирования. Например, аскомицеты Trichothecium roseum (M 95.56) [Gong et al., 2014] и зигомицеты Circinella muscae AS 3.2695 [Yan et al., 2018] катализировали процесс окисления ОК с образованием 7β,15α-дигидрокси-3-оксо-ОК (3). При этом из среды биотрансформации T. roseum (M 95.56) был дополнительно выделен единичный интермедиат процесса мо- нодроксилирования – 15α-гидрокси-3-оксо-ОК (4) [Gong et al., 2014], в то время как зигомицетовые грибы C. muscae AS 3.2695 одновременно катализировали самые разнообразные реакции гидроксилирования и гликозилирования с образованием 7β-гидрокси-ОК (5), 7β,21β-дигидрокси-ОК (6), 7α,21β-дигидрокси-ОК (7), 7β,15α-дигидрокси-ОК (8), 7β-гидрокси-3-оксо-ОК (9), 7β,15α-дигидрокси-ОК 28-O-β-D-глюкопиранозилового эфира (10), 21β-гидрокси-ОК 28-O-β-D-глюкопиранозилового эфира (11) и ОК 28-O-β-D-глюкопиранозилового эфира (12) [Yan et al., 2018] (рис. 2).
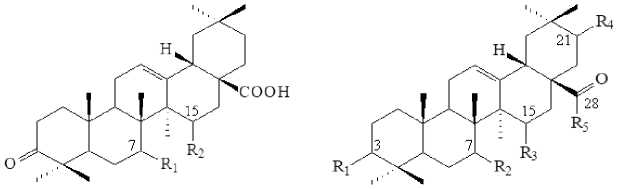
3R= pOH,R2= aOH
-
5: R] = Ra = ₽OH,R3 = & = H, R§ = OH
-
6: Ri = R2 = &= pOH, R3 = H, R$ = OH
-
7: R1 = R4=POH,R2 = aOH,R3 = H,& = OH
-
8: Rj = R2 = pOH,R3 = aOH, & = H, R5 = OH
-
9: R] = O?R2 = pOH,R3 = & = H 、 R5 = OH
-
10: Ri = R2 = pOH, R3 = aOH. R4 = H, R5 = G - p - £ - глюкогп q> ано зил
-
11: Ri = R4 = pOH, R2 = R3 = H 、 R5 = □- p- D- ЕЛЮКОГП ц^анознл
-
12: R] = pOH, R2 = R3 = R4 = H, R5 = C - p-D- ілюкопіірано зил
Рис. 2. Структурные формулы соединений 3–12
[Structural formulas of compounds 3–12 ]
Штамм C . muscae AS 3.2695 был также активен в отношении ГК и инициировал реакции окисления, ацетилирования и гликозилирования с образованием 7β-гидрокси-ГК ( 13 ), 15α-гидрокси-ГК ( 14 ), 7β,15α-дигидрокси-ГК ( 15 ), 7β-гидрокси-3-оксо-ГК ( 16 ), 7β,15α-дигидрокси-3-оксо-ГК ( 17 ), 7β-гидрокси-ГК 3- О -β- D -глюкопиранозида ( 18 ), 7β-гидрокси-ГК 3- О -β- D -6'- О -ацетил-глюкопиранозида ( 19 ), 15α-гидрокси-ГК 3- О -β- D -глюкопиранозида ( 20 ), 15α-гидрокси-ГК 3- О -β- D -6'- О -ацетил-глюкопиранозида ( 21 ) и 7β-гидрокси-ГК 30- О -β- D -глюкопиранозида ( 22 ) [Fan et al., 2019]. Этой же группой авторов показано, что зигомицет Rhizopus arrhizus AS 3.2893 осуществлял окислительную трансформацию ГК с образованием 7β-гидрокси-ГК ( 13 ), 15α-гидрокси-ГК ( 14 ), 7β,15α-дигидрокси-ГК ( 15 ), 3β-ацетокси-7β-гидрокси-ГК ( 23 ), 7-оксо-ГК ( 24 ), 7α-гидрокси-ГК ( 25 ) и 15α-гидрокси-7-оксо-ГК ( 26 ) [Fan et al., 2019] (рис. 3).
13 : R1 = R2 = pOH.R3 = H
15: R] = R? = pOH. R3 = aOH
L6: R] = O, Ra = ₽OH,R3 = H
17: Ri = O. R2 = BOH. R3 = aOH
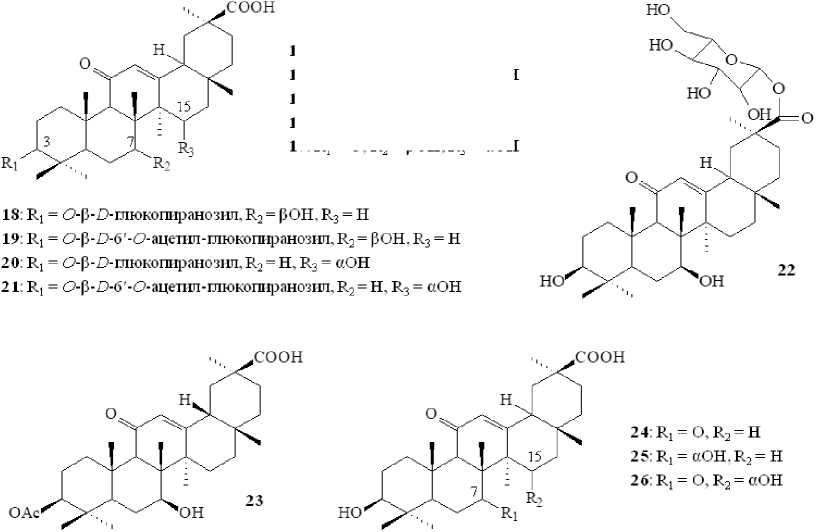
Рис. 3. Структурные формулы соединений 13–26 [Structural formulas of compounds 13–26 ]
Биологическое С7 и С15 гидроксилирование ГК с образованием в качестве основного метаболита 7β,15α-дигидрокси-ГК ( 15 ) протекало также с участием грибов Colletotrichum lini AS 3.4486 [He, Yang, Wu, 2015] и Absidia pseudocylinderospora ATCC 24169 [Maatooq et al., 2010] соответственно. Получение 7β-гидроксилированного производного ГК ранее было зафиксировано при использовании в качестве биокатализатора культур мукоровых зигомицетовых грибов рода Cunninghamella. Так, использование штамма C . elegans TSY-0865 приводило к трансформации ГК в единственное производное 7β-гидрокси-ГК ( 13 ) [Choudhary, Siddiqui, Nawaz, 2009], а C . blakealeana CGMCC 3.970 – к трансформации ГК в 7β-гидрокси-ГК ( 13 ), 3-оксо-7β-гидрокси-ГК ( 16 ) [Qin et al . , 2010], 15α,24-дигидрокси-ГК ( 27 ), 3-оксо-7β,15α,24-тригидрокси-ГК ( 28 ) и 3-оксо-7α,24-дигидрокси-ГК ( 29 ) [Ma et al . , 2017] (рис. 4).
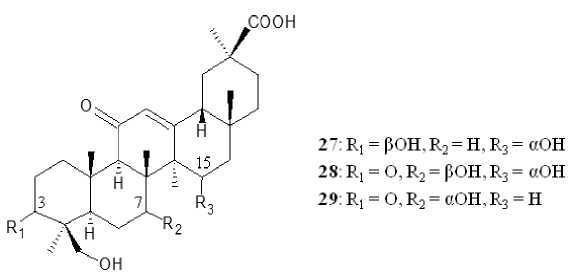
Рис. 4. Структурные формулы соединений 27–29 [Structural formulas of compounds 27–29 ]
Большинство из описанных выше производных ОК и ГК характеризуются противовоспалительным действием путем ингибирования липополисахарид (ЛПС)-индуцированной выработки NO в клетках RAW 264.7 при полумаксимальной ингибирующей концентрации (ИК 50 ) от 6.52 до 35.02 мкмоль. На примере 15α-гидрокси-ГК ( 14 ) группой авторов во главе с M. Zhang (2022) показано, что противовоспалительная активность тритерпеновых соединений может осуществляться путем избирательного действия на белок миелоидной дифференцировки 2 (MD2) для подавления активации сигнального пути ядерного фактора каппа B (NF-κB) посредством ингибирования ядерной транслокации p65 и подавления его целевых белков и генов в ЛПС-опосредованных RAW 264.7 клетках [Zhang et al., 2022]. Противовоспалительная активность производных тритерпеноидов показана также и на другой экспериментальной модели. Так, 15α,24-дигидрокси-ГК ( 27 ), 3-оксо-7β,15α,24-тригидрокси-ГК ( 28 ) и 3-оксо-7α,24-дигидрокси-ГК ( 29 ) продемонстрировали выраженную противовоспалительную активность в отношении нейронных тканей путем ингибирования ЛПС-индуцированного выделения NO в клетках микроглии мышей BV2 со значениями ИК 50 0.76, 0.94 и 0.16 ммоль/л соответственно [Ma et al., 2017]. В сравнительных исследованиях показано, что С7-гидроксилирование и С28-гликозилирование способствует повышению противовоспалительной активности производных ОК [Yan et al . , 2018], тогда как в случае ГК повышению ингибирующей активности в отношении выработки NO в клетках RAW 264.7 способствует гидроксилирование при 7 и 15 атомах углерода, а снижению – карбонилирование при 3 атоме углерода [Fan et al . , 2019].
При использовании бактериальных культур катализируемые реакции биотрансформации характеризуются более выраженной стерео- и региоселективностью. Так, бактериальная культура Streptomyces griseus ATCC 13273 катализировала процесс гидроксилирования и сайт-специфичного окисления С29 метильной группы ОК до карбоксильной с образованием 3β-гидрокси-олеан-12-ен-28,29-диовой кислоты ( 30 ), 3β,24-дигидрокси-олеан-12-ен-28,29-диовой кислоты ( 31 ) и 3β,21β,24-тригидрокси-олеан-12-ен-28,29-диовой кислоты ( 32 ) (рис. 5). Среди полученных соединений только 3β,21β,24-тригидрокси-олеан-12-ен-28,29-диовая кислота ( 32 ) проявляла противовоспалительный эффект путем ингибирования ЛПС-индуцированного выделения NO в клетках RAW 264.7 на уровне ИК 50 1.230 мкмоль [Xu, Zhang, et al., 2017]. Другая актиномицетальная культура Rhodococcus rhodochous ИЭГМ 1360 катализировала процесс С3-селективного окисления ОК с образованием 3-оксо-ОК ( 33 ) [Лучникова и др., 2023]. Ранее было показано, что 3-оксо-ОК проявляет выраженную противовоспалительную активность путем ингибирования продукции лейкотриена В в перитонеальных лейкоцитах крысы при ИК 50 17 мкмоль [Giner-Larza et al., 2001].
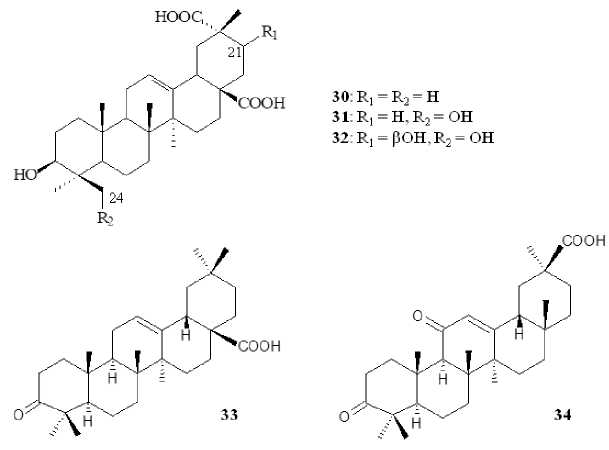
Рис. 5. Структурные формулы соединений 30–34 [Structural formulas of compounds 30–34 ]
Получение производных ОК и ГК с противоопухолевыми свойствами
Использование штамма Rhizomucor miehei CECT 2749 широко распространенных в природе грибов приводило к частичной метаболизации ОК с образованием 1β,30-дигидрокси-ОК ( 35 ), 7β,30-дигидрокси-ОК ( 36 ) и 30-гидрокси-ОК ( 37 ) [Martinez et al., 2013] (рис. 6). Соединение 36 , известное как кверетаровая кислота, впервые было выделено из произрастающих на территории Мексики кактусов Lemaireocereus queretaroensis и L . beneckei [Djerassi et al., 1956]. Ранее она была получена при трансформации ОК в течение 24 ч клетками E . coli , экспрессирующими гены цитохром P450 moxA актиномицетов Nonomuraea recticatena и Pseudomonas редокс-партнера camAB [Fujii et al., 2006]. Данное соединение проявляет in vitro умеренную противоопухолевую активность в отношении клеток HeLa (рак шейки матки) путем ингибирования эффектов, вызванных опухолевыми промоторами. В частности, кверетаровая кислота ингибирует 12- O -тетрадеканоилфорбол-13-ацетат-усиленное включение радиоактивной метки32P в структуру фосфолипидов опухолевых клеток [Kinoshita et al., 1999].
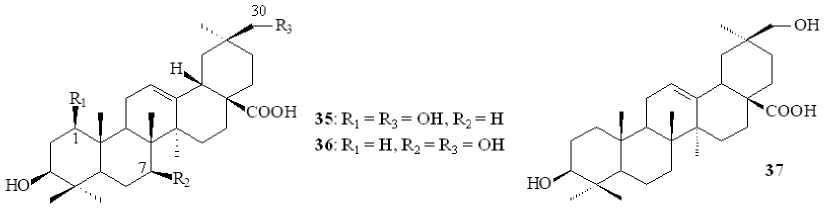
COOH
, COOH 35: Ri = R3 = OH 、 R2 = H
36:Rl = H.R2 = R3 = OH
Рис. 6. Структурные формулы соединений 35–37 [Structural formulas of compounds 35–37 ]
Актиномицеты рода Nocardia катализируют метилирование С28 карбоксильной группы пентацикли-ческих тритерпеноидов [Zhang et al., 2005]. Использование клеток N . iowensis (DSM 45197, NRRL 5646) в качестве биокатализатора процесса трансформации ОК приводило преимущественно к образованию метилового эфира ОК ( 38 ), а также небольшого количества метилового эфира 3-оксо-ОК ( 39 ) и не идентифицированного авторами метаболита 40 [Ludwig et al., 2015] (рис. 7). Показано, что процесс метилирования способствует повышению противоопухолевой активности тритерпеновых кислот. Так, метиловый эфир ОК 38 (10 мкмоль) ингибировал рост клеток SiHa (карцинома шейки матки), A-549 (карцинома легких) и IMR-32 (нейробластома) на 21, 48 и 55% соответственно и был в 2–7 раз более эффективен, чем ОК [Mallavadhani et al . , 2013]. Метиловый эфир 3-оксо-ОК ( 39 ) также характеризовался противоопухолевой активностью и подавлял рост клеток SR (лимфома), PC-3 (аденокарцинома простаты) и T-47D (рак груди) на 36, 18 и 24% соответственно при концентрации 10 мкмоль [Kaminskyy et al., 2012].
Рис. 7. Структурные формулы соединений 38–40 [Structural formulas of compounds 38–40 ]
Получение производных ОК и ГК с антибактериальными свойствами
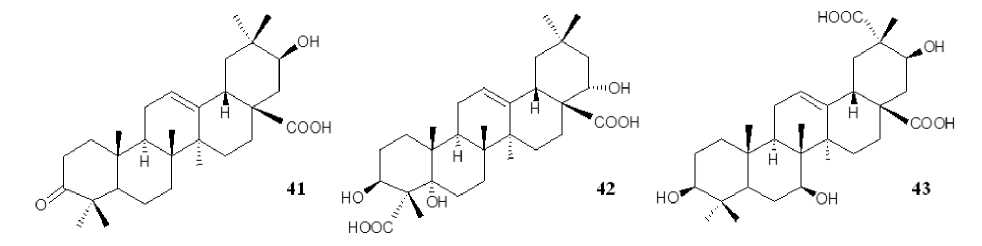
Одной из естественных функций тритерпеноидов как вторичных метаболитов является защита растений от инфекций [Guimaraes et al., 2019], в связи с чем многие производные ОК и ГК характеризуются выраженным антибактериальным действием. Трансформация ОК клетками грибов Mucor rouxii NRRL 1894 приводила к образованию 3-оксо-21β-гидрокси-ОК ( 41 ) [Capel et al., 2011], которая ранее была получена с использованием Penicillium chrysogenum [Hikino, Nabetani, Takemoto, 1971]. 3-Оксо-21β-гидрокси-ОК ( 41 ) проявляла выраженную антибактериальную активность в отношении вызывающей парадантоз бактерии Porphyromonas gingivalis (минимальная ингибирующая концентрация (МИК) 13.2 мкмоль) [Capel et al., 2011]. Полиоксигенирование ОК клетками штамма R . rhodochrous ИЭГМ 757 приводило к образованию нового производного – 5α,22α-дигидроксигипсогеновой кислоты ( 42 ) (рис. 8), которая обладала умеренной (МИК 0.781 мг/мл) активностью в отношении грамположительных бактерий Micrococcus luteus , Staphylococcus aureus , Bacillus subtilis [Luchnikova et al . , 2022]. Широко распространенное производное ГК 7β-гидрокси-ГК ( 13 ), полученное с использованием целого ряда культур грибов ( C . muscae AS 3.2695, R. arrhizus AS 3.2893 [Fan et al., 2019], C. blakesleana CGMCC 3.970 [Qin et al., 2010], C. elegans TSY-0865 [Choudhary, Siddiqui, Nawaz, 2009], R. chinensis CICC 40335 [Xu et al., 2020]), как и его С3-окисленное производное 3-оксо-7β-гидрокси-ГК ( 16 ), демонстрировали ингибирующую активность в отношении антибиотикоустойчивого бактериального штамма Enterococcus faecalis с зоной ингибирования роста 15–20 мм при концентрации 40 мкл [Qin et al., 2010].
но ноос'
Рис. 8. Структурные формулы соединений 41–43 [Structural formulas of compounds 41–43 ]
Получение производных ОК и ГК с другими видами биологической активности
Помимо противовоспалительных, противоопухолевых и антибактериальных свойств, характерных для производных ОК и ГК, в литературе представлены исследования и других типов биоактивности данных соединений. В дополнение к противовоспалительному эффекту показано, что ОК 28- O -β- D -глюкопиранозиловый эфир ( 12 ), образующийся в процессе трансформации ОК грибами C. muscae AS 3.2695 [Yan et al., 2018] или бактериями B. subtilis ATCC 6633 [Wang et al . , 2017], проявлял ингибирующий эффект в отношении ЛПС-стимулированной прокоагулянтной активности тканевого фактора в клетках THP-1 человека (ИК 50 0.035 нмоль). Авторами сделано предположение, что количество и положение свободных гидроксильных групп в структуре производных ОК влияет на их антикоагулянтный эффект [Wang et al . , 2017]. Другой широко распространенный противовоспалительный агент 7β,15α-дигидрокси-ГК ( 15 ) проявлял также антиоксидантную и гепатопротекторную активность. Показано, что данное соединение в концентрации 50 мг/кг значительно снижало уровень АЛТ, АСТ и малонового диальдегида, которые являются маркерами повреждения тканей печени. Кроме того, 7β,15α-дигидрокси-ГК ( 15 ) обеспечивала увеличение продукции NO в макрофагах крыс, обработанных CCl 4 [Maatooq et al . , 2010]. Нейропротекторный эффект 3β,7β,21β-тригидрокси-олеан-12-ен-28,29-диовой кислоты ( 43 ) (рис. 8), полученной с использованием S . griseus ATCC 13273, заключался в ее способности увеличивать выживаемость CoCl 2 -индуцированных нейроноподобных клеток PC12. При этом производные с С3-глюкопиранозиловой группой в структуре молекулы ОК теряли нейро-протекторные свойства, тогда как карбоксилирование при С29, вероятно, являлось ключевым для проявления данной активности [Xu et al . , 2020]. Антипаразитарная активность 3-оксо-ОК ( 33 ) заключалась в обеспечении 80% смертности популяции Meloidogyne incognita (галловая нематода) в течение 72 ч при концентрации 0.0625% [Begum et al . , 2015], а также ингибировании роста эпимастигот-ной формы Trypanosoma cruzi , вызывающей болезнь Шагаса, и промастиготной формы Leishmania amazonensis , вызывающей лейшманиоз, при ИК 50 18.5 и 29.9 мкмоль соответственно [Funari et al . , 2016]. С применением in silico анализа показана перспективность 3-оксо-ГК ( 34 ) в качестве агента ингибирования репликации вируса Sars-Cov-2 [Florez, Singh, 2020].
Заключение
Микробная трансформация пентациклических тритерпеноидов олеананового ряда является перспективным подходом для получения производных с выраженной биологической активностью, которые могут быть использованы для разработки новых высокоэффективных противовоспалительных, противоопухолевых, антибактериальных, антиоксидантных, антикоагулянтных, нейро- и гепатопро-текторных средств (таблица). Данный метод получения биологически активных соединений и их интермедиатов при условии его более активного развития как междисциплинарного инструмента перспективен для разработки новых лекарственных агентов для борьбы со злокачественными образованиями и нейродегенеративными заболеваниями, а также новых антибактериальных препаратов, эффективных в отношении антибиотикоустойчивых патогенных микроорганизмов. Стоит отметить, что среди эффективных биотрансформаторов ОК и ГК наиболее часто используются представители мицелиальных грибов, применение которых технологически нецелесообразно вследствие спорового характера роста и потенциальной опасности из-за их способности их к выработке микотоксинов с выраженным мутагенным и канцерогенным действием [Paterson, 2006; Dean et al., 2012]. При этом бактериальные катализаторы представлены лишь единичными видами родов Bacillus , Nocardia , Strepto-myces , среди которых нередко встречаются патогенные штаммы. Недавно появились сведения об успешном использовании штаммов непатогенных актиномицетов рода Rhodochrous , способных катализировать полную конверсию ОК и ГК в крайне высоких концентрациях [Лучникова и др., 2023; Luchnikova, Grishko, Ivshina, 2020; Luchnikova et al., 2022]. Необходимы дальнейшие исследования процессов биологической трансформации пентациклических тритерпеноидов олеананового ряда и поиск новых непатогенных бактериальных штаммов, использование которых позволит реализовать высокоэффективный синтез тритерпеновых производных с выраженной биологической активностью.
Биоактивные производные ОК и ГК, полученные в результате микробной трансформации [Biologically active OA and GA derivatives obtained by microbial transformation]
|
Соединение (номер структурной формулы) |
Механизм действия |
Микробный трансформатор |
|
Противовоспалительная активность |
||
|
7β,15α-дигидрокси-3-оксо-ОК ( 3 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 34.63 ± 1.74 мкмоль [Yan et al . , 2018] |
Trichothecium roseum (M 95.56) [Gong et al . , 2014] Circinella muscae AS 3.2695 [Yan et al . , 2018] |
|
7β-гидрокси-ОК ( 5 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 9.24 ± 0.62 мкмоль [Yan et al . , 2018] |
C. muscae AS 3.2695 [Yan et al . , 2018] |
|
7β,15α-дигидрокси-ОК ( 8 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 10.06 ± 2.17 мкмоль [Yan et al . , 2018] |
|
|
7β,15α-дигидрокси-ОК 28- O -β- D - глюкопиранозиловый эфир ( 10 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 8.28 ± 0.74 мкмоль [Yan et al . , 2018] |
|
|
21β-гидрокси-ОК 28- O -β- D - глюкопиранозиловый эфир ( 11 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 40.74 ± 2.84 мкмоль [Yan et al . , 2018] |
|
|
7β-гидрокси-ГК ( 13 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 19.32 мкмоль [Fan et al . , 2019] |
C. muscae AS 3.2695 [Fan et al., 2019] Rhizopus arrhizus AS 3.2893 [Fan et al . , 2019] Cunninghamella blakesleana CGMCC 3.970 [Qin et al . , 2010] C. elegans TSY-0865 [Choudhary, Siddiqui, Nawaz, 2009] R. chinensis CICC 40335 [Xu et al . , 2020] |
|
15α-гидрокси-ГК ( 14 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 20.24 мкмоль [Fan et al . , 2019] |
C. muscae AS 3.2695 [Fan et al . , 2019] R. arrhizus AS 3.2893 [Fan et al., 2019] Aspergillus niger RG13B1 [Zhang et al . , 2022] |
|
ОК 28- O -β- D -глюкопиранозиловый эфир ( 12 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 11.28 ± 1.03 мкмоль [Yan et al . , 2018] |
C. muscae AS 3.2695 [Yan et al . , 2018] Bacillus subtilis ATCC 6633 [Wang et al . , 2017] |
|
7β,15α-дигидрокси-ГК ( 15 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 9.07 мкмоль [Fan et al . , 2019] |
C. muscae AS 3.2695 [Fan et al . , 2019] R. arrhizus AS 3.2893 [Fan et al . , 2019] Colletotrichum lini AS 3.4486 [He, Yang, Wu, 2015] Absidia pseudocylinderospora ATCC 24169 [Maatooq et al . , 2010] |
|
3-оксо-7β-гидрокси-ГК ( 16 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 57.63 мкмоль [Fan et al . , 2019] |
C. muscae AS 3.2695 [Fan et al., 2019] C. blakesleana CGMCC 3.970 [Qin et al . , 2010] |
Продолжение таблицы
|
Соединение (номер структурной формулы) |
Механизм действия |
Микробный трансформатор |
|
7β,15α-дигидрокси-3-оксо-ГК ( 17 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 33.09 мкмоль [Fan et al., 2019] |
C. muscae AS 3.2695 [Fan et al., 2019] |
|
7β-гидрокси-ГК 3- О -β- D -глюкопиранозид ( 18 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 8.39 мкмоль [Fan et al., 2019] |
|
|
7β-гидрокси-ГК 3- О -β- D -6'- О -ацетил-глюкопиранозид ( 19 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 6.52 мкмоль [Fan et al., 2019] |
|
|
15α-гидрокси-ГК 3- О - β- D -глюкопиранозид ( 20 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 12.42 мкмоль [Fan et al., 2019] |
|
|
15α-гидрокси-ГК 3- О -β- D -6'- О -ацетил-глюкопиранозид ( 21 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 10.13 мкмоль [Fan et al., 2019] |
|
|
7β-гидрокси-ГК 30- О - β- D -глюкопиранозид ( 22 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 30.27 мкмоль [Fan et al., 2019] |
|
|
3β-ацетокси-7β-гидрокси-ГК ( 23 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 15.54 мкмоль [Fan et al., 2019] |
R. arrhizus AS 3.2893 [Fan et al., 2019] |
|
7-оксо-ГК ( 24 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 35.02 мкмоль [Fan et al., 2019] |
|
|
7α-гидрокси-ГК ( 25 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 20.43 мкмоль [Fan et al., 2019] |
|
|
15α-гидрокси-7-оксо- ГК ( 26 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 12.25 мкмоль [Fan et al., 2019] |
|
|
3β,21β,24-тригидрокси-олеан-12-ен-28,29- диовая кислота ( 32 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках RAW 264.7 при ИК 50 1.230 мкмоль [Xu, Wang, et al., 2017] |
Streptomyces griseus ATCC 13273 [Xu, Wang, et al., 2017] |
|
3-оксо-ОК ( 33 ) |
Ингибирование продукции лейкотриена В в перитонеальных лейкоцитах крысы при ИК 50 17 мкмоль [Giner-Larza et al . , 2001] |
Rhodococcus rhodochrous ИЭГМ 1360 [Лучникова и др., 2023] |
|
15α,24-дигидрокси-ГК ( 27 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках BV2 микроглии мышей при ИК 50 0.76 мкмоль/л [Ma et al . , 2017] |
C. blakesleana CGMCC 3970 [Ma et al . , 2017] |
|
3-оксо-7β,15α,24-тригидрокси-ГК ( 28 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках BV2 микроглии мышей при ИК 50 0.94 мкмоль/л [Ma et al . , 2017] |
|
|
3-оксо-7α,24-дигидрокси-ГК ( 29 ) |
Ингибирование ЛПС-индуцированного выделения NO в клетках BV2 микроглии мышей при ИК 50 0.16 мкмоль/л [Ma et al . , 2017] |
Противоопухолевая активность
|
30-гидрокси-ОК ( 37 ) |
Ингибирование усиленного 12-O-тетрадеканоилфорбол-13-ацетатом включение 32Pi в фосфолипиды опухолевых клеток на 41.4% при дозе 50 мкг/мл [Kinoshita et al . , 1999] |
R. miehei CECT 2749 [Martinez et al . , 2013] |
|
3-оксо-ГК ( 34 ) |
Ингибирование действия липоксигеназ при ИК 50 144.2 ммоль [Choudhary, Siddiqui, Nawaz, 2009] |
R. rhodochrous ИЭГМ 1360 [Лучникова и др., 2023] Fusarium lini NRRL-68751 [Choudhary, Siddiqui, Nawaz, 2009] |
|
Продолжение таблицы |
||
|
Соединение (номер структурной формулы) |
Механизм действия |
Микробный трансформатор |
|
3-оксо-ОК ( 33 ) |
Ингибирование роста клеток HepG2 при ИК 50 1.4 мкмоль [Irungu et al . , 2014]. Ингибирование роста клеток A549 при ИК 50 14.7 мкмоль, H460 при ИК 50 17.5 мкмоль, HGC27 при ИК 50 15.4 мкмоль [Wu et al . , 2011]. Ингибирование роста клеток B16-BL6 при 10.8 мкмоль [Huang et al . , 2006] |
R. rhodochrous ИЭГМ 1360 [Лучникова и др., 2023] |
|
Метиловый эфир ОК ( 38 ) |
Ингибирование роста клеток SiHa на 21%, HeLa на 18%, A-549 на 48%, IMR-32 на 55% при концентрации 10 мкмоль [Mallavadhani et al., 2013] |
Nocardia iowensis (DSM 45197, NRRL 5646) [Ludwig et al . , 2015] |
|
Метиловый эфир 3- оксо-ОК ( 39 ) |
Ингибирование роста клеток SR на 36%, PC-3 на 18%, T-47D на 24% при концентрации 10 мкмоль [Kaminskyy et al . , 2012] |
|
|
Антибактериальная активность |
||
|
5α,22α- дигидроксигипсогено-вая кислота ( 42 ) |
Ингибирование роста клеток Micrococcus luteus , Staphylococcus aureus , Bacillus subtilis при МИК 0.781 мг/мл [Luchnikova et al . , 2022] |
R. rhodochrous ИЭГМ 757 [Luchnikova et al . , 2022] |
|
3-оксо-21β-гидрокси-ОК ( 41 ) |
Ингибирование роста клеток Porphyromonas gingivalis при МИК 13.2 мкмоль [Capel et al . , 2011] |
Mucor rouxii NRRL 1894 [Capel et al . , 2011] Penicillium chrysogenum [Hikino, Nabetani, Takemoto, 1971] |
|
3-оксо-7β-гидрокси-ГК ( 16 ) |
Ингибирование роста клеток антибиотикоустойчивого штамма Enterococcus faecalis при концентрации 40 мкл (зона ингибирования 15 мм) [Qin et al . , 2010] |
C. muscae AS 3.2695 [Fan et al . , 2019] C. blakesleana CGMCC 3.970 [Qin et al . , 2010] |
|
7β-гидрокси-ГК ( 13 ) |
Ингибирование роста клеток антибиотикоустойчивого штамма Enterococcus faecalis при концентрации 40 мкл (зона ингибирования 20 мм) [Qin et al., 2010] |
C. muscae AS 3.2695 [Fan et al . , 2019] R. arrhizus AS 3.2893 [Fan et al . , 2019] C. blakesleana CGMCC 3.970 [Qin et al . , 2010] C. elegans TSY-0865 [Choudhary, Siddiqui, Nawaz, 2009] R. chinensis CICC 40335 [Xu et al . , 2020] |
|
Антикоагулянтная активность |
||
|
ОК 28- O -β- D -глюкопиранозиловый эфир ( 12 ) |
Ингибирование ЛПС-стимулированной прокоагулянтной активности тканевого фактора в клетках THP-1 человека при ИК 50 0.035 нмоль [Wang et al . , 2017] |
C. muscae AS 3.2695 [Yan et al . , 2018] B. subtilis ATCC 6633 [Wang et al . , 2017] |
|
Нейропротекторная активность |
||
|
3β,7β,21β-тригидрокси-олеан-12-ен-28,29- диовая кислота ( 43 ) |
Увеличение выживаемости CoCl 2 -индуцированных нейроноподобных клеткок PC12 при ЭД 50 0.48 мкмоль [Xu et al . , 2020] |
S. griseus ATCC 13273 [Xu et al . , 2020] |
|
Антипротозойная активность |
||
|
3-оксо-ОК ( 33 ) |
Обеспечение 80% смертности популяции Meloidogyne incognita в течение 72 ч при концентрации 0.0625% [Begum et al . , 2015] Ингибирование роста эпимастиготной формы Trypanosoma cruzi при ИК 50 18.5 мкмоль [Funari et al . , 2016] Ингибирование роста промастиготной формы Leishmania amazonensis при ИК 50 29.9 мкмоль [Funari et al . , 2016] |
R. rhodochrous ИЭГМ 1360 [Лучникова и др., 2023] |
|
Соединение (номер структурной формулы) |
Механизм действия |
Микробный трансформатор |
Антиоксидантная активность
|
7β,15α-дигидрокси-ГК ( 15 ) |
Защитное действие от перекисного окисления липидов, индуцированного аскорбатом железа, с антиоксидантным индексом 61±6.1 [Maatooq et al . , 2010] |
C. muscae AS 3.2695 [Fan et al . , 2019] R. arrhizus AS 3.2893 [Fan et al., 2019] C. lini AS 3.4486 [He, Yang, Wu, 2015] A. pseudocylinderospora ATCC 24169 [Maatooq et al . , 2010] |
Гепатопротекторная активность
|
7β,15α-дигидрокси-ГК ( 15 ) |
Дозозависимое стимулирование продукции NO в макрофагах крыс, обработанных CCl 4 . Снижение уровня АЛТ, АСТ и малонового диальдегида в гепатоцитах мышей, обработанных CCl 4 , при концентрации 50 мг/кг [Maatooq et al., 2010] |
C. muscae AS 3.2695 [Fan et al., 2019] R. arrhizus AS 3.2893 [Fan et al . , 2019] C. lini AS 3.4486 [He, Yang, Wu, 2015] A. pseudocylinderospora ATCC 24169 [Maatooq et al . , 2010] |
Список литературы Поиск новых биоактивных соединений на основе микробной трансформации пентациклических тритерпеноидов олеананового ряда
- Гришко В.В., Ноговицина Е.М., Ившина И.Б. Бактериальная трансформация терпеноидов // Успехи химии. 2014. Т. 83, № 4. С. 323-342.
- Лучникова Н.А. и др. Rhodococcus rhodochrous ИЭГМ 1360 - эффективный биокатализатор С3 окислительной трансформации олеанановых тритерпеноидов // Микробиология. 2023. Т. 92, № 2. С. 184196.
- Atanasov A.G. et al. Natural products in drug discovery: advances and opportunities // Nature Reviews Drug Discovery. 2021. Vol. 20. P. 200-216.
- Azerad R. Microbial transformations of pentacyclic triterpenes // Green Biocatalysis. 2016. P. 675-714.
- Begum S. et al. Nematicidal triterpenoids from Lantana camara // Chemistry and Biodiversity. 2015. Vol. 12. P. 1435-1442.
- Blanco-Cabra N. et al. Novel oleanolic and maslinic acid derivatives as a promising treatment against bacterial biofilm in nosocomial infections: an in vitro and in vivo study // ACS Infectious Diseases. 2019. Vol. 5. P. 1581-1589.
- Capel C.S. et al. Biotransformation using Mucor rouxii for the production of oleanolic acid derivatives and their antimicrobial activity against oral pathogens // Journal of Industrial Microbiology & Biotechnology. 2011. Vol. 38. P. 1493-1498.
- Choudhary M.I., Siddiqui Z.A., Nawaz S.A. Microbial transformation of 18p-glycyrrhetinic acid by Cunninghamella elegans and Fusarium lini, and lipoxygenase inhibitory activity of transformed products // Natural Product Research. 2009. Vol. 23. P. 507-513.
- Dean R. et al. The top 10 fungal pathogens in molecular plant pathology // Molecular Plant Pathology. 2012. Vol. 13. P. 414-430.
- Djerassi C. et al. Terpenoids. XXIV. The structure of the cactus triterpene queretaroic acid // Journal of the American Chemical Society. 1956. Vol. 78. P. 3783-3787.
- Fan B. et al. Anti-inflammatory 18p-glycyrrhetinic acid derivatives produced by biocatalysis // Planta Medica. 2019. Vol. 85. P. 56-61.
- Florez H., Singh S. Bioinformatic study to discover natural molecules with activity against COVID-19 // F1000Research. 2020. Vol. 9. P. 1-15.
- Fujii Y. et al. Hydroxylation of oleanolic acid to queretaroic acid by cytochrome P450 from Nonomuraea recticatena // Bioscience, Biotechnology, and Biochemistry. 2006. Vol. 70. P. 2299-2302.
- Funari C.S. et al. Oleanonic acid from Lippia lupulina (Verbenaceae) shows strong in vitro antileishmanial and antitrypanosomal activity // Acta Amazonica. 2016. Vol. 46. P. 411-416.
- Giner-Larza E.M. et al. Oleanonic acid, a 3-oxotriterpene from Pistacia, inhibits leukotriene synthesis and has anti-inflammatory activity // European Journal of Pharmacology. 2001. Vol. 428. P. 137-143.
- Gong T. et al. Microbial transformation of oleanolic acid by Trichothecium roseum // Journal of Asian Natural Products Research. 2014. Vol. 16. P. 383-386.
- Guimaraes A.C. et al. Antibacterial activity of terpenes and terpenoids present in essential oils // Molecules. 2019. Vol. 24. P. 2471.
- He C.J., Yang Y.M., Wu K.Y. Microbial transformation of glycyrrhetinic acid by Colletotrichum lini AS 3.4486 // Advanced Materials Research. 2015. Vol. 1120-1121. P. 877-881.
- Hikino H., Nabetani S., Takemoto T. Microbial transformation of oleanolic acid // Yakugaku Zasshi. 1971. Vol. 91. P. 637-640
- Huang D. et al. Anti-tumor activity of a 3-oxo derivative of oleanolic acid // Cancer Letters. 2006. Vol. 233. P. 289-296.
- Irungu B.N. et al. Constituents of the roots and leaves of Ekebergia capensis and their potential antiplasmodial and cytotoxic activities // Molecules. 2014. Vol. 19. P. 14235-14246.
- Kaminskyy D. et al. Synthesis of new potential anticancer agents based on 4-thiazolidinone and oleanane scaffolds // Medicinal Chemistry Research. 2012. Vol. 21. P. 3568-3580.
- Kinoshita K. et al. Inhibitory effect of some triterpenes from cacti on 32Pi-incorporation into phospholipids of HeLa cells promoted by 12-O-tetradecanoylphorbol-13-acetate // Phytomedicine. 1999. Vol. 6. P. 73-77.
- Lisiak N. et al. Biological activity of oleanolic acid derivatives HIMOXOL and Br-HIMOLID in breast cancer cells is mediated by ER and EGFR // International Journal of Molecular Sciences. 2023. Vol. 24. P. 5099.
- Luchnikova N.A. et al. Biotransformation of oleanolic acid using Rhodococcus rhodochrous IEGM 757 // Catalysts. 2022. Vol. 12. P. 1352.
- Luchnikova N.A., Grishko V.V., Ivshina I.B. Biotransformation of oleanane and ursane triterpenic acids // Molecules. 2020. Vol. 25. P. 5526.
- Ludwig B. et al. Whole-cell biotransformation of oleanolic acid by free and immobilized cells of Nocardia iowensis: Characterization of new metabolites // Engineering in Life Sciences. 2015. Vol. 15. P. 108115.
- Ma Y. et al. Microbial transformation of glycyrrhetinic acid and potent neural anti-inflammatory activity of the metabolites // Chinese Chemical Letters. 2017. Vol. 28. P. 1200-1204.
- Maatooq G.T. et al. Bioactive microbial metabolites from glycyrrhetinic acid // Phytochemistry. 2010. Vol. 71. P. 262-270.
- Mallavadhani U.V. et al. Synthesis and anti-cancer activity of some novel C-17 analogs of ursolic and oleanolic acids // Medicinal Chemistry Research. 2013. Vol. 22. P. 1263-1269.
- Martinez A. et al. Biotransformation of oleanolic and maslinic acids by Rhizomucor miehei // Phytochemistry. 2013. Vol. 94. P. 229-237.
- Paterson R.R.M. Fungi and fungal toxins as weapons // Mycological Research. 2006. Vol. 110. P. 10031010.
- Qin Y.J. et al. Biotransformation of glycyrrhetinic acid by Cunninghamella blakesleeana // Chinese Journal of Natural Medicines. 2010. Vol. 8. P. 373-381.
- Rakhimova M.B. et al. Effect of glycyrrhetic acid derivatives on regulation of thymocyte volume // Bulletin of Experimental Biology and Medicine. 2023. Vol. 175. P. 27-31.
- Sattarova I. et al. Research of antioxidant and prooxidant properties of glycyrrhetic acid derivatives // Science and Innovation. 2023. Vol. 2. P. 5-9.
- Shah S.A.A. et al. Microbial-catalyzed biotransformation of multifunctional triterpenoids derived from phytonutrients // International Journal of Molecular Sciences. 2014. Vol. 15. P. 12027-12060.
- Vishwakarma S. et al. In vitro and in silico studies of glycyrrhetinic acid derivatives as antitubercular agents // Letters in Drug Design & Discovery. 2022. Vol. 20. P. 479-487.
- Wang W.W. et al. Microbial hydroxylation and glycosylation of pentacyclic triterpenes as inhibitors on tissue factor procoagulant activity // Bioorganic and Medicinal Chemistry Letters. 2017. Vol. 27. P. 1026-1030.
- Wu S.B. et al. Cytotoxic triterpenoids and steroids from the bark of Melia azedarach // Planta Medica. 2011. Vol. 77. P. 922-928.
- Xu S.H., Zhang C. et al. Site-selective biotransformation of ursane triterpenes by Streptomyces griseus ATCC 13273 // RSC Advances. 2017. Vol. 7. P. 20754-20759.
- Xu S.H., Wang W.W. et al. Site-selective oxidation of unactivated C-H sp3 bonds of oleanane triterpenes by Streptomyces griseus ATCC 13273 // Tetrahedron. 2017. Vol. 73. P. 3086-3092.
- Xu S.H. et al. Application of tandem biotransformation for biosynthesis of new pentacyclic triterpenoid derivatives with neuroprotective effect // Bioorganic and Medicinal Chemistry Letters. 2020. Vol. 30. P. 126947.
- Yan S. et al. Microbial hydroxylation and glycosidation of oleanolic acid by Circinella muscae and their anti-inflammatory activities // Natural Product Research. 2018. Vol. 33. P. 1849-1855.
- Zhang J. et al. Novel biotransformation of pentacyclic triterpenoid acids by Nocardia sp. NRRL 5646 // Tetrahedron Letters. 2005. Vol. 46. P. 2337-2340.
- Zhang M. et al. Biotransformation of 18p-glycyrrhetinic acid by human intestinal fungus Aspergillus niger RG13B1 and the potential anti-inflammatory mechanism of its metabolites // Journal of Agricultural and Food Chemistry. 2022. Vol. 70. P. 15104-15115.
