Полиплоидия в аквакультуре: преимущества и перспективы (обзор)
")
Автор: Голотин В.А., Филатова Т.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Обзоры, проблемы
Статья в выпуске: 2 т.61, 2026 года.
Бесплатный доступ
Рыба — одним из важнейших источников питательных веществ. Объемы вылова, производства и потребления рыбы неуклонно растут. Еще в 1960-х годах на душу населения планеты приходилось всего 9 кг рыбы в год. В настоящее время на каждого человека приходится около 16 кг (ФАО, 2022). В результате сокращения масштабов дикого рыболовства аквакультура стала одной из самых быстроразвивающихся отраслей сельского хозяйства. Чтобы повысить устойчивость аквакультуры, специалисты проводят селекционное разведение рыб. К сожалению, на устойчивость многих секторов отрасли негативно влияют депрессия инбридинга, вспышки болезней, недостаточное производство и низкое качество мяса. Для решения этих проблем существует настоятельная необходимость в разработке высококачественных пород рыб, обладающих высокими темпами роста, устойчивостью к болезням и/или более высокой питательной ценностью. Традиционные методы, такие как внутривидовое скрещивание и межвидовая гибридизация, успешно используются в течение нескольких десятилетий. Однако скрещивание требует гибридизации нескольких поколений, чтобы привнести желаемый признак. Кроме того, результаты часто непредсказуемы, поскольку основные механизмы, контролирующие желательные черты, неизвестны. В связи с этим существует необходимость в разработке более эффективных, точных и предсказуемых методов получения большого количества высококачественной рыбы. Достижения в области методологии разведения, основанной на классических манипуляциях с геномом и недавно разработанных методах редактирования генома, играют важную роль в будущем генетического разведения рыб. Среди известных методов, таких как племенная работа, выведение межвидовых гибридов и других, наиболее быстрыми, дешевыми и безопасными методами манипуляции с геномом являются методы полиплоидии (R.S. Rasmussen с соавт., 2007). Такие организмы не относятся к генно-модифицированным (ГМО), но при этом обладают лучшими потребительскими характеристиками по сравнению с природными видами, имеющими диплоидный набор хромосом. Поскольку они условно стерильны, то не проходят половое созревание, что, в свою очередь, позволяет сохранить потребительские свойства мяса. К тому же это обеспечивает безопасность для природных популяций от использования такой технологии в аквакультуре (L. Zhou с соавт., 2017). В настоящей статье представлен обзор мирового состояния аквакультуры, описаны известные способы получения рыб-полиплоидов, а также их характеристики и способы диагностики плоидности. В аквакультуре используют как природные полиплоиды и, в частности, триплоиды (осетровые, карповые, лососевые) (R.A. Leggatt с соавт., 2003), так и искусственно индуцированные. Основной способ получения триплоидов — блокирование экструзии второго полярного тельца после оплодотворения с помощью теплового шока или гидростатического давления (H. Komen с соавт., 2007; I.I. Solar с соавт., 1984). Тетраплоиды получают подавлением первого митотического деления, однако их хозяйственная ценность невысока; их применяют для скрещивания с диплоидами с целью получения триплоидов (Б.И. Гомельский с соавт., 1988; D. Chourrout с соавт., 1986; D. Chourrout с соавт., 1987). Перспективны также межвидовые триплоидные гибриды, например гибриды радужной форели и лосося Oncorhynchus masourhodurus(лосось Бива), отличающиеся быстрым ростом и очень хорошими органолептическими качествами (K. Hattori с соавт., 1998). Методы гиногенеза и андрогенеза позволяют получать однополые и стерильные популяции, в том числе супер-самцов желтого сома (H. Komen с соавт., 2007; D. Wang с соавт., 2009). Диагностика плоидности включает цитоморфологию крови (размеры ядер эритроцитов) (В. Tóth с соавт., 2005), подсчет метафазных хромосом (G.H. Thorgaard с соавт., 1981), проточную цитофлуорометрию (В.А. Голотин с соавт, 2023) и микросателлитный анализ (C. Howard с соавт., 2023). Ранняя диагностика на стадии бластулы особенно важна для холодноводных видов (V. Golotin с соавт., 2023). Приведены данные о более высокой продуктивности триплоидов по сравнению с диплоидами (K. Poontawee с соавт., 2007; T. Kobayashi, 1992), а также об успешном использовании триплоидной радужной форели в спортивном рыболовстве (J.R. Kozfkay с соавт., 2006).
Полиплоиды, триплоидия, диагностика плоидности, продовольственная безопасность
Короткий адрес: https://sciup.org/142247685
IDR: 142247685 | УДК: 639.3:575.113 | DOI: 10.15389/agrobiology.2026.2.226rus
Polyploidy in aquaculture: advantages and prospects (review)
Fish is one of the most important sources of nutrients. Fish harvesting, production, and consumption are steadily increasing. As recently as the 1960s, global per capita fish consumption stood at just 9 kg per year. Today, it is approximately 16 kg per person (FAO, 2022). Due to the decline in wild fisheries, aquaculture has become one of the fastest-growing sectors of agriculture. To improve the sustainability of aquaculture, experts are conducting selective breeding of fish. Unfortunately, the sustainability of many sectors of the industry is negatively affected by inbreeding depression, disease outbreaks, low productivity, and inferior meat quality. To address these issues, there is an urgent need to develop high-quality fish breeds with rapid growth rates, disease resistance, and/or improved nutritional value. Traditional methods, such as intraspecific crossbreeding and interspecific hybridization, have been used successfully for several decades. However, these methods require several generations of hybridization to introduce the desired trait. Furthermore, the results are often unpredictable, as the underlying mechanisms controlling the desired traits are unknown. Consequently, there is a need to develop more efficient, precise, and predictable methods for producing large quantities of high-quality fish. Advances in breeding methodology (based on classical genome manipulation and recently developed genome editing techniques) play a crucial role in the future of fish genetic breeding. Among well-known methods (breeding, the production of interspecific hybrids, and others), the fastest, cheapest, and safest genome manipulation methods are polyploidy induction (R.S. Rasmussen et al., 2007). Polyploid organisms are not genetically modified organisms (GMOs), but possess superior consumer qualities compared to natural species with a diploid chromosome set. Since they are effectively sterile, they do not undergo sexual maturation, which preserves meat quality. Moreover, this protects wild populations from the potential impact of using such technology in aquaculture (L. Zhou et al., 2017). This article provides an overview of the global state of aquaculture, describes known methods for producing polyploid fish, their characteristics, and methods for ploidy determination. In aquaculture, both natural polyploids, specifically triploids (sturgeons, carp, and salmonids) (R.A. Leggatt et al., 2003), and artificially induced polyploids are used. The primary method for producing triploids involves blocking the extrusion of the second polar body after fertilization using thermal shock or hydrostatic pressure (H. Komen et al., 2007; I.I. Solar et al., 1984). Tetraploids are produced by inhibiting the first mitotic division; however, they have a little practical value and can be used for crossing with diploids to produce triploids (B.I. Gomelsky et al., 1988; D. Chourrout et al., 1986; D. Chourrout et al., 1987). Interspecific triploid hybrids also show promise, for example, hybrids of rainbow trout and Oncorhynchus masou rhodurus (Biwa salmon), which are characterized by rapid growth and excellent organoleptic qualities (K. Hattori et al., 1998). Methods of gynogenesis and androgenesis allow for the production of unisexual and sterile populations, including “super-males” of the yellow catfish (H. Komen et al., 2007; D. Wang et al., 2009). Ploidy diagnosis includes blood cytomorphology (red blood cell nucleus size) (V. Tóth et al., 2005), metaphase chromosome counting (G.H. Thorgaard et al., 1981), flow cytometry (V,A, Golotin et al., 2023), and microsatellite analysis (C. Howard et al., 2023). Early diagnosis at the blastula stage is particularly important for cold-water species (V. Golotin et al., 2023). Data are presented on the higher productivity of triploids compared to diploids (K. Poontawee et al., 2007; T. Kobayashi, 1992), as well as on the successful use of triploid rainbow trout in sport fishing (J.R. Kozfkay et al., 2006).
Текст обзорной статьи Полиплоидия в аквакультуре: преимущества и перспективы (обзор)
Рост численности населения в мире, деградация экосистем, клима-
* Исследование выполнено при частичной поддержке Российского научного фонда (грант № 23-26-00257).
тические кризисы, утрата биоразнообразия угрожают окружающей среде и мировой экономике (1). При этом последствия пандемии COVID-19, политических кризисов и военных конфликтов усугубляет проблему (2). Продолжающийся рост населения ведет к увеличению объемов потребления. Полагают, что глобальный спрос на продовольствие будет повышаться еще как минимум 40 лет (3).
В настоящее время, по данным ООН, в мире насчитывается более 800 млн голодающих. Кроме того, около 3 млрд человек недоедают или не могут позволить себе сбалансированное питание, из-за чего развиваются различные патологии. Например, при голодании в раннем возрасте у людей развивается метаболический синдром и другие нарушения (4). Из-за большого числа голодающих необходимы преобразования агропромышленных систем для обеспечения продовольственной безопасности, когда при сохранении природных ресурсов население обеспечивается доступным и высококачественным здоровым питанием.
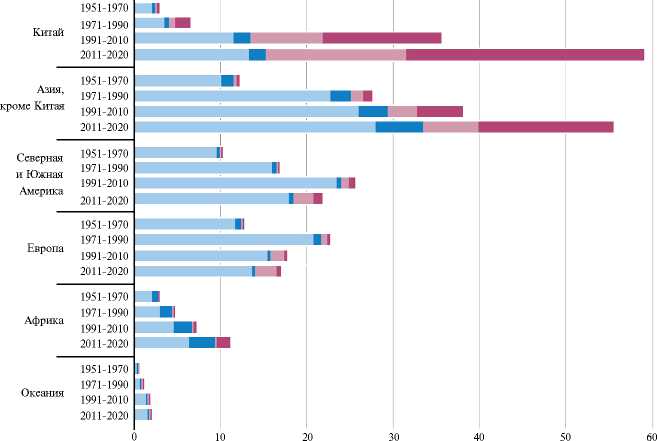
Важная роль отводится продукции из водных биоресурсов, служащих источником не только белка, но и рекордного количества уникальных полиненасыщенных жирных кислот и биодоступных микроэлементов. В 2020 году в сегменте рыболовства и аквакультуры был произведен наибольший объем продукции: 214 млн т общей суммой около 424 млрд долларов, что в среднем на 60 % превышало показатель в 1990-е годы, значительно опередив рост населения. Это обусловлено преимущественно резким скачком производства в аквакультуре (рис. 1). Стоит обратить внимание, что к 2020 году увеличилось потребление водных биоресурсов до 20 кг на душу населения, удвоив таковой показатель полвека назад (5).
Рис. 1. Объемы продукции мирового промышленного рыболовства и аквакультуры и прогноз до 2030 года (11) .
Обращаясь к истории развития аквакультуры, можно отметить, что самая ранняя ее форма в эпоху неолита заключалась в отлове диких водных животных в прудах, лагунах, мелководных озерах. И если в Китае методы разведения карпа совершенствовались уже в 2600 г. до н.э. (6, 7), то в Европе аналогичные методы стали развиваться только в средние века, особенно в монастырях, нуждавшихся в немясном источнике белка в течение многодневных христианских постов (8).
Цель настоящего обзора — проанализировать мировое состояние аквакультуры, обобщить имеющиеся способы получения рыб-полиплоидов, перспективных для выращивания в целях обеспечения продовольственной безопасности, а также охарактеризовать их биологические особенности и методы диагностики плоидности.
Мировое состояние аквакультуры. Между континентами и странами существуют значительные различия в производстве водных биоресурсов (9). Так, несмотря на пик пандемии коронавируса в 2020 году, больше всего продукции производилось в странах Азии — 70 % общего объема, следом шли страны Северной и Южной Америки, Европы, Африки и Океании. Лидером остается Китай, поставивший на рынки 35 % продуктов водных биоресурсов (см. рис. 1, 2) (10).
К 2030 году прогнозируется рост производства гидробионтов на 15 %. Преимущественно он будет обеспечен за счет прироста объемов аквакультуры, которые к 2030 году могут достичь более 100 млн т (5, 11).
Объем производства, млн т
Промышленное рыболовство — морские районы
| Промышленное рыболовство — | Аквакультура — | Аквакультура — внутренние водоемы морские районы внутренние водоемы
Рис. 2. Доля регионов в общемировом объеме производства продукции промышленного рыболовства и аквакультуры (5) .
Аквакультурное производство играет одну из ключевых ролей для минимизации разрыва между спросом и предложением на такую пищевую продукцию (12). Прогнозируемая модернизация этого сектора, повышение производительности и интенсификация производства окажут, в том числе, положительные эффекты на занятость населения (5). Инновационные технологические решения позволят производить больше, используя меньше ресурсов за счет оптимизации производственных процессов. Эти тенденции в аквакультуре не угрожают ростом бедности, поскольку коммерческие системы рыбоводства ограничивают рост цен на рыбу, что ведет к увеличению ее потребления бедными и беднейшими слоями населения (13).
В то же время интенсификация аквакультуры может повысить риски ухудшения состояния окружающей среды, интродукции видов, развития болезней, что характерно для любой сельскохозяйственной системы при интенсификации производства продуктов питания (14, 15), в том числе из-за использования в аквакультуре антибиотиков (16). Для дальнейшего развития аквакультуры важно, чтобы ее рассматривали в контексте более широкого многоуровневого управления, то есть должны учитываться последствия как преднамеренных, так и непреднамеренных вмешательств в аквакультуру при формировании политики на различных территориальных уровнях (17). Большие ожидания связывают с инновациями, позволяющими повысить количество производимой продукции, а именно с цифровизацией процессов и продвинутыми технологиями (17).
Устойчивая аквакультура — это высокоэффективный способ производства белков животного происхождения при минимальном потреблении зерновых культур (18), вносящий значительный вклад в мировую продовольственную безопасность и питание человека (19, 20). Традиционные методы в аквакультуре, такие как племенная работа, выведение перспективных межвидовых гибридов, отбор по фенотипу, гибридизация между линиями, использование гетерозиса, занимают сравнительно продолжительное время. Дальнейшее развитие аквакультуры должно быть направлено на минимизацию ущерба от болезней, разрастания водорослей, а также на улучшение иммунитета и товарных качеств рыбной продукции.
Перспективным направлением в этом вопросе представляется выведение полиплоидных организмов, которые отличаются от диплоидных форм устойчивостью к различным заболеваниям и более крупной навеской преимущественно из-за сдерживания полового развития. Кроме того, мясо триплоидных особей обладает лучшими потребительскими качествами, например у форели цвет мяса становится более красным, а содержание влаги в нем снижается (эти работы подробнее рассмотрим ниже).
Перспективы полиплоидов в аквакультуре. Полиплоидия может быть естественным, природным, генетическим состоянием, а может быть создана искусственно у рыб и моллюсков посредством манипуляций с эмбрионами. Полиплоидные организмы имеют дополнительные наборы хромосом. И хотя это состояние смертельно для птиц и млекопитающих, в аквакультуре полиплоидия показала многообещающие результаты (21).
Несмотря на манипуляции с геномом гидробионтов, они не относятся к генно-модифицированной продукции, споры о продовольственной безопасности которой не утихают и по сей день. Полиплоидия создает множество преимуществ и возможностей, приводящих к увеличению аллельного разнообразия, появлению новых признаков, изменению сложности генома, расширению географического распространения видов (22-25). Именно эти преимущества (увеличение аллельного разнообразия, появление новых признаков, изменение сложности генома, расширение географического распространения видов) придают полиплоидным организмам экономически важные хозяйственные признаки, такие как быстрый рост, обширная приспособляемость и устойчивость к болезням, благодаря чему они востребованы для аквакультуры (26).
Мировая аквакультура широко использует как полиплоиды естественного происхождения (табл. 1), так и искусственно индуцированные (табл. 2) (27-30).
Искусственное дублирование хромосомы считается классическим подходом к манипулированию геномом. В природе многочисленные независимые полногеномные дупликации, ведущие к образованию полиплоидов из предковых диплоидов, сыграли важную роль в эволюции, предоставив избыточные копии генов, которые впоследствии могут накапливать аллельную изменчивость и функциональную диверсификацию, тем самым открывая возможности для полиплоидов занимать различные ниши и, в конечном счете, развиваться в новые виды (31).
-
1. Природные полиплоиды водных животных, используемые в аквакультуре
-
2. Искусственные полиплоиды, используемые в аквакультуре
Семейство
Вид или разновидность
Плоид-ность
Методы
Ссылка
Карповые
Белый амур ( Ctenopharyngodon idella )
3 n
Гидростатическое давление (60)
( Cyprinidae )
Карась Сянъюнь
6 n
Межвидовая гибридизация
(41, 61,
Карась Сянъюнь Crucian carp )
«Changfeng» Серебряный карась ( Synthetized allopolyploid )
6 n
8 n
и межплоидное скрещивание
Включение чужеродного
генома в сперматозоиде
62)
Вьюновые ( Cobitidae )
Амурский вьюн ( Misgurnus anguillicaudatus )
3 n
Межплоидное скрещивание
Лососевые ( Salmonidae )
Радужная форель ( Oncorhynchus mykiss )
3 n
Тепловой шок, гидростатическое давление или межплоидное скрещивание
(28, 29, 63-65)
Кумжа ( Salmo trutta )
Американская палия ( Salvelinus fontinalis ) Атлантический лосось ( Salmo salar ) Чавыча ( Oncorhynchus tshawytscha ) Арктический голец ( Salvelinus alpinus ) Форель Бива ( Oncorhynchus masourhodurus ) Сима ( Oncorhynchus masou )
Кижуч ( Oncorhynchus kisutch )
3 n
3 n
3 n
3 n
3 n
3 n
3 n
3 n
Гидростатическое давление
(66)
(67)
(68)
(69)
(70)
(27)
(27)
(71)
Аллотриплоидный гибрид радужная форель ( Oncorhynchus mykiss ) ½ лосось амаго ( Oncorhynchus masou macrostomus )
3 n
Межвидовая гибридизация и тепловой шок
(72)
Кета ( Oncorhynchus keta ) ½ гибрид японского гольца ( Salvelinus leucomaenis )
3 n
Межвидовая гибридизация и тепловой шок
(27)
Сомовые ( Siluridae )
Тайский ходячий сом ( Clarias batrachus ) ½ гибрид африканского сома ( Clarias gariepinus )
3 n
Межвидовая гибридизация и тепловой шок
(73)
Плекоглоссовые Айю ( Plecoglossus. altivelis ) ( Plecoglossidae )
3 n
Тепловой шок или гидростатическое давление
(27-29)
Паралихтиевые ( Paralichthyidae )
Хираме ( Paralichthys olivaceus )
3 n
Обработка давлением
Моллюски, Устрицы ( Ostreidae )
Тихоокеанская устрица ( Crassostrea gigas )
3 n
Тепловой шок, гидростатическое давление или межплоидное скрещивание
(74, 75)
Пектинидовые ( Pectinidae )
Гребешок Катарина ( Argopecten ventricosus )
3 n
Цитохалазин В
(76)
|
Семейство, класс |
Вид |
Число хромосом |
Плоидность |
Ссылка |
|
Осетровые |
Веслонос ( Polyodon spathula ) |
120 |
4 n |
(35, 43) |
|
( Acipenseridae ) |
Русский осетр ( Acipenser gueldenstaedtii ) |
236±2 |
8 n |
(32) |
|
Китайский осетр ( A. sinensis ) |
240 |
8 n |
||
|
Стерлядь ( A. ruthenus ) |
116±4 |
4 n |
||
|
Амурский осетр ( A. ruthenus ) |
238±8 |
8 n |
||
|
Сибирский осетр ( A. baerii ) |
∼ 245 |
8 n |
||
|
Дунайский осетр ( A. gueldenstaedtii ) |
240-264 |
8 n |
||
|
Белый осетр ( A. transmontanus ) |
240-264 |
8 n |
||
|
Адриатический осетр ( A. naccarii ) |
240-264 |
8 n |
||
|
Калуга ( Huso dauricus ) |
268±4 |
8 n |
||
|
Белуга ( H. huso ) |
∼ 120 |
4 n |
||
|
Карповые |
Обыкновенный карп ( Cyprinus carpio ) |
100 |
4 n |
(44) |
|
( Cyprinidae ) |
Карась ( Carassius auratus ) |
100 |
4 n |
(45-47) |
|
Золотая рыбка ( C. auratus ) |
100 |
4 n |
||
|
Карп гибелио ( C. gibelio ) |
156 или 162 |
6 n |
||
|
Усач ( Barbus barbus ) |
100 |
4 n |
(48, 49) |
|
|
Алжирский шип ( B. callensis ) |
100 |
4 n |
||
|
Барбодес ( Spinibarbus denticulatus ) |
100 |
4 n |
||
|
( Schizothorax prenanti ) |
148 |
6 n |
(50) |
|
|
( S. davidi ) |
98 |
4 n |
||
|
Прудовый голец ( Misgurnus anguillicaudatus ) |
75, 100, 150 |
3 n , 4 n , 6 n |
(51-53) |
|
|
Чукучановые |
Большеротый буффало ( Ictiobus cyprinellus ) |
100 |
4 n |
(32) |
|
( Catostomidae ) |
||||
|
Лососевые |
Европейский сиг ( Coregonus lavaretus ) |
80 или 81 |
4 n |
(32, 54, 55) |
|
( Salmonidae ) |
Пелядь ( Coregonus peled ) |
74 |
4 n |
|
|
Арктический голец ( Salvelinus alpinus ) |
78 |
4 n |
||
|
Ручьевая форель ( S. fontinalis ) |
84 |
4 n |
||
|
Атлантический лосось ( Salmo salar ) |
54-58 |
4 n |
||
|
Коричневая форель ( S. trutta ) |
78-84 |
4 n |
||
|
Севанская форель ( S. ischchan ) |
80-82 |
4 n |
||
|
Чавыча ( Oncorhynchus tshawytscha ) |
68 |
4 n |
||
|
Кета ( O. keta ) |
74 |
4 n |
||
|
Кижуч ( O. kisutch ) |
60 |
4 n |
||
|
Радужная форель ( O. mykiss ) |
58-64 |
4 n |
||
|
Золотая форель ( O. mykiss aguabonita ) |
58 |
4 n |
||
|
Лосось Масу ( O. masou ) |
66 |
4 n |
||
|
Нерка ( O. nerka ) |
57 или 58 |
4 n |
||
|
Дунайский лосось ( Hucho hucho ) |
82 |
4 n |
||
|
Европейский хариус ( Thymallus thymallus ) |
102 |
4 n |
||
|
Корюшковые |
Речная малоротая корюшка ( Hypomesus |
|||
|
( Osmeridae ) |
olidus ) |
56 |
4 n |
|
|
Саланксовые |
Clearhead icefish ( Protosalanx hyalocranius ) |
56 |
4 n |
|
|
( Salangidae ) |
||||
|
Сомообразные |
Филиппинский сом ( Clarias batrachus ) |
100 |
4 n |
(32) |
|
( Siluriformes ) |
Мешкожаберный сом ( Heteropneustes fossilis ) |
87, 116 |
3 n , 4 n |
(56) |
|
Ракообразные |
Красный болотный рак ( Procambarus clarkii ) |
192/188 |
4 n |
(57, 58) |
|
( Crustacea ) |
Японский рак ( Cambarus japonicas ) |
196 или 200 |
4 n |
|
|
Лангуст ( Nephropsis carpenter ) |
152 |
4 n |
||
|
Японский лобстер ( N. japonicus ) |
136 или 164 |
4 n |
||
|
Моллюски |
Улитка ( Neptunea cumingi ) |
60 |
4 n |
(59) |
|
( Mollusca ) |
Задокументировано влияние тетраплоидии, происходящей от независимой полногеномной дупликации, на организацию и модификацию геномов в течение эволюционного времени для рыб (32-34). Также существуют четко установленные полиплоидные виды, такие как осетр или веслонос, предки которых, как полагают, стали тетраплоидами около 200 млн лет назад (35). Кроме того, есть группы видов, у которых плоидность пока не стабилизировалась, например комплекс Rutilus alburnoides ( Cyp-rinidae , Leuciscinae ), в котором существуют диплоидная, триплоидная и тетраплоидная формы. В дополнение к этому имеются сообщения о спонтанно возникающих полиплоидах у различных видов рыб, которые теоретически могут дать начало будущим полиплоидным видам (34).
Считается, что добавление только одного набора хромосом (три- плоидия), как правило, становится эволюционным тупиком, поскольку обладание нечетным числом гомологичных хромосом мешает мейозу и приводит к бесплодию, за некоторыми заметными исключениями (23). Среди рыб стерильность триплоидов открывает возможности для их использования в аквакультуре и управления рыболовством для предотвращения нежелательного полового созревания и размножения (36).
У некоторых видов рыб темпы роста различаются у самок, самцов и бесплодных особей. Чтобы воспользоваться этим, рыбоводы выращивают однополые или бесплодные популяции для повышения продуктивности. Для достижения контроля пола и фертильности во время разведения рыбы было использовано полиплоидное размножение как одно из наиболее эффективных (26). Этот метод также может быть использован для быстрого получения гомозиготной популяции и уменьшения продолжительности процесса генетического размножения в поколениях.
Концепция полиплоидного размножения была разработана на основе исследований гиногенеза. У рыб второе мейотическое деление завершается вскоре после овуляции или оплодотворения. Этот процесс может быть подавлен холодовым шоком, что приводит к дублированию хромосом в яйцеклетке (37, 38). Комбинация физических методов воздействия, таких как тепловой удар и гидростатическое давление, дает наилучшие результаты (39). Гиногенез у рыб индуцируется термическим шоком или обработкой гидростатическим давлением после оплодотворения яйцеклетки инактивированным сперматозоидом (40, 41). И наоборот, ан- дрогенез индуцируется термическим шоком или обработкой гидростатическим давлением инактивированной яйцеклетки, оплодотворенной нормальной спермой. Как гиногенез, так и андрогенез были использованы на различных видах рыб для получения двойных гаплоидов (39). Производство двойных гаплоидов может быть использовано для поддержания моно-сексуальной популяции. У видов с определением пола XX-XY ожидается, что потомство, полученное в результате гиногенеза, будет полностью женского пола, а потомство, полученное в результате андрогенеза, будет иметь геном XX или YY. Эти признаки меняются местами у видов с определением пола ZW-ZZ. Использование гиногенеза и андрогенеза для контроля пола подходит только для производства небольших популяций из-за относительно высокой смертности, которая, вероятно, вызвана повреждением от облучения, побочными эффектами от теплового/давящего шока и депрессией инбридинга (39). Примечательно, что методы размножения с полиплоидией можно комбинировать с другими классическими методами, такими как индуцированное изменение пола для контроля пола потомства и уменьшения продолжительности процесса размножения. Один из успешных примеров такого подхода — производство «супер-самцов» желтого сома (Pelteobagrus fulvidraco), у которого самцы растут быстрее, чем самки (42). Популяция желтого сома YY-мужского пола получена в результате гиногенеза индуцированных физиологических самок генотипа XY, а YY-линия поддерживается посредством скрещивания с индуцированными самками YY с измененным полом. Как и в случае с другими животными, стерильные рыбы обычно имеют более высокие темпы роста, чем фертильные особи.
По хромосомному составу и механизмам образования полиплоиды делятся на аутополиплоиды и аллополиплоиды (23) (см. табл. 1, 2).
Нельзя обойти вниманием триплоидных межвидовых гибридов, которых применяют в аквакультуре очень давно. Благодаря триплоидизации генома становится возможным получение не скрещивающихся в природе жизнеспособных гибридов лососевых. Так, некоторые из межвидовых три-плоидных гибридов, например гибридов самок форели Oncorhynchus mykiss и самцов лосося Oncorhynchus masou rhodurus , обладают ценными хозяйственными признаками, такими как ускоренный рост, отличные вкусовые качества мяса. У гибридов на теле отсутствуют пятна, что нравится конечным потребителям (72).
Индуцированная тетраплоидия у рыб. Получение искусственных полиплоидов казалось перспективной идеей, и более полувека назад были начаты первые эксперименты по получению тетраплоидной радужной форели (77). Организм, содержащий четыре набора хромосом, возможно создать из оплодотворенной яйцеклетки, подавив первое деление митоза. После такого воздействия образуется одна тетраплоидная клетка вместо двух диплоидных, дающая начало тетраплоидному организму (рис. 3). Для подавления митотического деления оплодотворенной яйцеклетки применяют различные воздействия, а именно гидростатический или тепловой шок. Последний способ более эффективен для устойчивых к холоду представителей лососевых. Были получены тетраплоидные радужная форель ( O. mykiss) (78, 79), атлантический лосось ( S. salar ) (80) и кумжа S. trutta (81). Однако зафиксирован низкий темп роста тетраплоидов на примере радужной форели. Также оказалась низкой фертильность самцов и общая выживаемость тетраплоидов (82).
Таким образом, эксперименты показали, что получение и поддержание тетраплоидных линий лососей в принципе возможно, но их хозяй-232
ственные показатели бесперспективны. Однако применение этим рыбам было найдено: их используют для получения триплоидов, оказавшихся в производственном отношении более перспективными, посредством скрещивания с диплоидными особями (83, 84). В то же время необходимо отметить некоторые недостатки этого метода получения триплоидов. На практике при скрещивании диплоидов с тетраплоидами радужной форели в потомстве могут появляться гаплоиды, диплоиды и мозаики, поскольку предполагаемые тетраплоиды могут быть на самом деле диплоидно-тетра-плоидными мозаиками (82).
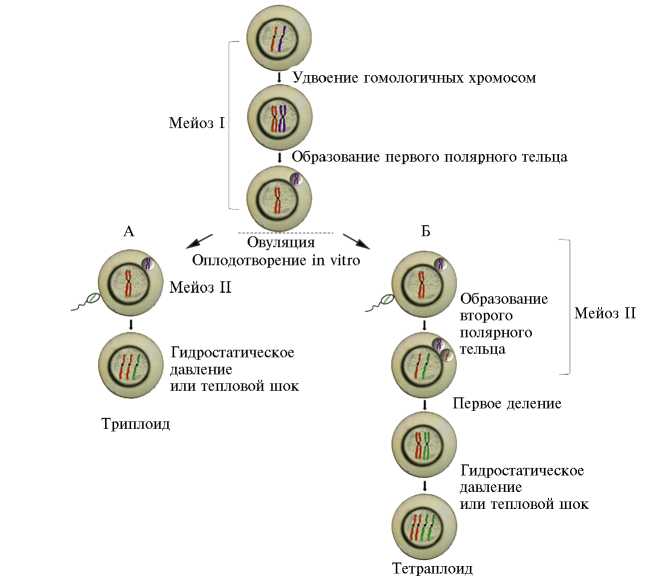
Рис. 3. Схематическое изображение триплоидных и тетраплоидных манипуляций у рыб: А — индукция искусственного триплоида посредством блокирования экструзии второго полярного тельца за счет гидростатического давления, холода или теплового шока, Б — индукция искусственного тетраплоида посредством подавления первого деления дробления за счет гидростатического давления, холода или теплового шока (26).
Индуцированная триплоидия у рыб. Первые успешные опыты по плоидным манипуляциям на хромосомах рыб, сравнение роста и полового созревания с аналогичными показателями у контрольных диплоидных рыб были проведены в 1940-1950-х годах. Как следствие, индукция триплоидии стала гораздо более частой и в настоящее время распространена на многие виды морских и пресноводных рыб (85). Получение рыб-триплоидов особенно востребовано в связи с тем, что триплоидия представляет собой сравнительно дешевую манипуляцию для получения желаемых товарных характеристик, в отличие от относительно более долгих и дорогих работ по выведению межвидовых гибридов и селекционных работ по выведению новых пород рыб.
Кроме скрещивания диплоидов и тетраплоидов, для получения триплоидов используют такие же воздействия (тепловой и холодовой шок, гидростатическое давление), но после оплодотворения, подавляя не первое дробление зиготы, а второе деление мейоза, начинающегося вследствие активации икринки вскоре после осеменения (86). У рыб, в отличие от высших позвоночных, второе деление мейоза происходит вне организма (87). Шоковое воздействие приводит к разрушению веретена деления, оставляя ядро икринки диплоидным, а после его слияния с ядром гаплоидного мужского пронуклеуса оплодотворенная икринка обретает дополнительный набор хромосом (см. рис. 3).
По расчетам ученых из Франции, продуктивность триплоидов, на практике опережает диплоидов после достижения трехлетнего возраста. Так, продуктивность самок-триплоидов разных видов лососевых на четвертом году жизни (возраст 3+) составляет более 130 % от продуктивности контроля (88). Опубликованы факты о том, что в возрасте 6,5 лет трипло-иды радужной форели ( O. mykiss ) имели массу 5,9 кг, диплоиды — 4,2 кг (89). В другом исследовании масса триплоидных самок радужной форели возрастом 59 мес составляла 6,6 кг, что примерно на 30 % больше, чем у контрольных самок (90). Помимо этого, их отличает лучшее качество мяса в сравнении с теми же диплоидами (89, 91).
Выпуски радужной форели с тройным набором хромосом в водоемы на территории США, где практикуется спортивное рыболовство, полностью себя оправдали и в ряде штатов стали ежегодной практикой (92). В Канаде в водохранилище выловили триплоидную рыбу массой 19,67 кг, оказавшуюся самой крупной радужной форелью среди выловленных в пресной воде в мире. Таким образом, триплоиды служат прекрасными объектами для спортивной рыбалки (93). Успешное использование трипло-идов в качестве объектов спортивного рыболовства объясняются условной стерильностью таких рыб и невозможностью натурализоваться в водоемах, куда их выпускают. В то же время они могут пребывать в водоемах длительно, не гибнут из-за неподходящих для нереста условий, в том числе разной солености, и продолжают постоянно расти. Солевая адаптация три-плоидов радужной форели одинаковая с таковой у диплоидов (94).
Отмечается, что попавшие в море триплоиды атлантического лосося намного реже, чем диплоиды, заходят в реки (95), вследствие чего сбежавшие из рыбоводных садков триплоиды значительно меньше оказывают влияние на пресноводные экосистемы, не занося в них патогены.
Триплоидия влияет на лекарственную устойчивость культивируемой кишечной микробиоты у молоди лосося, что, вероятно, способствуют здоровью триплоидных рыб и имеют важные последствия для использования антибактериальных препаратов в аквакультуре (96).
Иногда у триплоидов бывают пороки развития, редко встречающиеся у диплоидных рыб. Например, отмечаются аномалии строения позвоночника и нижней челюсти (97, 98). Механизмы этого явления пока неизвестны, но триплоиды в сравнении с диплоидами имеют другие пищевые потребности, что, возможно, и становится причиной деформации скелета. Триплоиды имеют, в отличие от диплоидов, другие температурные границы, поэтому важно определить физиологические эффекты температуры на триплоидов. Температура может влиять на возникновение деформаций, работу сердечно-сосудистой системы, физиологическую реакцию на стресс и иммунную компетентность на всех этапах жизни. Таким образом, устойчивость к температуре может быть основной причиной более низкой производительности триплоидов по сравнению с диплоидами как в культуре, так и в естественных условиях (99).
Основные методы получения полиплоидов рыб. Для получения триплоидов радужной форели обычно используют тепловой шок (78). К преимуществам этого метода, безусловно, можно отнести 234
возможность проведения процедуры без специального оборудования, что значительно удешевляет процесс. Однако при получении триплоидов с использованием термошока наблюдаются значительно большие отходы икры, чем в контроле. Выход личинок — не более 70-80 % от нормы. Экспериментально установлено, что у разных пород радужной форели при одинаковом режиме термошока доля триплоидов различается. При получении триплоидов у осенненерестующих пород форели термошок более эффективен в сравнении с зимненерестующими (100). Помимо этого, прослеживается разный выход триплоидов в разных семействах и даже породах одних и тех же видов рыб (99).
Используя практику работы с радужной форелью, для создания триплоида атлантического лосося также используют повышенное давление и тепловой шок; в частных экспериментах высокое давление сочетали с действием анестетиков. Однако при использовании холодового шока три-плоиды получить не удавалось (78, 101, 102).
По мере развития и совершенствования приемов получения три-плоидных лососей кроме термошока все чаще стали использовать метод гидростатического давления. По данным исследований (103), отходы икры в процессе инкубации и доля уродливых экземпляров в потомстве при применении этого метода были меньше.
Суть метода состоит в том, что оплодотворенная икра помещается в герметичную емкость, и спустя время, необходимое для формирования веретена второго деления мейоза (10-25 мин), в этой емкости создается давление, превышающее атмосферное. Параметры давления и время воздействия на икру подбираются экспериментально. В научной литературе, однако, эти параметры не всегда сообщаются, поскольку параметры получения триплоидной икры, остаются коммерческой тайной (104).
Использование гидростатического давления при промышленном создании радужной форели с тройным набором хромосом в последнее время позволяет получать стерильных рыб. Таких рыб можно безопасно выпускать в естественные водоемы без риска их гибридизации с особями из природных популяций. Таким же образом получают триплоидов кумжи (105).
В эксперименте по триплоидизации беломорской горбуши Oncorhyn-chus gorbuscha были сделаны выводы о нецелесообразности использования триплоидов горбуши в аквакультуре (106).
Показано, что экономически целесообразно получать и выращивать только триплоидных самок в связи с различиями в степени развития гонад у триплоидных самок и самцов (у самцов гонады более развиты) (107).
Диагностика плоидности. Важнейшая методическая часть при изучении природной и индуцированной полиплоидии — ее своевременная и точная диагностика. В зависимости от потребностей исследования, наличия соответствующего лабораторного оснащения и квалификации персонала, существует несколько подходов к такой диагностике у рыб (рис. 4).
Самым примитивным способом считается цитоморфология крови, результаты которой основаны на данных о количестве ДНК в ядрах эритроцитов, соотношении площадей ядер эритроцитов у особей с различным уровнем плоидности (108) и подсчете максимального количества ядрышек в ядрах эритроцитов на препаратах крови, окрашенных 50 % AgNO3 (109). Площади ядер считают в графических редакторах в ручном визуальном или автоматическом режиме с помощью различного программного обес- печения (110), а полученные значения используют для расчета объемов клеток и их ядер (111). Соотношения объемов клеток и их ядер коррелируют с плоидностью (рис. 4, А).
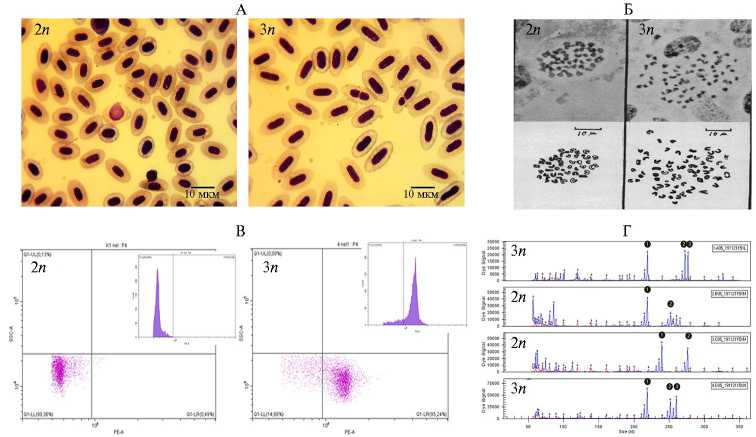
Рис. 4. Методы диагностики триплоидии у рыб. А — визуальный метод, основанный на размерах ядер клеток: слева — клетки крови сеголеток диплоидной нельмы ( Stenodus leucichthys nelma ), справа — клетки триплоидов нельмы (122). Б — митотические пластинки эпителиальных клеток Poeciliopsis lucida ; метод, основанный на подсчете числа хромосом: слева — митотическая пластика диплоида (48 хромосом), справа — триплоида (72 хромосомы) (123). В — точечные графики и гистограммы распределения содержания ДНК в выделенных эмбриональных единичных клетках нельмы (124), окрашенных иодидом пропидия, полученные методом проточной цитометрии (РЕ-канал): на оси абсцисс показано содержание ядерной ДНК клеток в фазе G1 клеточного цикла, которое выражается в произвольных единицах интенсивности флуоресценции, все оси и разделительные линии зафиксированы в одинаковых положениях (слева — диплоиды, справа — триплоиды). Г — способ определения плоид-ности фрагментным анализом ДНК (на примере Salmo salar ): три отчетливых пика (верхний и нижний графики) или два пика (два графика посередине) свидетельствуют о триплоидии и диплоидии (и/или гомозиготный триплоидии) соответственно; пики показывают интенсивность флуоресценции при определенных размерах фрагментов; ранние пики около 50-100 п.н. являются ошибками ПЦР (121).
Пожалуй, наиболее трудоемкий способ определения плоидности — подсчет числа хромосом (см. рис. 4, Б). Для такого анализа готовят хромосомные препараты из различных тканей, взятых на разных стадиях жизни рыб. Высокой митотической активностью обладают эмбрионы рыб, а также пронефрос взрослых особей (112). Морфологию хромосом и подсчет их числа осуществляют с помощью микроскопа во время митоза, особенно в середине митоза на стадии метафазы, когда они максимально спирализова-ны (113). Чтобы заблокировать деление хромосом на этой стадии, рыбам делают инъекцию или обрабатывают используемые для приготовления препаратов ткани химическими веществами (колхицин, колцемид сульфат винбластина), которые предотвращают формирование нити веретена деления (114). При отсутствии видовых особенностей число хромосом кратно числу гаплоидных хромосомных наборов.
Наиболее часто проводят анализ плоидности методом проточной цитофлуорометрии (см. рис. 4, В), который позволяет достаточно точно и оперативно (на ранних этапах эмбриогенеза или у взрослых особей) оценить плоидность рыб, например при индуцированной триплоидизации (60, 115). Материалом для проточного цитофлуориметра служат суспензии живых или фиксированных клеток, содержащих ядро и окрашенных флуоресцентными красителями, такими как DAPI или иодид пропидия, которые довольно специфично связываются с нуклеиновыми кислотами клетки (116). Проточный цитометр способен анализировать огромное количество клеток за малый промежуток времени (размеры клеток и интенсивность их флуоресценции при окрашивании). По интенсивности излучаемой флуорсценции клеток можно диагностировать триплоидию у рыб относительно известного диплоидного контроля. Интенсивность флуоресценции коррелирует с числом наборов хромосом в каждом анализируемом массиве клеток.
При применении вышеописанных методов степень генетических различий плоидности оценивают, используя изначально живые клетки или ткани. Эти ограничения существенны при работе с субфоссильными и музейными образцами. Наиболее доступен в таком случае анализ ДНК (117). Среди большинства известных методов при анализе ДНК главным недостатком остается ее внутривидовая изменчивость, затрудняющая анализ плоидности, например при наличии межвидовой гибридизации (118-120). Анализ коротких повторов ДНК, как правило, лишен этого недостатка, поэтому микросателлитный анализ плоидности ДНК-содержащих тканей таких образцов имеет преимущество. ПЦР-продукты микросателлитных локусов разделяют с помощью гель-электрофореза или капиллярного электрофореза. Анализируя длину фрагментов микросаттелитов можно судить об аллелях локуса и об их количествах (повторов) (см. рис. 4, Г). Как известно диплоидный организм содержит два локуса гомологичных хромосом, поэтому по числу таких локусов возможно определить также и пло-идность (121).
Таким способом плоидность можно диагностировать разными способами в зависимости от задач. Все вышеописанные методы в настоящее время используют в научных исследованиях. Для получения рыб-три-плоидов в индустриальных условиях важно диагностировать триплоидию именно на самых ранних этапах эмбриогенеза (на стадии бластулы), особенно для холодноводных видов рыб, эмбриональное развитие которых продолжается до полугода (124).
Итак, аквакультура как одно из самых быстроразвивающихся направлений сельского хозяйства в мире представляется перспективным способом получения необходимого количества различных питательных веществ, в первую очередь усвояемого белка и полиненасыщенных жирных кислот, омега-3, которые в большом количестве содержатся практически только в водных объектах. Индуцированная полиплоидия, в частности триплоидия, — эффективный и безопасный метод генетической манипуляции в аквакультуре. Полученные организмы не являются ГМО, обладают условной стерильностью, не проходят полового созревания, что позволяет сохранить высокие потребительские свойства мяса, исключает риск гибридизации с природными популяциями. Основными способами получения триплоидов служат блокирование экструзии второго полярного тельца после оплодотворения с помощью теплового шока или гидростатического давления. Тетраплоиды получают подавлением первого митотического деления, однако их хозяйственная ценность невысока, и их применяют главным образом для скрещивания с диплоидами с целью массового получения триплоидов. Перспективны также межвидовые триплоидные гибриды, которые отличаются быстрым ростом и высокими органолептическими качествами. Методы гиногенеза и андрогенеза позволяют полу- чать однополые и стерильные популяции, в том числе «супер-самцов» с ускоренным ростом. Для диагностики плоидности применяют комплекс методов: цитоморфологию крови (устаревший метод), подсчет метафазных хромосом (трудоемкий метод с применением опасных для природы реагентов), проточную цитофлуорометрию (наиболее быстрый, статистически точный метод), а также микросателлитный анализ (пригоден даже для музейных образцов). Экспериментальные данные исследователей показывают, что триплоидная радужная форель превосходит диплоидов по массе и качеству мяса. Выпуски триплоидной радужной форели в водоемы для спортивного рыболовства полностью себя оправдали, а триплоиды атлантического лосося реже заходят в реки, что снижает риск заноса патогенов в природные экосистемы. В России аквакультура пока развита слабо из-за больших естественных запасов биоресурсов, однако наличие научно-исследовательских организаций и уже разработанных методов получения и диагностики полиплоидов позволяет прогнозировать ее рост в будущем. Представленные в обзоре подходы готовы к внедрению в промышленное рыбоводство и обеспечивают экономически оправданную альтернативу традиционным селекционно-племенным работам.
Санкт-Петербургский филиал ФРБНУ ВНИРО ГосНИОРХ им. Л.С. Берга»,
