Полиморфизм нуклеолярных районов хромосом у пихты сибирской в различных экологических условиях

Автор: Калашник Надежда Александровна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 5-4 т.20, 2018 года.
Бесплатный доступ
Проведены исследования полиморфизма нуклеолярных районов хромосом у пихты сибирской, произрастающей в различных экологических условиях (равнины, поймы и долины рек, возвышенности и предгорья, промышленное загрязнение в сравнении с контролем). Полученные результаты свидетельствуют о повышении функциональной активности нуклеолярных районов хромосом у пихты сибирской как в природных экстремальных условиях так и в условиях техногенного загрязнения.
Нуклеолярные районы хромосом, пихта сибирская, различные экологические условия
Короткий адрес: https://sciup.org/148314100
IDR: 148314100 | УДК: 502.3
Polymorphism nucleolar chromosome regions in Abies sibirika under different ecological conditions
Polymorphism of nucleolar regions of chromosomes have been studied in Abies sibirika under different ecological conditions (plains, river valleys and floodplains, hills and foothills, under industrial pollution as compared to a control one). The results provide evidence for an enhancing of extreme environmental factors and technogenic pollution on functional activity of nucleolar regions chromosomes in Abies sibirika.
Текст научной статьи Полиморфизм нуклеолярных районов хромосом у пихты сибирской в различных экологических условиях
Изучение структуры кариотипа пихты сибирской проводилось на территории Казахстана [1-3], Томской области [4], Красноярского края [5-7]. Авторы, давая стандартную кариологиче-скую характеристику (число, морфометрические параметры, морфологические типы хромосом), особое внимание уделяют полиморфизму нуклеолярных районов хромосом, которые цитологически выявляются в виде вторичных перетяжек хромосом. Исследователи отмечают, что для пихты сибирской характерен высокий полиморфизм по числу, частоте встречаемости и локализации вторичных перетяжек в хромосомных наборах. Мнения авторов о причинах данного явления неоднозначны, поэтому дальнейшие исследования в этом направлении являются актуальными.
В настоящей работе представлены результаты изучения нуклеолярных районов хромосом у пихты сибирской, произрастающей на территории Южного Урала в различных экологических условиях.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
районов исследования, а также комплексная характеристика интенсивности загрязненности территорий по состоянию атмосферного воздуха, водоемов и почвенного покрова [9, 10].
Описание пробных площадей.
В районе г. Златоуста одна ПП находится на расстоянии 1.5–2 км от промышленной зоны города, вторая ПП расположена на территории свалки промышленных отходов, обе ПП подвержены многолетнему сильному техногенному загрязнению. В районе промышленной зоны г. Златоуста находятся металлургический комбинат, завод металлоконструкций, абразивный завод и ряд других производств. В районе Национального парка «Таганай» ПП находится в предгорье горы Таганай, на незначительном удалении от г. Златоуст.
В районе г. Аша ПП находится в черте города, вблизи от железнодорожного полотна, подвержена также поливалентному техногенному загрязнению со стороны промышленных предприятий города – металлургического и химического заводов. В районе г. Сим ПП находится на окраине города, вблизи (200–300 м) от автотрассы. Помимо крупной автомагистрали, источником загрязнения в городе являются выбросы ОАО «Агрегат». Загрязнение этих ПП можно охарактеризовать как умеренное.
В районе пос. Веселовка ПП находится на расстоянии 20 км южнее г. Златоуста, в долине реки Ай. В районе пос. Точильный ПП находится на расстоянии 30 км северо-восточнее г. Аша в предгорье Воробьиных гор. ПП в районе пос. Караидель и пос. Павловка находятся вблизи Павловского водохранилища на достаточно большом расстоянии от промышленных предприятий. Эти ПП определены нами как оптимальные, и использованы в качестве контрольных.
В целом, для насаждений из условий техногенного загрязнения, когда выбросы загрязнителей значительно превышают ПДК, характерно наличие большого числа суховершинных деревьев, усыхание боковых побегов, пожелтение, а чаще покраснение хвои. В этих условиях состояния насаждений охарактеризованы как «сильно ослабленные» и «ослабленные». В оптимальных и контрольных условиях состояния насаждений определены как «здоровые».
В качестве материала для проведения исследований использовали меристематическую ткань проростков семян. Исследования проводили на давленных препаратах, используя методику классического кариологического анализа, модифицированную применительно к хвойным породам [11]. Вторичные перетяжки изучали на увеличенных микрофотографиях метафаз-ных пластинок при общем увеличении x2500. По каждой ПП исследовали не менее 50 мета-фазных пластинок. Определяли среднее число вторичных перетяжек на кариотип, частоту их встречаемости (перетяжки с частотой встречаемости более 50% считали постоянными, менее 50% – непостоянными) и место локализации на хромосомном плече (sc), определяемое как отношение расстояния от вторичной перетяжки до центромеры к общей длине данного плеча, выраженное в процентах. Статистическую обработку результатов проводили общепринятыми методами [12].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
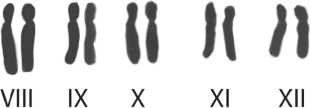
Результаты проведенных исследований показали, что среднее число вторичных перетяжек на кариотип на исследуемы пробных площадях находится в пределах от 3,12±0,15 до 7,34±0,27, их общее число варьирует – от 4 до 9, постоянных перетяжек 3–5, непостоянных перетяжек 1–4 (табл. 1). Постоянные вторичные перетяжки локализованы, как правило, на хромосомах группы I- VII (предположительно на II, III, IV и VI парах), в группе хромосом VIII-XII они встречаются реже (предположительно на IX или X парах), чаще вторичные перетяжки расположены на более коротком плече хромосомы. Непостоянные вторичные перетяжки в группе хромосом I-VII локализованы, как правило, на тех же хромосомах, что и постоянные, в группе хромосом VIII-XII они встречаются на хромосомах X, XI и XII). Некоторые вторичные перетяжки можно наблюдать на представленных микрофотографиях метафазных пластинок и систематизированных хромосомных наборах пихты сибирской исследованных ПП (рис.1, рис. 2). Место локализации как постоянных так и непостоянных вторичных перетяжек очень сильно варьирует как внутри популяций, так и между популяциями (табл.2).
Наибольшее число вторичных перетяжек наблюдается на ПП, находящихся в условиях сильного промышленного загрязнения, средние значения этого показателя наблюдается на ПП, находящихся в условиях предгорий и умеренного промышленного загрязнения, наименьшие значения - в условиях равнин, пойм и долин рек, а также условиях контроля.
В целом, у пихты сибирской исследованных ПП наблюдается тенденция увеличения числа нуклеолярных районов хромосом в экстремальных условиях. Следует отметить, что именно нуклеолярные районы хромосом (или ядрышковые организаторы хромосом) участвуют в жизненно важной функции организма – механизме белкового синтеза, а в основе интенсификации любых биосинтетических про-
Таблица 1. Число вторичных перетяжек в кариотипах пихты сибирской исследуемых пробных площадей
|
Название пробной площади |
Число вторичных перетяжек |
|||
|
постоянных |
непостоянных |
общее |
среднее |
|
|
г. Златоуст (7-ой участок) |
4 |
4 |
8 |
6,05±0,25 |
|
г. Златоуст (свалка) ** |
5 |
4 |
9 |
7,34±0,27 |
|
Национальный парк "Таганай" *** |
4 |
1 |
5 |
5,42±0,20 |
|
пос. Веселовка |
3 |
1 |
4 |
3,15±0,15 |
|
г. Аша * |
4 |
3 |
7 |
6,24±0,26 |
|
г. Сим * |
3 |
3 |
6 |
5,64±0,21 |
|
пос. Точильный |
4 |
1 |
5 |
5,21±0,19 |
|
пос. Караидель |
3 |
1 |
4 |
3,21±0,16 |
|
пос. Павловка |
3 |
1 |
4 |
3,12±0,15 |
Примечание: * – умеренное загрязнение; ** – сильное загрязнение; *** – предгорья; без обозначений –равнины, поймы и долины рек, условия контроля
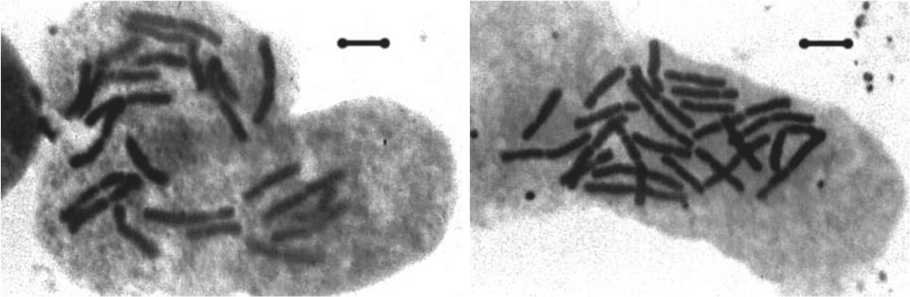
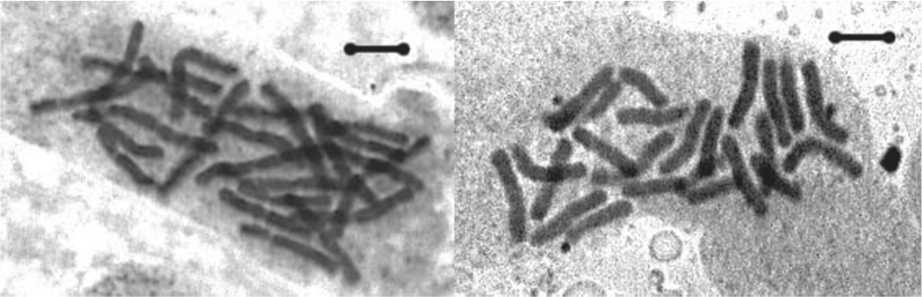
Рис. 1. Микрофотографии метафазных пластинок пихты сибирской, полученные методом классической окраски. Масштабная линия соответствует 10 мкм.
II КIIX к КII« к Х Л «(IИ
1 11 111 IV V Vl VI1 I II III IV V VI VII
U И л a»
VIII IX X XI XII
a 6
Рис. 2. Хромосомные наборы пихты сибирской исследованных ПП:
а - г. Златоуст (свалка); б - пос. Веселовка. Масштабная линия соответствует 10 мкм цессов лежит изменение активности генома, в том числе, той его части, которая ответственна за синтез рРНК [13-16]. Вероятно, пихта сибирская, произрастающая в более экстремальных условиях претерпевает значительное давление стрессовых факторов, что приводит к активизации латентных нуклеолярных локусов, то есть подключаются компенсаторные механизмы для обеспечения полноценного функционирования генома.
Полученные данные в целом согласуются с результатами исследований у пихты сибир- ской ядрышкообразующих районов хромосом, проведенных ранее другими авторами. Анализ этих результатов показывает наличие высокого полиморфизма по числу, частоте встречаемости и локализации вторичных перетяжек в кариотипах исследуемого вида. Так, в хромосомных наборах пихты сибирской, произрастающей в в различных частях ареала авторами обнаружено разное число вторичных перетяжек. Например, у пихты сибирской, произрастающей на территории Казахстана обнаружено три постоянные вторичные пе-
Таблица 2. Локализация вторичных перетяжек в кариотипах пихты сибирской исследуемых пробных площадей
ВЫВОДЫ
-
1. Результаты исследования девяти пробных площадей пихты сибирской показали, что в условиях предгорий и промышленного загрязнения у данного вида наблюдается увеличение числа функционирующих нуклеолярных районов хромосом (увеличивается общее и среднее число вторичных перетяжек на кариотип, а также число перетяжек с постоянной и непостоянной локализацией).
-
2. У пихты сибирской, произрастающей в экстремальных условиях, вероятно, происходит активизация латентных нуклеолярных локусов, то есть подключаются компенсаторные механизмы, обеспечивающие организмам процессы адаптивных изменений.
Список литературы Полиморфизм нуклеолярных районов хромосом у пихты сибирской в различных экологических условиях
- Бударагин В.А. Кариотип пихты сибирской Казахстанского Алтая // Цитология. Т.14., №1. 1972. С. 130-133.
- Бударагин В.А. Кариотип пихты сибирской Джунгарского Алатау // Леса и древесные породы Северного Казахстана. Л.: Наука, 1974. С. 81-84.
- Бударагин В.А. Кариотипы основных хвойных видов Казахстана // Тр.Каз.НИИЛХа. Алма-Ата: Кайнар, 1980. Т.2: Защитное лесоразведение и вопросы селекции в Северном Казахстане. С. 116-122.
- Седельникова Т.С. Кариологическое изучение болотной и суходольной популяций пихты сибирской (Abies sibirika Ledeb.) / Т.С.Седельникова Т.С., А.В. Пименов// Известия РАН. Серия биологическая. 2005. № 1. С. 23-29.
- Квитко О.В. Цитогенетическая и кариологическая характеристика пихты сибирской (Abies sibirika Ledeb.): автореферат дис. кандидат биологических наук: 03.00.05 / Квитко Ольга Викторовна. - Институт леса им. В.Н. Сукачева СО РАН. Красноярск, 2009. - 19 с.
