Полевые сборы мелких млекопитающих на территории Малого Хингана и Буреинского хребта

Автор: Фрисман Л.В., Капитонова Л.В., Картавцева И.В., Шереметьева И.Н.
Журнал: Региональные проблемы @regionalnye-problemy
Рубрика: Биология
Статья в выпуске: 2 т.22, 2019 года.
Бесплатный доступ
Систематизированы результаты экспедиционных сборов на территории российской части Малого Хингана и на Буреинском хребте, выполненных с 2010 по 2018 годы. В результате проведенных полевых сборов обнаружено 11 видов грызунов. Представители родов Apodemus (A. peninsulae), Myodes (M. rutilus), Craceomys (C. rufocanus) Tamias (T. sibiricus) распространены от южной до северной точек рассматриваемой территории. Apodemus agrarius и два вида восточноазиатских полевок Alexandromys (A. fortis и A. maximowiszii) распространены в южной части Хингано-Буреинского нагорья. Северная граница ареала полевой мыши здесь представлена трансектой север Малого Хингана - юг Буреинского хребта. Эта граница совпадает с северным краем ареала дальневосточной (большой) полевки. Полевка Максимовича по Буреинскому хребту проникает значительно севернее. Домовая мышь, мышь-малютка и крыса-пасюк встречены в природных биотопах единичными экземплярами. Эворонская полевка обнаружена локально на севере рассматриваемой территории.
Малый хинган и буреинский хребты, видовое разнообразие, грызуны
Короткий адрес: https://sciup.org/143167176
IDR: 143167176 | УДК: 575.22:599.32(571.62) | DOI: 10.31433/2618-9593-2019-22-2-13-25
Field capture of small mammals in the northeast of Lesser Khingan and the Bureinsky range
The paper considers the results of expedition captures in Lesser Khingan and the Bureinsky range, for the period of 2010-2018. They have resulted in finding of eleven species of rodents. The representatives of the genera Apodemus (A. peninsulae), Myodes (M. rutilus), Craceomys (C. rufocanus), and Tamius (T. sibiricus) are widespread from the southern to the northern points of the territory. The Apodemus agrarius and two species of the East Asian voles (Alexandromys fortis and A. maximowiszii) are spread in the southern part of the Khingan-Bureya Highland. The northern boundary of the A. agrarius habitat stretches from the north of Lesser Khingan to the south of the Bureinsky Range. This boundary coincides with the northern edge of the A. fortis habitat. The A. maximowiszii moved the much further to the north along the Bureinsky range. Single specimen of the Mus musculus, Micromis minutes and Rattus norvegicus species occur in natural biotopes. The Alexandromys evoronensis was locally found in the north of the area.
Текст научной статьи Полевые сборы мелких млекопитающих на территории Малого Хингана и Буреинского хребта
Мелкие млекопитающие играют важную роль в экосистемах различного типа, являясь массовыми потребителями первичной продукции и прокормителями многих видов хищных животных. В связи с этим велико их значение для человека как вредителей сельскохозяйственных угодий и основы кормовой базы ряда охотничье-промыс-ловых видов. Практическая значимость изучения биоразнообразия и распространения видов мелких млекопитающих, особенно грызунов, кроме того, определяется их санитарно-эпидемиологическими характеристиками как носителей различающихся спектров природно-очаговых инфекционных заболеваний.
Во второй половине прошлого века были проведены обширные работы по изучению фауны мелких млекопитающих Буреинского хребта, в том числе в связи с анализом территорий, прилежащих к восточной зоне Байкало-Амурской магистрали [3, 11, 16, 17, 24]. Использование классических признаков для определения видового статуса таксономически сложных форм, таких как морфо- логически близкие виды серых полевок, приводило к неоднозначности в трактовке выявленного видового разнообразия и требовало привлечения дополнительных видоспецифичных маркеров. Происходящее в тот период бурное развитие генетических методов, таких как кариология, ал-лозимный анализ и молекулярно-генетические исследования ДНК, показало их перспективность как источников таксономически значимых маркеров межвидового уровня.
Высокая численность и быстрая смена поколений позволяют рассматривать некоторые виды грызунов в качестве высоко востребованных модельных объектов при проведении фундаментальных и прикладных исследований. Фундаментальное направление наших работ было определено эволюционно-генетическими задачами, находящимися в рамках исследования процессов видообразования, такими как генетическое разнообразие, внутривидовая и межвидовая подразделенность, генетическая изменчивость, структура краевых популяций. При полевых работах эволюционно-генетической направленности, проводи- мых на территории Среднего Приамурья, мы основное внимание уделяли природным популяциям модельных видов (серые и лесные полевки, восточноазиатская и полевая мыши) и интересным районам их обитания, при этом учитывая все попутно обнаруженное видовое разнообразие грызунов в каждом из исследованных локалитетов. В силу этого в ходе сбора материала для эволюционно-генетических исследований накапливались сведения о видовом составе родентофауны территории.
Еще одна часть наших полевых сборов в Среднем Приамурье определялась работами по исследованию родентофауны используемых и заброшенных сельскохозяйственных угодий территории Еврейской автономной области (далее – ЕАО), в основном локализованных на Среднеамурской (Амуро-Сунгарийской) низменности [21]. Для сравнения были проведены сборы в природных биотопах прилежащих низкогорий Малого Хингана.
Таким образом, в ходе сборов полевого материала нами накоплены сведения о видовом составе родентофауны хребтов Буреинский и Малый Хинган. Эти данные могут предоставить и уточнить информацию по современному биоразнообразию мелких млекопитающих природных территорий Среднего Приамурья, в настоящее время слабо затронутых давлением антропогенного пресса. Систематизация результатов экспедиционных сборов родентофауны на территории российской части Малого Хингана и Буреинского хребта, проводенных в полевые сезоны 2010–2018 гг., является целью настоящего исследования.
Материалы и методы
В полевые сезоны 2010–2014, 2016 и 2018 гг. обследовано тридцать четыре местообитания в двадцати локальностях на территории Малого Хингана и Буреинского хребта. Отработано 2736 ловушко-суток. Отловлено 447 экземпляров грызунов. Карта-схема сбора материала приведена на рис. Сбор материала проводился в теплое время года (июнь–октябрь) с помощью ловушек Шермана и ловушек Геро. Учет численности грызунов в рамках данной работы не проводился, так как при отлове ловушками Шермана не использовались специальные методы учета численности и соответствующие методики установки ловушек. При описании видового состава результаты отлова ловушками разного типа суммировались.
Для определения видовой принадлежности отловленных мелких млекопитающих использованы как классические морфологические признаки, так и таксономически значимые генетические характеристики. При определении видовой принадлежности серых полевок рода Alexandromys были использованы кариотипические и аллозимные характеристики [19, 20, 22], а также молекулярно-генетические характеристики митохондриальной ДНК [25, 26]. Для северной пищухи использованы молекулярно-генетические характеристики митохондриальной ДНК и хромосомные характеристики [7, 27]. Для восточноазиатских мышей были исследованы особенности числа и морфологии добавочных хромосом, характеризующих их кариотип [15]. При работе с лесными полевками наряду с морфологическими признаками (окрас шкурки, ширина резцов, зубы) рассматривались маркерные аллозимные характеристики, описанные нами ранее [18, 23]. Следует отметить, что на таксономическую значимость признака «ширина резцов» при разделении видов красной и красно-серой лесных полевок одному из соавторов любезно указал А.М. Долгих в устном сообщении. По нашим данным, этот признак удобен для разделения половозрелых животных и показывает некоторые затруднения в использовании при определении ювенильных экземпляров красно-серых полевок.
Результаты и обсуждение
В табл. приведены полученные данные по встречаемости мелких млекопитающих отряда Rodentia в исследованных локалитетах Буреин-ского нагорья, а именно на Буреинском хребте и по российской части хребта Малый Хинган. Последний рассматривается нами в трактовке А.Е. Криволуцкого [10] как расчлененное плоскогорье высотой 600–700 м, локализованное между рекой Амур, Зейско-Буреинской и Сренеамурской низменностями, а значит, включающий в себя возвышения Сутарского и Помпеевского хребтов. Северной границей Малого Хингана служит гребень водораздельного хребта между реками Бурея и Бира.
Самые южные сборы были проведены нами вдоль южной и юго-западной окраин российской части Малого Хингана в долине р. Амур от р. Столбуха (южная оконечность хребта) до р. Большая Солони (локалитеты 1-5 рис. и табл.). Здесь выявлено обитание восьми видов грызунов. Полевая мышь ( Apodemus agrarius Pallas, 1771), восточноазиатская мышь ( A. peninsulae Thomas, 1907), домовая мышь (Mus musculus L. ) , красно-серая лесная полевка ( Craceomys rufocanus ), красная лесная полевка ( Myodes. rutilus, Pallas, 1779), два вида серых полевок рода Alexandromys – полев-
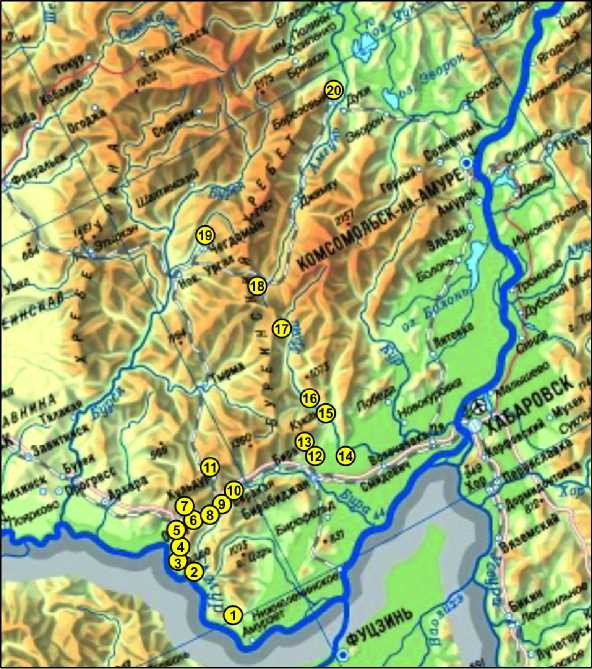
Рис. Карта-схема мест сбора материала.
Номера точек на карте соответствуют таковым в табл.
Fig. Map of the collection sites. Point numbers on the map correspond to those in Table ка Максимовича (A. maximowiszii Schrenk, 1859) и дальневосточная (большая) полевка (A. fortis Buchner, 1889) обнаружены по отловам. Визуально и по отловам наблюдался бурундук (Tamias si-biricus Laxmann, 1769).
Полевая мышь – наиболее часто встречающийся вид в открытых луговых биотопах, в том числе заросших кустарниками, также встречалась и на лесных участках вдоль дорог. Восточноазиатская мышь чаще отлавливалась в лесных биотопах, как смешанных лиственных долинных и приуроченных к сопкам, так и кедрово-широколиственных. В единичных случаях этот вид был обнаружен в приречных биотопах. Лесные полевки, представители родов Craceomys и Myodes, а также восточноазиатские серые полевки рода Alexandromys обнаружены в биотопах открытых пространств и на лесных опушках. Единственный экземпляр домовой мыши отловлен в окрестностях поселения Столбовое на открытом луговом пространстве (точка 1 на рис. и табл.). Возможно, что она появилась здесь в результате сезонной от- кочевки из строений поселения.
На северной части Малого Хингана отловы проводились в долине реки Хинган (вдоль трассы Облучье–Хинганск). В смешанном лиственном лесу на склоне сопки в окрестностях г. Облучье (локалитет 6) отловлена только восточноазиатская мышь, а бурундук был отмечен визуально . Рядом в распадке между сопками, заросшими кустарником и высоким разнотравьем, наряду с восточноазиатской мышью были обнаружены полевая мышь и красно-серая полевка. Дальневосточная полевка, полевая и восточноазиатская мыши отловлены на окраине г. Облучье на мокром лугу около р. Хинган. В смешанном лесу на склоне горы и на облесенных приречных террасах вдоль дороги Облучье–Хинганск (локалитет 7) в отловах преобладала восточноазиатская лесная мышь и встретилась красная полевка. Единственный экземпляр полевой мыши был отловлен здесь на разнотравном лугу, вероятно, используемом для покоса. Лесная мышь и красно-серая полевка преобладали в этом отлове.
Таблица
s
|
О |
СЧ |
ОО |
о |
О. |
40 |
||||
|
)S d> о о о со S § со и m |
СЧ со 1 Со £ |
7 ■S t: 1 ^ Ьс 8, £ а, |
О. со ^^ s 3 о 3 |
ОО Со 3 Со £ |
со Si, Со £ |
3 s 'с .8 3 3 |
£ |
m г| ■S t: 1 ^ £ 43 в |
.Г1 О! 8 § £ 8 о ^to |
|
в к Й о § ° а к |
СП СЧ Q ОО а 04 S Ln СО 4^ Е^ ^ |
ей о & S § § § 9 К О о о = & 2? “ ° ч ч- |
S о о ч® Й Сч м ОО м 40 2 ? 1 9 g fl О Ln 9 9 |
к & о CQ О А 3 О Ln и По СП ° СП о № । СП сч сч со । , § Ч- ^. Ч ч- ч- |
сЗ & ОО D СП 5 о S ч д о о 22 Ч « и ОО а S оо U ч- |
g Ч m м оо ел S и ” |
s' ч & м S s § ей 2 § S о m 2 S 2 S Р 3 Я СО О Q 'g СП § ’S о и 3 д ч О; — к Г| св Й S о В н s yi Ч О Св Ч Й Щ в ч 9 о S св У оо |
6 н & ^ 5 7 ч § ? о И ел ей ей И о Г - ООО |
|
|
ей и ей Й S § о |
5 о о . Ч о Й д ч о ч 2 ей О ГП и ч о к ч й ч к 8 9 2 2 ч g ч g & s g S ч ™ U о § Й g 6 ЙО Й ° S И) < Я 2 И И И Я |
& g 5 g 8 ^ Й S >3 9 m О И О V ^ 9 ^® m о S |
К & со со ^7 о g И К |
1 хЗ о к К со я 2 g и о -j c Г| “ 2 S 2 со 9 3о 4 » Щ К гм |
5 OJ оо о о< |
S ^ Ci СО а" й & о § g к о g § § S В о ц гм о I | о -ЗиЛ 9 g @ о щ 1 W сч |
|||
|
< к |
СП |
ч- |
LT) |
||||||
Продолжение таблицы Table continuation
|
ГЧ ГЧ |
04 |
LT) |
|||||
|
)й d> XO о о о со й й § со й и й m |
СЧ 53 Si, Со |
ип й е 3 К S3 be § g « si, X X £ £ § Чз Чз й О О Q |
т 1 so 4 -S .й ОД й> чз чз S о о 8 Si, S3, |
Ln Со , к S § 3 5 |
40 йГ е 3 В е с£ S3 Si, Со Со S S § О О Q и |
7 3 m § § « ^ ' ^S ■~ ^3 1 5 g ^5 й е g §, | ■2 о s 9 « |
40 Со Со Si, S3, |
|
в к Й р к к |
й й s й й а й и 8 5 м s к « в со а о & g “ оо g a g 2 9 ° 3 ^ В к а ° а m 2 । >s СО g \ S § S и М S , Я S LD Й Н й О U О о U 5? о й |
S й со й ы й и 9 >3 & й - а § о л tt m К s св о « g. и ж св Р- m |
_ о s к § К д О У & О К 9 R о о d^a в « ^g vg °° & u S Г4 И И s § § 8 2 и & й -В о U Щ й . г = со щ оо Г|^ и 2 к о ЙОЙО, йн 4 И тГ |
s В н S й G й о о к к од m & 40 § У >< О -н g, в и В м R о 5 5^$ ЙН КО S |
й И X S й о 1^ S g Н о 6У со § р “ О', |
й и й й m СО KN Н о U Ц ни g § о 2 о , Ч К оо X к |
й s g Й а И й 3 7 m 3 i S 1 S? и й СО й g S у 6 SO § В 7 X к ” |
|
й и й Й й й § о |
ж & . Q У g О С| |
О О О о |
2 й X И 1 о g g О г 1 о < "7 Щ оо ri |
й й й ° ё | § д о g rn И К И в 2 г 1 Ц д" ГП 2 О д в s Q У 7 о < Щ н О о-. |
ж Й Й о g сп И К и О ” В 2 Q 3 ^о < g,2 ^, Щ н О о-, |
||
|
40 |
ОО |
||||||
Продолжение таблицы Table continuation
|
3g g S' |
40 |
г- |
40 |
О |
ОО |
||
|
)S d> KO о о о со S § со Д и ЕД m |
и СП m ' о о 43 43 g £558 |
ио ио Со S с & о ■2 i g о е |
I Со ем §
Со Со Со О
^ ^ ^ ^ |
40 §
« ^ S. Ьо S Р & ® S ■2 43 Й g £ g g 8 |
ем — .У ем | « m 8 s Co Co ~ 0 43 43 u g g g g 8 S' S' r\ ~ U |
§ -2 S 2 s ^^ S t s S £ ^ s g 9 g 8 й ^ ^ и |
|
|
в к Й о s |
S bi s О U д д оо О СП Д g ° , О Г X О СО у й ио |
Д § л ° s и м И Й „ Й у л § S 5 т я ко § ° д ° ° о 2 о Ч к о S х ° д д о 0 в Й и So - S и Я К S Z О', 6 § |Bs о Н О а 5 s о X К S П К тГ |
СО д 6 & S ° )S ® О в; « В s 5 О « СО « |
а ж = о СО СО & СО О О д о s 8 22 ^^^ п & & О m = сг |
g § Я 0 а й § ° 0 c3 bi Д О й « § 0 ^ О g S 0 s a 8 « a G ® g S S g В VO G M g g m ° 2 О g m O £ S 0 vr> я 5 и VO ° Ц g 0 I ем о m & н m -e s 5 g ^ S н cn ко s S cn д И 0 о д н оо W О К О Д- |
св Л s к 5 У S S н 8 Я 3 о й Д S S у У а 3 s g „ s Oggs g § а Ч S 3 ч ’2 § § vo А s О 9 2 3 ® й ^ о S tn н о 5 3S й о о 2 ч ” &= § 8 i ^2 8 а 6 В в ° д & 2 | со ко д о ко д- |
|
|
Д ко и Д Й ед S bi |
,2 8 «к S bi О А С 3 ri О 1 и О', Щ О |
о < 0 |
5 oq 00 0 91 00 0 о< |
S х со д s а о >s 2 1 ° 3 S « ем VO 2 о А 2 |
Б ем и S ^ |
||
|
< К |
о |
||||||
Продолжение таблицы Table continuation
|
a 2 g S' я |
40 |
СП |
40 |
ОО |
9 |
|
|
)S d> KO о о о со S § со сЗ и m |
m vo В g m § « li | | < й § 3 3 Р £ S л л § о 3 5 5x38 S Я Я S |
ГЧ § 3 « ■S -2 s X Я О я |
40 Г- <3 S |в 1 ■Е । Л Si 1 || § its |
oq Со Q S § ОО § Si 1 || § its |
oq Со S' S |
Со е X S |
|
в к Й о к |
К К сЗ И S О = S н °^ О g ” В ц сп И 2 се — о у = о g Я X со си 5 а ОО О в |
S § ОО * 1Г) 2 СП К ел й 40 & ц о |
g ко 2 8 g « D СП § 40 & ^ СЗ СП К ел U й и )S со = 05 Н ОО О CV СП |
со S S 8 rn й, о^ СП й 5 м дУ в « В в Л о 5 * g |
в & ко & ° в о В Щ Щ В о g I оо И § » О си СП св 2 я я * а о о М Й СП СП К 3 2 2 сз 2 од X " ип 40 о X X о о К О ип ип |
§ § ° g g m сЗ s О О СП W П -я s й О 2 § о и = о В ОО й СО о 2 хс Я Я <г> |
|
сЗ и сЗ Й S |
а У сз Й X § « нс & О 3 g s м "-> о — VO X О ОО |
& & се д X ^ "-> о — О 8 Я & о X О ОО |
я ® & o' g S о в X в & | 5 в 5 g и Н & 5 7 vo g о X § об |
& со & X g В Св & со 2 .^с ^ g VO дЯ X vo об |
Рч Щ « Й & g ^ 2 « Я 5 Й ° rs am |
|
|
< К |
40 |
г- |
ОО |
|||
Продолжение таблицы Table continuation
|
3g g |
ОЧ |
ч5 |
|
LT) LT) |
LT) |
О |
ОО LT) |
||
|
)S о KO о о о со S § со сЗ и m |
О |
СЧ со ГМ £ S й s ^§2^2 £ ° g |
Со s й S 1 6 § S § S 4 5 s' о s ■2 11 § |
со Со Со S й |
40 1 Со ^ Co 11 L« 3- St |
co |
со S4 Со |
со е 5 ^ X 2 t6 |
Со m | К § s 11 u bo S С РЗ 1 Р ! ■§ S | ^ 1 <1 tS |
|
в к Й о м м w i |
О 5 в со S н о u К СТ ед и а В о В о ^ Ц g m О и „ s « у m U ® А СМ КО о >, ID g ^ S ’g к СП g м к 5 5 | и s * У у g и и ^ В о и о g й ^.Д | | m 2 2 СО ^ U^ |
со , S & О о с со в ^ о & ст к ° а гм $5 У Д м ч д о § g з СП о й й m и и u U « м В % * ” § £ К ГП М Q » Ч ^ сс> |
В О н в g а О со о g 8 S | £ ^ ? g u и о-, о К ю см _ И s о Mog о у ” в § ^ к м О g 'О S н m г s У 5 о » - с 'л |
м О О м о ч о « м щ щ § о м 2 2 5 у о ^ ^ PQ м о Е; оо й о м Й О и g В О & ^ ugg см У ^ ^ СП В m ’В )Н г , М g М 4, ugg to W тГ s ^ 3 в 3 U3 о х СО Г4 СО Щ о О ОО Щ а к ко in |
2 у a ^ КО о R Ot )S о О О rc ^ |
в u a u ^ cq о a m О . Щ 4 Q = о в 9 >4 Й о Bom о у U U ОО св м с® ° MD U о к> g « го о Д К К m |
2 к ОО м < 5 св 31" U S «и |
ко КО щ в а ^ щ бг СО S У и и и -m m св аК" 2 св он О и 6 И w |
|
|
сЗ ко и сЗ Й S й |
А X a ci |
40 о ОО о |
40 О О об ОО о |
& М М & W 2 g М см g о О о g ^ §с U ^ ГО ^ О сб |
ОО О Оч О |
||||
|
< К |
ОЧ |
О |
|||||||
В хвойно-лиственных лесах вдоль трассы между населенным пунктом Известковый и городом Облучье (локалитеты 8-9) наиболее часто отлавливалась восточноазиатская лесная мышь, встреченная здесь одна либо совместно с лесными полевками и бурундуком.
На опушке мелколиственного леса и вторичных лугах по сопкам в окрестностях поселка Известковый (локалитет 10) в отловах преобладали полевая мышь и полевка Максимовича: 34 и 17 экземпляров соответственно. Также здесь были отловлены восточноазиатская мышь и бурундук.
На Малом Хингане в сборах, приуроченных к долине р. Кульдур в окрестностях пос. Кульдур (локалитет 11), обнаружено 7 видов грызунов ( T. sibiricus , A. peninsulae , A. agrarius , M. rutilus , С. rufocanus , A. fortis и А. maximowiszii ). В зарослях кустарников с вейниково-осоковым кочкарником вдоль ручья на территории санатория Кульдур обнаружены восточноазиатская лесная мышь и красно-серая полевка, при этом в отловах здесь преобладала полевая мышь . На разнотравном лугу в прибрежной кустарниковой зоне в долине р. Кульдур были отловлены восточноазиатская лесная мышь совместно с лесными полевками и восточноазиатскими серыми полевками. Генетическое типирование восточноазиатских полевок показало присутствие здесь в разные годы одного из двух видов: полевка Максимовича в отловах 2010 г. [20] и большая полевка в отловах 2011 г. (определена на основании кариотипирования К.В. Коробицыной).
Следующий район сборов (локалитет 12) расположен на юго-западной оконечности Буре-инского хребта, в белоберезовой релке и на переходе от смешанного лиственного леса к открытым лугово-болотным пространствам. Здесь было отловлено 5 видов грызунов – Tamiаs sibiricus, Apodemus peninsulae, Myodes rutilus, Сraceomys. rufo-canus и Alexandromys maximowiszii. В сухом лесу в отловах преобладала M. rutilus . В зеленомошном смешанном лесу (локалитет 13) были отловлены эти же 5 видов грызунов – T. sibiricus, A. peninsulae, M. rutilus, С. rufocanus и A. maximowiszii. При этом наиболее часто в сборах оказывалась восточноазиатская мышь. Здесь же был добыт представитель мелких млекопитающих из отряда зайцеобразных – северная пищуха ( Oсhotona hуperborea Pallas, 1811) [27]. Южнее, в охранной зоне кластера Бастак заповедника Бастак в лиственничном редколесье на разнотравном лугу, была обнаружена мышь-малютка ( Micromys minu-tus , Pallas, 1771) .
В юго-восточном районе Буреинского хребта сборы проведены в долине р. Урми между поселками Кукан и Догордон (локалитеты 15-17). Кроме того, проведены сборы южнее – в долине р. Аур, в окрестностях пос. Томское (локали-тет 14). Было отловлено 6 видов грызунов: полевая и восточноазиатская мыши, красная и красно-серая лесные полевки, восточноазиатские серые полевки – большая и Максимовича, визуально был отмечен бурундук. Полевая мышь и большая полевка встречены только в окрестностях пос. Томское. Полевая мышь здесь преобладала в отловах. Этот участок представляет типичное разнообразие видов мелких млекопитающих открытых заброшенных антропогенно-трансформированных районов. В остальных местообитаниях (локали-теты 15-17), расположенных вдали от поселков, в долинном лесу и на небольших участках луга с кочкарником в отловах чаще преобладала восточноазиатская мышь, которая встречалась здесь наряду с красной и красно-серой полевками, а в локалитете 15 еще и с полевкой Максимовича.
Самые северные точки наших сборов на западе Буреинского хребта локализованы в верховьях р. Бурея, в долинах ее притоков Сололи и Чег-домын в окрестностях одноименных населенных пунктов, находящихся на трассе БАМ (локалитеты 18-19). Здесь были отловлены 5 видов грызунов: T. sibiricus , A. peninsulae , M. rutilus , C. rufocanus . Генетическое типирование впервые обнаружило в окрестностях с. Чегдомын эворонскую полевку ( Alexandromys evoronensis Kowalskaia et Sokolov, 1980) [25]. На лесных участках (лиственница, осина, много валежника) с небольшими марями преобладала красно-серая полевка, а в долинном лесу на осоковом лугу с кустарниками – восточноазиатская мышь. Также отлавливались единичные экземпляры красной лесной и эворонской полевок. Эворонская полевка обнаружена и на покосе, на разнотравном лугу. Обращает на себя внимание большое количество бурундуков (10 особей из 36 экземпляров грызунов), отловленных в окрестностях г. Чегдомына.
Самые северные сборы на востоке Буреин-ского хребта были проведены в окрестности пос. Березовый (локалитет 20). В смешанном долинном лесу недалеко от р. Амгунь были добыты восточноазиатская мышь, красно-серая и красная полевки. В зарослях кустарников вдоль берега реки Вели, наряду с этими видами, были отловлены бурундук и крыса (Rattus norvegicus Berkenhout, 1769). В окрестностях пос. Березовый найдены останки ежа (Erinaceus), представителя еще од- ного отряда мелких млекопитающих – насекомоядные. Видовая принадлежность не определена. Следует добавить, что другой представитель этого отряда (Crocidura lasiura) была отловлена нами на Малом Хингане (локалитеты 1, 5) и на юго-западе Буреинского хребта (локалитеты 14-17). Последние точки сбора находятся севернее представленной в литературе границы ареала этого вида [6].
На Хингано-Буреинском нагорье обнаружено 11 видов грызунов, принадлежащих к 7-ми родам. Из них домовая мышь, мышь-малютка и крыса-пасюк встречены в природных биотопах единичными экземплярами, а эворонская полевка – локально. Подтверждены данные В.И. Волкова с соавторами [3] и В.Т. Тагировой [16, 17] о широком распространении представителей родов лесных и полевых мышей ( Apodemus ), лесных полевок ( Craceomys, Myodes ), восточноазиатских серых полевок ( Alexandromys ) и бурундуков (Tamias) по рассматриваемой территории.
Согласно литературным данным, в Среднем Приамурье восточноазиатская лесная мышь ( A. peninsulae ) встречается в разнообразных лесных биотопах, отдавая предпочтение кедрово-широколиственным, а при высокой численности мелколиственным лесам, при этом данный вид избегает ельников и листенничников [8]. Практически во всех рассматриваемых в настоящей работе локалитетах A. peninsulae был отмечен (в 22 местообитаниях), при этом в 12 местообитаниях преобладал в отловах.
Полевая мышь (Apodemus agrarius) на юге Дальнего Востока считается доминантным видом безлесных территорий [8], при этом оптимум ареала популяций приурочен к сельскохозяйственным землям на месте неморальной растительности [9]. На территории Амуро-Сунгарийской низменности полевая мышь распространена в открытых природных и антропогенно-трансформированных биотопах, доминирует в сельскохозяйственных угодьях [12, 21]. Именно такими являются локалитеты 4, 10 и 14 (табл.), подвергающиеся в прошлом или в настоящем жесткой антропогенной трансформации. По мнению В.И. Волкова с соавторами [4], расселение полевой мыши на дальневосточном участке ареала в пределах нашей страны началось с XVIII столетия, оно вызвано освоением лесов и развитием сельского хозяйства. По мнению В.А. Костенко [8], присутствие этого вида в лесах является следствием их нарушенности. В локалите-тах рассматриваемого горного комплекса полевая мышь появляется лишь на антропогенно измененных участках и единично на открытых участках вдоль дорог, попадая сюда благодаря деятельности человека. Судя по нашим сборам, северная граница ареала этого вида на рассматриваемой территории представлена трансектой Облучье–Кульдур–Том-ское (локалитеты сбора 6, 7, 11, 14). Н.С. Беляева [2] наблюдала полевую мышь севернее поселка Кульдур, в окрестностях пос. Тырма Верхнебуре-инского района Хабаровского края.
Вышеуказанная трансекта совпадает с северным краем ареала еще одного вида – дальневосточной (большой) полевки. Этот вид является типичным представителем пойменных луговых и переувлажненных луговых участков лесостепной зоны в пределах Амуро-Сунгарийской низменности, по долинам рек может проникать в долинные широколиственные леса. При этом по направлению к северу встречаемость вида сильно снижается. Следует отметить, что на исследованной территории дальневосточная полевка обитает симпатрически с еще одним представителем восточно-азиатских серых полевок – полевкой Максимовича. Этот вид так же широко распространен в пределах Амуро-Сунгарийской низменности, однако, по-видимому, проникает севернее, чем большая полевка. Так, если в окрестностях пос. Куль-дур мы обнаружили оба вида, в окрестностях пос. Томское только большую полевку (локалитет 14), то на территории кластера «Бастак» и около его юго-западной оконечности (локалитеты 12-13), а также в долине р. Урми (локалитет 15) обнаружена только полевка Максимовича. Стоит отметить, что дальневосточная полевка и полевка Максимовича могут встречаться симбиотопически, при этом они могут отлавливаться совместно [20] или сменять друг друга в разные годы (локалитеты 4, 11 настоящего исследования).
Ранее, при описании видового разнообразия грызунов восточной зоны Байкало-Амурской магистрали, серые полевки из окрестностей поселка Чегдомын были определены как полевка Максимовича [1, 11] либо как дальневосточная (большая) полевка [8, 24]. Проводимые нами полевые сборы и их анализ с привлечением генетических методов идентификации видов позволяет уточнить и дополнить картину биоразнообразия Буреинского хребта. Привлечение генетических признаков в качестве таксономических характеристик позволило обнаружить в Верхнебуреинской впадине эворонскую полевку [25]. Находка эво-ронской полевки в данном регионе и западнее – на северо-востоке Верхнезейской равнины в долине реки Арги в Амурской области [26, 28] меняет представление о распространении этого вида. Его ареал, прежде рассматриваемый как узкий – приуроченный к Эворон-Чукчагирской впадине, приобретает неопределенную пока протяженность на запад, по меньшей мере до верховьев реки Зея. Судя по материалам проведенного в 1964–1976 гг. подробного исследования видового состава грызунов восточной части БАМа [3], серые полевки здесь чрезвычайно редки. Их малочисленность вытекает и из наших данных. На 1085 ловуш-ко-суток в трех северных локалитетах нами были отловлены только четыре особи данного вида. Это значит, что их поселения должны быть изолированы друг от друга. В пределах рассматриваемой нами Хингано-Буреинской горной системы остается под вопросом видовая принадлежность представителей рода Alexandromys на неисследованной территории, расположенной южнее железнодорожной ветки восточного БАМа и севернее линии Облучье–Кукан. Иными словами, пока нет полной ясности, насколько далеко полевка Максимовича (так же, как и большая полевка) проникает на север, а эворонская полевка на юг. Для эворон-ской полевки ареал только начинает определяться.
Наши полевые сборы на территории восточного БАМа проведены в окрестностях ныне существующих населенных пунктов, в долинных биотопах, где распространены вторичные лесные группировки (мелколиственные, преимущественно березово-осиновые леса, кустарники, вырубки, мари). Наиболее часто встречаемыми видами грызунов здесь являются лесные полевки и восточно-азиатская лесная мышь. Эти виды распространены и в южной части рассматриваемой нами территории Хингано-Буреинского нагорья, где кроме них широко представлены полевая мышь, полевка Максимовича и большая полевка. Перечисленные шесть видов несомненно приспособились к существующим условиям антропогенного давления. Нередко антропогенная трансформация (главным образом сельскохозяйственная деятельность) поставляет грызунам дополнительную кормовую базу и позволяет расширять ареалы. Особенно ярко это демонстрирует пример полевой мыши. В литературе упоминается о возможности завоза человеком данного вида в нетипичные местообитания, такие как остров Кунашир [13] и Магаданская область [5, 12]. Этот вид в настоящее время, несомненно, расширяет ареал, хотя пока не обнаружен в сборах на территории востока и севера Буреинского хребта.
Расширение кормовой базы на фоне высокой плодовитости определяет возможность резкого увеличения численности мелких мле- копитающих, что может привести к вспышкам природно-очаговых инфекций. Отслеживание и мониторинг их видового разнообразия в условиях изменяющегося антропогенного давления на окружающую природную среду является важным компонентом экологических исследований.
Первые этапы наших экспедиционных исследований на территории Малого Хингана были проведены совместно с К.В. Коробицыной, чей профессионализм полевика и кариолога мы глубоко чтим. Выражаем искреннюю признательность О.Л. Ревуцкой, Д.И. Шишацкой (ИКАРП ДВО РАН) и Т.В. Васильевой (ФНЦ биоразнообразия наземной биоты Восточной Азии ДВО РАН) за помощь в проведении полевых работ.
Работа выполнена в рамках государственного задания ИКАРП ДВО РАН и при частичной финансовой поддержке программы ДВО РАН «Дальний восток», грант № 18-5-013.
Список литературы Полевые сборы мелких млекопитающих на территории Малого Хингана и Буреинского хребта
- Беляева Н.С. Мышевидные грызуны южной части Хабаровского края//Известия Иркутского государственного научного-исследовательского противочумного института Сибири и Дальнего Востока. Иркутск: Кн. изд-во, 1963. Т. 25. С. 304-308
- Беляева Н.С. Мышевидные грызуны южной части Хабаровского края и динамика их численности//Вопросы географии Дальнего Востока. Владивосток: Дальневост. кн. изд-во, 1965. Сб. 7. С. 252-267
- Волков В.И., Долгих А.М., Кацко В.И., Зарубина В.И., Прасолова Н.Н. Эктопаразиты мелких млекопитающих восточной части БАМа//Паразитология. 1978. Т. 12, № 6. С. 529-537
- Волков В.И., Черных П.А., Долгих А.М., Яковлева Т.В. Распространение и основные черты экологии полевой мыши (Apodemus agrarius) в Приамурье//Зоологический журнал. 1979. Т. 18, № 8. С. 1202-1212
- Докучаев Н.Е., Поспехов В.В., Лазуткин А.Н. Нежелательная интродукция полевой мыши в Магаданскую область//Колымские вести. 2001. № 14. С. 19-20
- Зайцев М.В., Войта Л.П., Шефтель Б.И. Млекопитаюшие фауны России и сопредельных стран. Насекомоядные. СПб.: Наука, 2014. 389 с
- Картавцева И.В., Шереметьева И.Н., Гуськов В.Ю., Вакурин А.А., Кумакшева Е.В., Фрисман Л.В. К уточнению таксономического положения северной пищухи Ochotona hyperborea Сихотэ-Алиня//Вестник ДВО РАН. 2014. № 2. С. 79-83
- Костенко В.А. Грызуны (Rodentia) Дальнего Востока России. Владивосток: Дальнаука, 2000. 210 с
- Костенко В.А., Нестеренко В.А. Особенности территориального распределения грызунов и его динамики в Южном Приморье//Териологические исследования на юге Дальнего Востока. Владивосток: ДВО АН СССР, 1989. C. 54-71
- Криволуцкий А.Е. Амурско-Приморская страна//Физико-географическое районирование СССР. М.: Изд-во Моск. ун-та, 1968. C. 503-542
- Кузиков И.В., Ковалевский Ю.В., Липаев В.М., Чипанин В.И., Митейко В.Ф. и др. Мелкие млекопитающие Амуро-Буреинского участка БАМа//Бюллетень Московского Общества испытателей природы. Отд. биол. 1979. Т. 84, № 2. С. 3-14
- Лапин А.С. Мелкие млекопитающие южной части Хабаровского края и Еврейской автономной области (фауна, экология, эпизоотологическое значение: автореф. дисс. … канд. биолог. наук. Хабаровск, 2013. 24 с
- Межжерин С.В. Полевая мышь Apodemus agrarius Pall. (Rodentia, Muridae) на о. Кунашир//Вестник зоологии. 1992. № 3. С. 71
- Переверзева В.В., Примак А.А., Павленко М.В., Докучаев Н.Е., Евдокимова А.А. Генетические особенности и возможные источники формирования изолированных популяций полевой мыши (Apodemus agrarius Pallas 1771) на территории Магаданской области//Российский журнал биологических инвазий. 2016. № 4. С. 93-109
- Рослик Г.В., Картавцева И.В., Фрисман Л.В., Горобейко У.В. Сравнительное исследование морфотипов В-хромосом восточноазиатской мыши (Apodemus peninsulae) Приамурья//Региональные проблемы. 2016. Т. 19, № 3. С. 113-122
- Тагирова В.Т. Результаты учета грызунов в некоторых ландшафтах Приамурья//Численность грызунов на Дальнем Востоке СССР в 1982 г. и ее прогноз на 1983 г. Владивосток: БПИ ДВНЦ АН СССР, 1983. С. 43-44
- Тагирова В.Т. Наземные позвоночные среднего и нижнего Приамурья (фауна, зоогеография, проблемы охраны и рационального использования): дис. в виде науч. докл. д-ра биол. наук/Ин-т проблем экологии и эволюции им. А.Н. Северцова РАН. М., 1998. 48 с
- Фрисман Л.В., Картавцева И.В, Павленко М.В., Костенко В.А., Сузуки Н., Иваса М., Накато К., Чернявский Ф. Геногеографическая изменчивость и генетическая дифференциация лесных полевок рода Clethrionomys (Rodentia, Cricetidae) Приохотья//Генетика. 2002. Т. 38, № 5. С. 655-664
- Фрисман Л.В., Коробицына, К.В. Картавцева И.В., Шереметьева И.Н., Войта Л.Л. Серые полевки (MICROTUS Shrank, 1798) Дальнего Востока России: аллозимная и кариологическая дивергенция//Генетика 2009. Т. 45, № 6. С. 804-812
- Фрисман Л.В., Картавцева И.В., Капитонова Л.В., Высочина Н. П., Рябкова А.В. Генетическое исследование серых полевок рода Microtus территории Еврейской автономной области//Региональные проблемы. 2011. Т. 14, № 2. С. 70-77
- Фрисман Л.В., Капитонова Л.В., Поляков А.В. Родентофауна Среднеамурской низменности и прилежащих низкогорий//Региональные проблемы. 2013. Т. 16, № 2. С. 47-53
- Фрисман Л.В., Картавцева И.В., Шереметьева И.Н., Павленко М.В., Кораблев В.П. Аллозимная дифференциация и кариотипические особенности восточноазиатских полевок Дальнего Востока России//Вecтник СВНЦ ДВО РАН. 2016. № 3. С. 93-103
- Фрисман Л.В., Картавцева И.В., Павленко М.В., Капитонова Л.В. Геногеографическая изменчивость гемоглобина у красной полевки (Мyodes rutilus Pallas, 1779) юга Дальнего Востока России//Региональные проблемы. 2016. Т. 19, № 3. С. 123-130
- Хамаганов С.А. Материалы по грызунам и их экто-паразитам северных районов Хабаровского края//Вопросы географии Дальнего Востока. Хабаровск: Дальневост. гос. изд-во, 1973. Сб. 11. С. 167-172
- Шереметьева И.Н., Картавцева И.В., Васильева Т.В., Фрисман Л.В. Серые полевки рода Alexandromys из Верхнебуреинской впадины//Зоологический журнал. 2016. Т. 95, № 5. С. 597-603
- Шереметьева И.Н., Картавцева И.В., Васильева Т.В. Обитает ли эворонская полевка (Alexandromys evoronensis) на северо-востоке Верхнезейской равнины?//Зоологический журнал. 2017. Т. 96, № 4. С. 477-484
- Kartavtseva I.V., Vakurin A.A., Lissovsky A.A., Frisman L.V. Cytogenetic characteristics of pikas Ochotona hyperborea and O. mantchurica (Lagomorpha, Mammalia) in Amur River Basin//Russian Journal of Theriology. 2016. Vol. 15, N 2. Р. 84-90
- Sheremetyeva I. N., Kartavtseva I.V., Vasiljeva T. N. Frisman L. Voles of the Genus Alexandromys from the Verkhnebureinskaya Depression//Biology Bulletin. 2017. Vol. 44, N 7. P. 813-819
