Полиамины как регуляторы биоплёнкообразования природных изолятов Escherichia coli с разной степенью устойчивости к фторхинолонам

Автор: Нестерова Лариса Юрьевна, Караваева Елена Анатольевна, Ткаченко Александр Георгиевич
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 2, 2011 года.
Бесплатный доступ
На основе анализа природных штаммов Escherichia coli показано, что микроорганизмы, устойчивые к действию фторхинолонов, обладают повышенной способностью к биоплёнкообразованию и высокой активностью орнитиндекарбоксилазы - ключевого фермента системы синтеза полиаминов. Установлено, что сублетальные концентрации фторхинолонов стимулируют биоплёнкообразование. Полиамины путресцин и спермидин оказывают стимулирующее действие на способность к биоплёнкообразованию у штаммов с разной степенью устойчивости к фторхинолонам как в присутствии антибиотика, так и в его отсутствие.
Биоплёнкообразование, антибиотикорезистентность, полиамины, орнитиндекарбоксилаза, левофлоксацин
Короткий адрес: https://sciup.org/147204545
IDR: 147204545 | УДК: 579.22
Polyamines as regulators of biofilm formation in Escherichia coli isolates with different levels of fluoroquinolone resistance
Fluoroquinolone-resistant Escherichia coli isolates were shown to have an elevated ability to form biofilms and possess by increased activity of ornithine decarboxylase, the key enzyme of polyamine synthesis. Upregulation of biofilm formation under sublethal fluoroquinolone treatment was observed. Putrescine and spermidine were found to have positive effect on biofilm formation in strains with different levels of levofloxacin resistance in both the presence of this antibiotic and its absence.
Текст научной статьи Полиамины как регуляторы биоплёнкообразования природных изолятов Escherichia coli с разной степенью устойчивости к фторхинолонам
Образование биопленок микроорганизмами представляет собой одну из основных проблем не только в области медицины, но и в экономической сфере, что связано с большим материальным уроном, наносимым обрастанием внутренней поверхности трубопроводов различного назначения.
Ранее показано, что в естественной среде обитания микроорганизмы часто образуют биоплёнки, которые представляют собой одну из наиболее распространенных форм их существования в природе. В структуре биоплёнок бактерии, находясь в состоянии поверхностной или межклеточной адгезии, заключены в синтезированный ими полимерный матрикс и по сравнению с планктонными клетками характеризуются специфичными физиологическими и метаболическими свойствами, обусловленными изменением их генной экспрессии. (Pratt, Kolter, 1999; Landini, 2009). Существование в виде биоплёнки даёт микроорганизмам ряд преимуществ, основным из которых является повышенная устойчивость к различным неблагоприятным воздействиям. Чувствительность бактерий, включенных в структуру биоплёнки, к антибактериальным препаратам может снижаться в 10–1000 раз по сравнению с планктонными формами (Naves et al., 2008), что создает трудности в терапии инфекционных заболеваний. Известно, что возбудителями более 60% внутрибольничных инфекций являются микроорга- низмы, растущие в виде биоплёнок (Davey, O’Toole, 2000). Однако, несмотря на интенсивные исследования в области структуры биоплёнкообразования, проблема регуляции этого процесса до сих пор остаётся малоизученной. Экспериментально доказано, что неблагоприятные внешние воздействия различной природы могут индуцировать и поддерживать образование биоплёнок (Landini, 2009), установлена роль многочисленных внутриклеточных факторов в их биогенезе, что указывает на сложность процесса биопленкообразования (Ghannoum, O’Toole, 2004). Показано, что в формировании биопленок существенную роль играют различные системы общей клеточной адаптации (Beloin, Roux, Ghigo, 2008).
Полученные в последнее время данные свидетельствуют о том, что существенную роль в адаптации микроорганизмов к различным видам стресса играют полиамины, универсальные клеточные регуляторы, присутствующие в клетках всех живых организмов (Tkachenko, Nesterova, Pshenichnov, 2001; Ткаченко, Нестерова, 2003; Ткаченко, Пожидаева, Шумков, 2006). Основными представителями соединений этого класса у прокариот являются аминосодержащие алифатические углеводороды путресцин и спермидин (Igarashi, Kashivagi, 2000). Ранее нами показано, что в ответ на воздействие фторхинолоновых антибиотиков в клетках E. coli происходит рост активности ферментов синтеза
полиаминов, что приводит к внутриклеточному накоплению этих соединений, где они играют роль фактора адаптации (Ахова, Ткаченко, 2009).
В связи с этим значительный интерес представляет изучение распространенности этого механизма адаптации среди природных штаммов микроорганизмов, что могло бы прояснить вопрос о его селективной значимости как признака антибиотико-устойчивости и повлиять на выбор стратегии усиления действия антибиотиков.
Исходя из этого, целью настоящей работы является изучение активности полиаминсинтезирующей системы в связи со способностью к образованию биоплёнок у природных штаммов Escherichia coli , выделенных из различных источников и обладающих разной степенью устойчивости к фторхиноло-ну левофлоксацину. Работа направлена также на исследование возможной роли полиаминов в механизме биопленкообразования у E. coli как в обычных условиях, так и при сублетальных воздействиях антибиотика.
Материалы и методы
Культивирование микроорганизмов. Природные штаммы E.coli сохраняли на скошенном агаре. При подготовке экспериментов штаммы высевали на пробирки, содержащие 5мл LB бульона и культивировали в термостате при 37°С. Затем полученную культуру доводили до оптической плотности (OD) 0.1 при λ=625 нм, готовили соответствующие методу разведения и инокулировали в 96-луночные иммунологические планшеты, после чего культивировали при 37°С.
Определение антибиотикочувствительности . Антибиотикочувствительность штаммов оценивали по значению минимальной подавляющей концентрации (МПК), которую определяли методом двукратных серийных разведений (Методические указания МУК 4.2.1890-04) с использованием иммунологических планшетов.
Определение ферментативной активности. Активность орнитиндекарбоксилазы определяли по количеству образующегося конечного продукта путресцина в единицу времени. Для этого исходную культуру (OD625=0.1) в объёме 50 мкл вносили в 96-луночные иммунологические планшеты, содержащие в лунках по 150 мкл LB бульона и культивировали при 37°С 4 ч. Оптическую плотность (OD600) контролировали на мультимодальном планшетном ридере Infinite M200 (Tecan, Швейцария). Клетки осаждали центрифугированием планшетных культур в течение 15 мин. при 3700 об/мин и температуре 0 ° С на центифуге 5804 R (Eppendorf, Германия), после чего отмывали физиологическим раствором. Ресуспендированные клетки разрушали ультразвуком (20 кГц, 130 Вт, 2 раза по 20 сек.) на ультразвуковом дезинтеграторе (Cole
Parmer, США), обломки клеток осаждали центрифугированием (20 мин., 3700 об/мин). Надосадочную жидкость использовали для проведения ферментативной реакции. Инкубационная смесь включала 100 мМ буфера ТРИС HCl (рН 8.0), 0.04 мМ пиридоксальфосфата, 1 мМ дитиотрейтола, 10 мМ орнитина и 50 мкл супернатанта, в конечном объеме 0.2 мл. Реакцию запускали добавлением субстрата, смесь инкубировали в течение 60 мин. при 37 ° С, реакцию останавливали внесением 50 мкл хлорной кислоты.
Определение содержания путресцина проводили методом ВЭЖХ его дансилированного производного. Хроматографическая система включала хроматограф Shimadzu LC-20A (Shimadzu, Япония); колонку Luna С18(2) размером 250 мм × 4.6 мм, 5 мкм (Phenomenex, США), предколонку С18 Securityguard, 4 мм × 3 мм, 5 мкм (Phenomenex, США); флуоресцентный детектор RF-10A XL (Shimadzu, Япония). Длины волн экстинкции и эмиссии устанавливали на 400 и 516 нм, соответственно. В качестве мобильной фазы использовали смесь воды и ацетонитрила, которую подавали на колонку со скоростью 1 мл/мин в линейном градиенте концентрации ацетонитрила от 40 до 100% в течение 10 мин. с последующим уравновешиванием в течение 10 мин. 40%-ным ацетонитрилом.
Определение количества белка проводили методом Лоури.
Биоплёнкообразование. С целью определения способности E. coli к биоплёнкообразованию культуру выращивали на синтетической глюкозо-минеральной среде М-9. В экспериментах по изучению влияния полиаминов на биопленкообразование в качестве инокулята использовали клетки, предварительно истощенные по эндогенным полиаминам с помощью 3-кратных пересевов на среде М-9. Планктонные клетки после культивирования удаляли из планшетов вместе со средой и лунки дважды отмывали 200 мкл дистиллированной воды. Планшеты подсушивали в течение 20 мин., после чего в каждую лунку добавляли по 150 мкл 0.1% генциан-виолета и выдерживали в течение 30 мин. при комнатной температуре. Краситель удаляли и лунки 5кратно промывали (добавление в лунки 200 мкл дистиллированной воды, встряхивание в течение 5 сек.). С целью стандартизации обработки все процедуры по промыванию планшетов проводили автоматически, в микропланшетном ридере. Планшеты высушивали, краситель экстрагировали абсолютным этанолом, после чего измеряли оптическую плотность (OD570). Измерения проводили в 16-кратной повторности. Способность к биоплёнкообразова-нию подсчитывали с использованием следующего уравнения: Биоплёнкообразование = (A – B) / C, где А – OD570 в опытной лунке; В – OD570 в контроль- ной лунке; C – OD625 культуры клеток (Naves et al., 2008).
Статистическая обработка результатов исследования проведена с использованием пакета стандартных программ Statistica 6.0 (StatSoft, Inc., 2001). На рисунках приведены данные из серии однотипных экспериментов (не менее трех), представленные в виде медианы с указанием величины 25 и 75 процентилей. Оценка статистической значимости различий произведена с использованием непараметрического критерия Манна-Уитни. Различия считали значимыми при р ≤ 0.05.
Результаты и их обсуждение
В качестве объекта исследования использованы природные штаммы E. coli , изолированные на птицеводческих предприятиях Пермского края. Выбор источника штаммов обусловлен широким использованием антибиотиков, в том числе фтор-хинолового ряда, для лечения и профилактики бактериальных инфекций на предприятиях промышленного птицеводства (Кузнецова и др., 2010). Систематическое использование летальных, сублетальных и суббактериостатических концентраций антибактериальных препаратов является селективным фактором для отбора среди естественной комменсальной микрофлоры птиц антибиотикорезистентных форм микроорганизмов с разным уровнем устойчивости.
По величине минимальной подавляющей концентрации (МПК) левофлоксацина (ЛФЦ) 74 штамма E. сoli были разделены на 3 группы: 1 – менее 2 мкг/мл; 2 – в диапазоне от 2 до 8 мкг/мл; 3 – более 8 мкг/мл. Это распределение соответствует группам чувствительных, переходных и устойчивых штаммов по классификации, применяемой в клинической практике (Методические указания …, 2004).
Предварительный анализ 216 природных штаммов Е. coli , выделенных из различных источников, показал, что минимальная подавляющая концентрация левофлоксацина для большинства штаммов, не обладающих резистентностью к фторхинолоновым антибиотикам, составляет 0.038 мкг/мл. Группа из 45 штаммов, имеющих это значение МПК, использована в настоящей работе в качестве контрольной.
Анализ экспериментальных данных показал, что штаммы с высоким уровнем антибиотикорези-стентности к левофлоксацину обладают повышенной способностью к биоплёнкообразованию (рис. 1). Это согласуется с данными о более высокой устойчивости клеток в составе биоплёнок к действию антибиотиков (Naves et al., 2008). Биоплёнкообразо-вание, наряду с другими неспецифическими механизмами, такими как ограничение транспорта антибиотиков в клетку, их активный выброс, общая стрессорная адаптация, может участвовать в фор- мировании устойчивости к действию антибактериального препарата. Однако антибиотикорезистент-ность высокого уровня, как правило, обеспечивается действием высокоэффективных специфических механизмов устойчивости, таких как модификация мишени действия и ферментативная инактивация антибиотика.
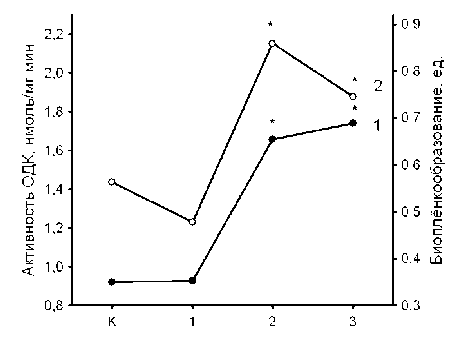
Рис. 1 . Зависимость интенсивности био-плёнкообразования и активности орнитиндекарбоксилазы E. coli от степени устойчивости к левофлоксацину:
К – контрольная группа; 1, 2, 3 – группы чувствительных, переходных и устойчивых к ЛФЦ штаммов, соответственно
В связи с этим представляло интерес изучение зависимости между интенсивностью биоплёнкооб-разования и уровнем устойчивости природных штаммов в условиях сублетального воздействия антибиотиков. С этой целью культуры штаммов E. сoli , относящиеся к разным группам по степени устойчивости к фторхинолонам, подвергали действию сублетальных концентраций левофлоксацина, оказывающих приблизительно 50%-ный ингибирующий эффект на уровень плотности культуры к 4-му часу культивирования по сравнению с контролем. Значения сублетальных концентраций (отдельно для каждого штамма) предварительно подбирали экспериментально. Добавка в среду антибиотика оказывала существенный стимулирующий эффект на интенсивность биоплёнкообразования как среди штаммов с низкой устойчивостью, так и у штаммов, имеющих высокий уровень устойчивости к левофлоксацину (рис. 2).
Известно, что ключевым ферментом синтеза полиаминов в клетках E. coli является орнитиндекарбоксилаза (ОДК), продукт которой, путресцин, является предшественником спермидина (Tabor, Tabor, 1985). Исследования базовой (неиндуциро-ванной антибиотиками) активности ОДК показали, что природные штаммы с высокой резистентностью к действию фторхинолонов обладают повышенной активностью этого фермента (рис. 1). Данная зависимость почти полностью повторяла описанную выше для биопленкообразования (рис. 1). Эти ре- зультаты свидетельствуют о том, что повышенная активность ОДК и высокая способность к биоплён-кообразованию являются признаками, свойственными фенотипу большинства устойчивых природных штаммов, прошедших отбор в условиях действия фторхинолонов. В то же время, сходный профиль кривых, описывающих эти зависимости, указывает на возможность существования взаимосвязи содержания полиаминов в клетках и их способности формировать биопленки.
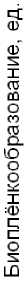
Рис. 2. Влияние сублетальной концентрации антибиотика на интенсивность биоплёнкооб-разования природных штаммов E. coli , обладающих разной степенью устойчивости к левофлоксацину.
Столбцами обозначены группы чувствительности к ЛФЦ: серые столбцы – чувствительная; заштрихованные – переходная, светлые - устойчивая.
1 – культура без добавки антибиотика; 2 – культура с добавкой сублетальной концентрации ЛФЦ.
* - статистически значимое отличие от контроля
Многолетние исследования функций полиаминов свидетельствуют о том, что эти соединения принимают участие в широком спектре адаптивных реакций бактериальной клетки (Ткаченко, Шумков, 2004; Ткаченко, Пожидаева, Шумков, 2006; Rhee, Kim, Lee, 2007), в том числе формируемых в ответ на действие антибиотиков (Ткаченко, Шумков, Ахова, 2009). Поскольку одним из механизмов, определяющих роль полиаминов в приспособлении бактерий к неблагоприятным условиям среды, является регуляция экспрессии адаптивных генов, мы предположили возможность участия этих соединений в регуляции генов биоплёнкообразования.
Результаты экспериментов с добавлением полиаминов в среду культивирования показали, что в контрольной культуре, выращенной в отсутствие антибиотика, экзогенные добавки 1 мМ путресцина и спермидина не проявляли заметного действия в разных группах чувствительности к антибиотику (рис. 3). Высокие концентрации полиаминов (10 мМ) оказывали умеренный эффект, в 1.5–2.5 раза повышая способность E. coli образовывать биоплёнки. В то же время на фоне добавки сублетальной концентрации антибиотика полиамины вызывали 2–3.5-кратную стимуляцию биоплёнкообразо-вания, причем выраженный эффект в данном случае имел место уже в ответ на добавку 1мМ путресцина и спермидина к среде культивирования (рис. 3).
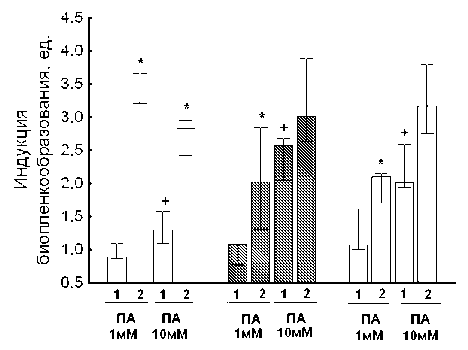
Рис. 2. Влияние полиаминов на биоплёнко-образование в отсутствие (1) и в присутствии (2) антибиотика.
Столбцами обозначены группы чувствительности к ЛФЦ: серые столбцы – чувствительная; заштрихованные – переходная, светлые - устойчивая. Сублетальные концентрации антибиотика и смесь полиаминов (ПА) путресцина и спермидина в концентрации 1 мМ и 10 мМ вносили в среду культивирования изначально.
Уровень индукции – отношение биоплёнкообразо-вания в культуре с добавкой полиаминов к контролю без полиаминов. В экспериментах с добавкой антибиотика в качестве контроля использовали культуру с добавкой левофлоксацина в отсутствие полиаминов.
* - статистически значимое отличие от контроля;
+ - статистически значимое отличие от культуры без добавки полиаминов, принятой за единицу
Исследованиями, проведенными на Y. pestis, показано, что полиамины принимают участие в регуляции синтеза белков, участвующих в первой стадии биоплёнкообразования – взаимодействии с твёрдой поверхностью (Patel et al., 2006; Wortham et al., 2010). Исходя из этого, можно предположить, что наблюдаемый нами положительный эффект путресцина и спермидина на биоплёнкообразование природных штаммов E. coli может быть обусловлен их способностью воздействовать на адгезию бактериальной клетки. Наряду с этим эффект полиаминов на биоплёнкообразование может быть опосредован через регуляцию полиаминами уровня экспрессии гена rpoS (Ткаченко, Шумков, Ахова, 2009). Под контролем продукта этого гена, σS-субъединицы РНК-полимеразы, находится по меньшей мере 14 из 17 генов, экспрессия которых возрастает в клетках E. coli во время биоплёнкооб-разования (Landin, 2009). На основании этого ка- жется обоснованным предположение, что полиамины, оказывая стимулирующее влияние на экспрессию гена rpoS (Ткаченко, Шумков, 2004; Ткаченко, Шумков, Ахова, 2009), могут оказать положительный эффект на интенсивность образования биопленок. Исходя из этого, складывается впечатление о том, что полиамины способны потенциировать индуцирующий эффект антибиотиков на биопленко-образование (рис. 3), посредством усиления экспрессии генов, участвующих в этом процессе.
Роль полиаминов в регуляции биоплёнкообразования в зависимости от присутствия левофлоксацина в среде
|
Добавка полиаминов |
Чувствительные |
Переходные |
Устойчивые |
|
1 мМ |
3.62 |
1.88 |
1.97 |
|
10 мМ |
2.18 |
1.17 |
1.58 |
Приведены значения отношения уровня индукции биоплёнкообразования полиаминами в присутствии левофлоксацина к индукции в отсутствие антибиотика.
Положительный эффект полиаминов на уровень индукции биоплёнкообразования был более выраженным в присутствии левофлоксацина у штаммов с низким уровнем устойчивости и существенно ослабевал в группе с высоким значением анртибио-тикоустойчивости (таблица). Эти различия могут быть обусловлены более существенной значимостью неспецифических механизмов адаптации, к которым относится синтез полиаминов, на начальных этапах отбора антибиотикоустойчивых вариантов. Данный признак может, по-видимому, сохраняться также в процессе селекции высокоустойчивых штаммов. С другой стороны, разница в эффекте экзогенных полиаминов на биоплёнкообразование может быть следствием повышенного содержания эндогенных путресцина и спермидина как результата высокой активности ферментов полиаминсинте-зирующей системы у высокоустойчивых штаммов, по сравнению с низкоустойчивыми (см. рис. 1).
Заключение
Результаты проведенных исследований свидетельствуют о том, что развитие антибиотикорези-стентности природных штаммов E. coli к фторхинолоновым антибиотикам сопровождается отбором признаков высокой активности системы синтеза полиаминов, связанной с повышенной способностью к биоплёнкообразованию. Фторхинолон левофлоксацин стимулирует биоплёнкообразование у штаммов как с низким, так и с высоким уровнем устойчивости к антибиотику. Полиамины оказывают положительное влияние на способность к био-плёнкообразованию у штаммов с различным уровнем устойчивости, которое существенно усиливается на фоне действия антибиотика и более выражено в группе чувствительных штаммов.
Таким образом, полиамины принимают участие в регуляции биоплёнкообразования – одного из об- щих механизмов адаптации, который участвует в развитии резистентности природных штаммов E. coli к фторхинолоновым антибиотикам как на начальных, так и на последующих этапах развития устойчивости. Полученные данные подтверждают предположение о том, что высокий уровень устойчивости E. coli к антибиотикам является результатом комбинации специфических и неспецифических механизмов резистентности.
Работа выполнена по программе Президиума РАН «Молекулярная и клеточная биология» (проект № 0120096390) и поддержана грантом РФФИ № 11-04-96001-р_урал_а.
Список литературы Полиамины как регуляторы биоплёнкообразования природных изолятов Escherichia coli с разной степенью устойчивости к фторхинолонам
- Ахова А.В., Ткаченко А.Г. Лизиндекарбоксилазная активность как фактор резистентности Escherichia coli к фторхинолонам//Микробиология. 2009. Т. 7, № 5. С. 636-640
- Кузнецова М.В. и др. Видовое разнообразие и антибиотикочувствительность грамотрицательных бактерий, изолированных в птицеводческом хозяйстве//Вестн. НГУ. 2010. Т. 8, вып. 3. С. 70-77.
- Методические указания МУК 4.2.1890-04. Определение чувствительности микроорганизмов к бактериальным препаратам//Клин. микробиол. антимикроб. химиотер. 2004. Т. 6. С. 306-359.
- Ткаченко А.Г., Нестерова Л.Ю. Полиамины как модуляторы экспрессии генов окислительного стресса у Escherichia coli//Биохимия. 2003. Т. 68. С. 1040-1048
- Ткаченко А.Г., Шумков М.С. Роль путресцина в регуляции уровня сигма S субъединицы РНК-полимеразы в клетках Escherichia coli при переходе к стационарной фазе//Биохимия. 2004. Т. 69, вып. 8. С. 1079-1087.
- Ткаченко А.Г., Пожидаева О.Н., Шумков М.С. Роль полиаминов в формировании множественной антибиотикоустойчивости Escherichia coli в условиях стрессорных воздействий//Биохимия. 2006. Т. 71, вып. 9. С. 1287-1296.
- Ткаченко А.Г., Шумков М.С., Ахова А.В. Адаптивные функции полиаминов Escherichia coli при сублетальных воздействиях антибиотиков//Микробиология. 2009. Т. 78, вып. 1. C. 32-41.
- Beloin C., Roux A., Ghigo J.M. Escherichia coli biofilms//Current topics in microbiology and immunology. 2008. Vol. 322. P. 249-289.
- Davey M.E., O'Toole G.A. Microbial Biofilms from Ecology to Molecular Genetics//Microbiol. Mol. Biol. Rev. 2000. Vol. 64, № 4. P. 847-867.
- Ghannoum M.А., O'Toole G.A. Microbial biofilms. Washington DC.: ASM Press, 2004. P. 426.
- Igarashi K., Kashiwagi K. Polyamines: mysterious modulators of cellular functions//Biochem. Biophis. Res. Commun. 2000. Vol. 271. P. 559-564.
- Landini P. Cross-talk mechanisms in biofilm formation, and responses to environmental and physiological stressin Escherichia coli//Research in Microbiology. 2009. Vol. 160. P. 259-266.
- Naves P. et al. Correlation between virulence factors and in vitro biofilm formation by Escherichia coli strains//Microbiоl. Pathogenesis. 2008. Vol. 45. P. 86-91.
- Patel C.N. et al. Polyamines are essential for the formation of plague biofilm//J. of bacterial.. 2006. Vol. 188, № 7. P. 2355-2363.
- Pratt L.A., Kolter R. Genetic analyses of bacterial biofilm formation//Curr. Opin. Microbiol. 1999. Vol. 2. P. 598-603.
- Rhee H.J., Kim E.J., Lee J.K. Physiological polyamines: simple primordial stress molecules//J. Cell Mol. Med. 2007. Vol. 11, № 4. P. 685-703.
- Tabor C.W., Tabor H. Polyamines in microorganisms//Microbiol. Rev. 1985. Vol. 49. P. 81-99.
- Tkachenko А., Nesterova L., Pshenichnov M. The role of the natural polyamine putrescine in defense against oxidative stress in Escherichia coli//Arch. Microbiol. 2001. Vol. 176. P. 155-157.
- Worthman B.W. et al. Polyamines are required for the expression of key Hms proteins important for Yersinia pestis biofilms formation//Environmental Microbiology. 2010. Vol. 12, № 7. P. 2034-2047.
