Полногеномный поиск ассоциаций с показателями экстерьера царскосельской породы кур

Автор: Азовцева А.И., Дементьева Н.В., Рябова А.Е.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Генетика
Статья в выпуске: 1, 2025 года.
Бесплатный доступ
Отрасль птицеводства, обеспечивающая население ценным белком животного происхождения, на сегодняшний день практически полностью зависит от закупки зарубежных кроссов. В связи с этим особое внимание необходимо уделить отечественному птицеводству и имеющимся генетическим ресурсам. Экстерьерная оценка является важным элементом в селекции, т.к. внешний вид позволяет судить о здоровье, породной принадлежности, предрасположенности к продуктивному использованию и приспособленности животного к условиям содержания. Тем не менее, темп селекции зависит от использования как фенотипических, так и генетических данных о животных. Особое внимание в последнее время уделяется GWAS-исследованиям - полногеномному поиску ассоциаций, который позволяет выявить участки генома, предположительно ответственные за проявление того или иного признака. Целью данного исследования было выявить генетические ассоциации с экстерьером у царскосельской породы кур. Для этого была проведена экстерьерная оценка птиц (n = 96), отбор крови и выделение ДНК, а также полногеномное генотипирование при помощи чипа средней плотности Illumina Chicken 60K SNP iSelectBeadChip (Illumina Inc., США). На основании данных полногеномного генотипирования было проведено GWAS-исследование и аннотирование кандидатных генов. В результате было получено 6 предположительно значимых SNP, ассоциированных с обхватом груди, длиной голени, длиной плюсны, а также с углом груди. Бо́льшая часть генов в идентифицированных локусах была вовлечена в процессы костеобразования и костного гомеостаза, которые косвенно регулируют биологический потенциал особи к росту. Выявленные гены-кандидаты могут быть рекомендованы к использованию в маркерной селекции для царскосельской породы кур. Для подтверждения фундаментальной роли выявленных генов в формировании генетического потенциала размерно-весовых характеристик необходимы исследования и на других породах птиц.
Селекция, экстерьерная оценка, птицеводство, промеры
Короткий адрес: https://sciup.org/147251167
IDR: 147251167 | УДК: 575.162 | DOI: 10.17072/1994-9952-2025-1-69-79
Genome-wide association analysis for exterior traits in Tsarskoye Selo chicken breed
To date, poultry industry, which is an indispensable supplier of animal protein, almost entirely depends on foreign commercial crosses. In this regard special attention should be paid to local poultry breeding and available genetic resources. Exterior assessment in an important part of selection, as animal’s appearance is an indication of health, breed affiliation, predisposition to productive use and adaptability to housing conditions. However, the selection rate depends on both phenotypic and genetic data. Recently special attention has been paid to GWAS studies - a genome-wide association search that identifies genome regions presumably responsible for certain traits’ expression. The aim of the study was to identify genetic associations with exterior traits in the Tsarskoye Selo chicken breed. For this purpose, we performed exterior assessment of the birds (n=96), blood sampling and DNA extraction, as well as genome-wide genotyping using Illumina Chicken 60K SNP iSelectBeadChip (Illumina Inc., USA). GWAS analysis and annotation of candidate genes were performed based on genome data obtained. As a result, 6 suggestive SNPs associated with chest girth, tibia length, tarsus length, and chest angle were obtained. Most genes in the identified loci are involved in processes of bone formation and bone homeostasis, which indirectly regulate the biological growth potential of an individual. The identified candidate genes can be recommended for use in marker-assisted selection of Tsarskoye Selo breed. Also, to confirm the fundamental role of the identified genes in formation of body size characteristics, studies on other chicken breeds are needed.
Текст научной статьи Полногеномный поиск ассоциаций с показателями экстерьера царскосельской породы кур
Селекционная работа в животноводстве в той или иной степени подразумевает использование знаний об экстерьере животных – т.е. особенностей их внешнего вида, телосложения. Это обусловлено тем, что экстерьерные данные позволяют судить о здоровье, породной принадлежности, предрасположенности к продуктивному использованию, конституциональной крепости, а также приспособленности к конкретным условиям разведения [Вахрамеев, Макарова, 2021]. Отметим, что экстерьер указывает на продуктивный потенциал животного, а не на фактический уровень продуктивности, который, в свою очередь, зависит как от генотипа животного, так и от влияния антропогенных факторов, в частности условий кормления и содержания. Отсюда следует, что повышение эффективности селекции происходит благодаря улучшению как условий содержания, так и генетики животных. Значительно повысить эффективность селекции позволило повсеместное развитие генетических технологий, способных расшифровать ДНК и установить взаимосвязь между ней и проявлением какого-либо конкретного признака.
Особое значение приобретает селекционная работа в сфере птицеводства, т.к. оно является динамично развивающейся и наукоёмкой отраслью, интенсификация которой требует создания специализированных пород и линий, на основе которых получают высокопродуктивные промышленные кроссы [Гришина, 2021]. В связи с этим в птицеводстве активно используются последние достижения генетики и смежных с ней наук. Особую популярность приобрел полногеномный поиск ассоциаций (Genome-wide association study, GWAS) генетической информации с фенотипическими признаками, который выявляет однонуклеотидные полиморфизмы (single nucleotide polymorphism, SNP) с высокой частотой встречаемости у особей со схожими значениями признака. На сегодняшний день существуют исследования, посвященные геномным ассоциациям с показателями яичной продуктивности [Kang et al., 2024; Yang et al., 2024], длиной шпор у петухов [Chen, Zhao, Zhao et al., 2024], цветом скорлупы яиц и пигментацией голени [Wang X.G. et al., 2024], продолжительностью оплодотворяемости кур-несушек [Luo et al., 2024], формой клюва [Chen, Zhao, Wen et al., 2024], качественными характеристиками мяса разных пород [Xiao et al., 2024], конверсией корма и среднесуточными приростами [Cai et al., 2024], крепостью костяка [Азов-цева, Дементьева, 2023], ростом и развитием в породах разных направлений продуктивности [Ларкина и др., 2021], а также с окрасом оперения [Hua et al., 2021] и др.
Царскосельская порода кур является результатом 30-летнего труда научных сотрудников ВНИИГРЖ, целью которых было выведение аутосексной породы комбинированного направления продуктивности с приятным декоративным окрасом [Вахрамеев и др., 2024]. Комбинированное направление характеризуется оптимальными значениями яйценоскости при сохранении хороших мясных характеристик, и это, наравне с экспоненциальным ростом интереса потребителей к органическим продуктам питания [Ales-sandroni, Sagratini, Gagaoua, 2024], делает царскосельскую породу перспективным объектом выращивания для фермеров. Тем не менее, успешное развитие породы во многом зависит от наличия интереса промышленности к ней. В большинстве промышленных предприятий РФ для производства мяса и яйца птицы используются зарубежные кроссы [Федорова, Станишевская, Дементьева, 2020], что обеспечивает рентабельность производства, но пагубно сказывается на сохранении биоразнообразия и на уровне развития отечественного птицеводства, а также создает угрозу для продовольственной безопасности страны [Буяров, Буяров, 2022]. Целесообразной является деятельность по внедрению российских пород в промышленное производство, однако для этого необходимо предоставить доказательства их практической и племенной ценности. Как упоминалось ранее, экстерьер является хорошим индикатором здоровья, потенциального уровня продуктивности и приспособленности к условиям содержания. В связи с этим целью данного исследования стало выявление геномных ассоциаций с показателями экстерьерной оценки царскосельской породы и аннотирование генов-кандидатов экстерьерных признаков.
Материалы и методы
В качестве материала для исследования была использована птица царскосельской породы (n = 96), содержащаяся в ЦКП «Генетическая коллекция редких и исчезающих пород кур» (ВНИИГРЖ, г. Санкт-Петербург – Пушкин). Для проведения исследования в возрасте 42 недель был проведен прижизненный сбор фенотипических данных птицы и отбор проб крови для выделения ДНК. Сбор фенотипических данных заключался в проведении экстерьерной оценки со следующими показателями: живая масса (ЖМ), обхват груди (ОГ), обхват плюсны (ОП), косая длина туловища (КДТ), длина корпуса и шеи (КШ), длина киля (ДК), длина бедра (ДБ), длина голени (ДГ), длина плюсны (ДП), глубина груди (ГГ), ширина в ключицах (ШК), ширина таза (ШТ), угол груди (УГ).
Выделение ДНК из крови проводилось по стандартной методике фенол-хлороформной экстракции. Определение концентрации и чистоты ДНК образцов проводилось методом спектрофотометрии при помощи прибора NanoDrop 2000c (Thermofisher Scientific Inc., США). Полученные образцы были отправлены на полногеномное генотипирование с использованием чипа Illumina Chicken 60K SNP iSelectBeadChip (Illumina Inc., США) с плотностью покрытия 57,636 SNP. Контроль качества генотипирования проводился при помощи программы Genome Studio (Illumina Inc., США). Образцы с качеством генотипирования более 95% были использованы в дальнейшем анализе. Редактирование полученных данных для создания файлов адаптивного расширения (.ped, .map, .fam, .bed, .bim) было проведено при помощи программного обеспечения PLINK 1.9. с частотой минорных аллелей (MAF) > 0.05. В результате применения данного фильтра количество однонуклеотидных полиморфизмов, участвующих в анализе, сократилось до 43,123 SNP.
Полногеномный поиск ассоциаций генетической информации с продуктивными признаками был проведен при помощи статистического программного обеспечения EMMAX. Для этого в EMMAX была сгенерирована матрица родства «идентичность по штатам» («identity-by-state»). Влияние SNP на признак было рассчитано в соответствии с моделью:
Y = Xb+u+e , где Y – вектор фенотипов; b – эффект SNP; X – расчётная матрица генотипов SNP, u – вектор аддитивных генетических эффектов, который считается нормально распределенным со средним значением, равным 0, и (ко)дисперсией σ2aG , где σ2a – аддитивная генетическая дисперсия, а G – матрица геномных отношений; e — вектор случайных остаточных эффектов.
С целью исключения ложноположительных результатов и установления уровней значимости для эффекта SNP была применена поправка Бонферрони. Достоверно значимый и предположительно значимый уровни были установлены как 1,16E-06 (0.05/43,123) и 2,31E-05 (1.00/43,123) соответственно. Общегеномная значимость оценена при помощи метода simple в R, а расчет эффективного числа независимых тестов – с помощью программы Meff. На основании результатов GWAS с использованием пакета qqman и ggplot2 в среде программирования R построены диаграммы типа Манхэттэн и графики «квантиль-квантиль» (Q-Q plot); при помощи пакета corrplot – корреляционная матрица для изучаемых признаков.
Гены, совпадающие с геномным регионом кандидатного SNP или близкие к нему, были аннотированы в геномном браузере ENSEMBL на основании сборки генома Chicken (Red Jungle Fowl) GRCg6a. Поиск информации по функциональным характеристикам кандидатных генов был осуществлен в базе данных NCBI.
Результаты и обсуждение
В результате проведения полногеномного анализа ассоциаций для прижизненно полученных фенотипических показателей было идентифицировано 6 предположительно значимых SNP (таблица). Из 12-ти анализируемых показателей ассоциации были получены только для 4, что, вероятно, обусловлено наличием высоких корреляционных связей между большинством признаков (рис. 1).
Для показателя ОГ был получен SNP rs312492432 (1,73E-05), локализованный в интронной части гена с неустановленной функцией (рис. 2 А).
Предположительно значимые SNP, ассоциированные с показателями экстерьера
[Suggestive SNPs associated with exterior traits]
ДГ 0.84 0.77 q
ИМ 0.77 0.7S 0.76 1
КДТ 0.82 0.77 0.76 0.68 S'
ДБ 0.72 0.72 0.68 0.64 ■ Ct
ОГ 0.04 0.67 0.68 0.76 0.57 0.03 О
ДК ■■143 0.50 0.56 0.53 0.52 § _ кш ■■■■ИВ ’41 *1 §
-
™ ЯВДВ^РРИД
ГГ 0Л9 Ml 0.42 0.33 0.37 0.38 0.34 0.48 0.27 0.37 lZ
ШТ 0.33 0.36 0.3 0.5 0.25 0.25 0.49 0.24 0.24 0.37 0.11 g
УГ 0.16 0.14 0.13 0.21 0.16 0.13 0.05 0.12 0.24 0.14 0.03 0.12
^......м
-
-1 -0.8 -0.6 -0.4 -0.2 0 0.2 0.4 0.6 0.8 1
Рис. 1 . Корреляционная матрица для показателей экстерьерной оценки царскосельской породы кур: ДП – длина плюсны, ОП – обхват плюсны, ДГ – длина голени, ЖМ – живая масса, КДТ – косая длина туловища, ДБ – длина бедра, ОГ – обхват груди, ДК – длина киля, КШ – длина корпуса и шеи, ШК – ширина в ключицах, ГГ – глубина груди, ШТ – ширина таза, УГ – угол груди [Correlation matrix of the exterior assessment traits of the Tsarskoye Selo chicken breed:
ДП is metatarsal length, OП – metatarsal girth, ДГ – shin length, ЖМ – body weight, KДТ – oblique body length, ДБ – thigh length, OГ – chest girth, ДК – keel length, KШ – the neck and body length, ШK – clavicle width, ГГ – chest depth, ШТ – pelvis width, УГ – chest angle]
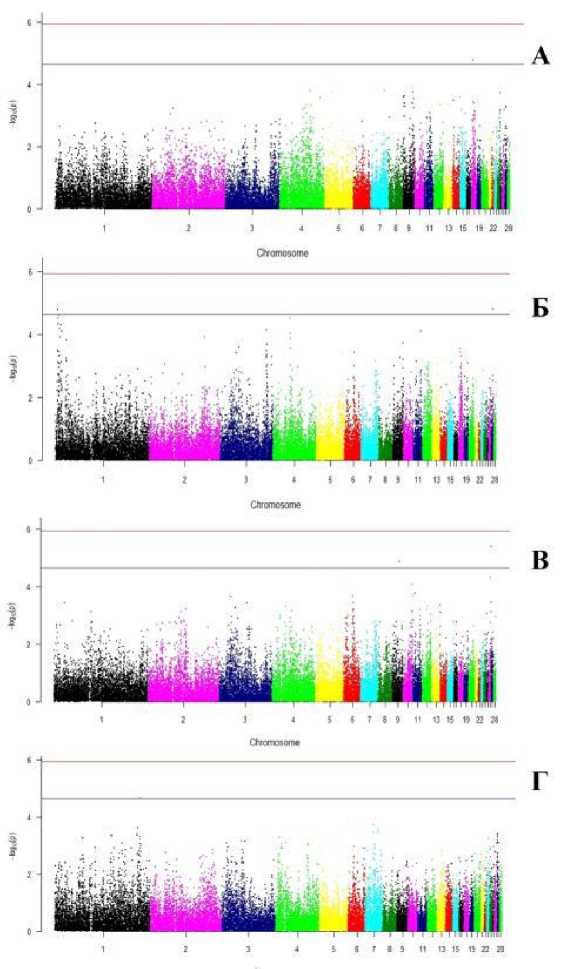
Рис. 2 . Графики Manhattan для обхвата груди (А), длины голени (Б), длины плюсны (В), угла груди (Г) царскосельской породы кур
[Manhattan plots for chest girth (A), shin length (Б), metatarsal length (В), chest angle (Г) of the Tsarskoye Selo chicken breed]
В пределах 0,3 Мб от целевого SNP расположен ген FOXK2 – регулятор транскрипции, который участвует в различных процессах, включая гликолиз, аутофагию и метаболизм глюкозы. В предыдущих исследованиях было отмечено его участие в сигнальном пути Wnt/β-catenin [Wang W. et al., 2015], который необходим для эмбрионального развития и регенерации гомеостаза костных тканей [Duan, Bonewald, 2016; Liu J. et al., 2022]. Эта информация позволяет объяснить наличие положительных корреляций ОГ с показателями развития костей, такими как ОП, ДП и ДБ (рис. 1). В другом исследовании обнаружено, что ген FOXK2 модулирует путь Wnt, который, как известно, связан с миогенезом, дифференцировкой типов мышечных волокон и рекрутированием сателлитных клеток после травмы [Petrie et al., 2016]. Все вышеперечисленное позволяет предположить, что сигнальный путь Wnt играет ключевую роль в эмбриональном развитии особи и, соответственно, обуславливает ее генетический потенциал роста, а ассоциация FOXK2 с ОГ обусловлена зависимостью данного показателя от количества и размеров мышечных волокон груди.
С показателем ДГ ассоциирован SNP rs13823757 (1,62E-05), локализованный в интронных частях сразу трёх РНК-кодирующих генов, а также SNP rs14707919 (1,56E-05), в пределах 0,1 Мб от которого рас- положены гены IGF2BP1, GIP, PHOSPHO1 и PHB1 (рис. 2 Б). Ген IGF2BP1 кодирует член семейства мРНК-связывающих белков инсулиноподобного фактора роста 2. Ранее этот ген был ассоциирован с массой и прочностью большеберцовой кости у уток породы Нонгхуа [Lu et al., 2024]. Ассоциация этого гена одновременно с ДГ и с показателями бедренной кости вполне логичны, т.к. для показателя ДГ обнаружены высокие положительные корреляции со всеми морфометрическими показателями задних конечностей – ДБ, ДП и ОП (рис. 1). В другом исследовании выявлено, что этот ген участвует в сигнальном пути Hedgehog и регулирует стабильность РНК нескольких генов-мишеней, связанных с ростом [Noubissi et al., 2018], и это также согласуется с положительной корреляцией ДГ с ЖМ и КДТ. Более того, в исследовании на козах, indel-мутации в гене IGF2BP1 также были ассоциированы с ростовыми показателями [Wang Z. et al., 2020]. Ген GIP – глюкозозависимый инсулинотропный полипептид – кодирует гормон инкретин и принадлежит к суперсемейству глюкагонов. Ранее установлено, что GIP играет роль в регуляции метаболизма костей, а именно подавляет процесс резорбции костей и активно стимулирует костеобразование [Gaudin-Audrain et al., 2013; Skov-Jeppesen et al., 2021; Gabe et al., 2022; Liu H. et al., 2024]. Ген PHOSPHO1 кодирует фосфатазу, которая предположительно участвует в минерализации и созревании костей. Экспрессия PHOSPHO1 значительно выше в минерализованных тканях и обнаружена у ряда видов, включая людей и мышей [Morcos et al., 2018]. Согласно имеющимся данным, PHOSPHO1 играет важную роль в восстановлении после переломов, в связи с чем может быть использован в качестве терапевтической мишени при их лечении [Dillon et al., 2019]. Мыши с дефицитом этого гена характеризуются аномальными пластинками эндохондрального роста и аномалиями скелета, включая остеомаляцию, сколиоз, деформацию длинных костей и спонтанные переломы [Yadav et al., 2011; Javaheri et al., 2015]. В исследованиях на птицах также выявлено, что подавление активности PHOSPHO1 приводит к нарушениям минерализации скелета во время развития конечностей цыпленка [Macrae et al, 2010]. PHB1 является эволюционно консервативным геном, предположительно участвующим в клеточном старении и подавлении опухолей. В исследовании Tabti с соавт. отмечено, что PHB1 вовлечен в этиологию диабета, ожирения и остеопороза [Tabti et al., 2021].
В исследовании ростовых показателей кур сразу три гена, IGF2BP1 , GIP и PHOSPHO1 , были предложены в качестве генов-кандидатов роста костей [Wang Y. et al., 2020], что согласуется с результатами, полученными в нашем исследовании. На основании полученных ассоциаций и корреляций можно выдвинуть предположение, что локус на 27 хромосоме, содержащий вышеперечисленные гены, играет важную роль в формировании размерно-ростовых показателей особей, в особенности конечностей.
Для показателя ДП было аннотировано 2 SNP – rs16204837 (4,01E-06) и rs315156385 (1,36E-05) на 26 и 9 хромосомах соответственно (рис. 2 В). SNP на 26 хромосоме был локализован в интронной части гена TAF8 , кодирующего один или несколько факторов, связанных с TATA-связывающим белком. TAF8 является белком развития, который вовлечен в процесс транскрипции и её регуляции. В исследовании Wong et al. (2022) для гена TAF8 описано 7 случаев мутаций, приводящих к нейродегенеративным заболеваниям у людей, которые сопровождаются задержкой роста, микроцефалией, прогрессирующей спастичностью, церебральной атрофией и развитием эпилепсии [Wong et al., 2022]. В пределах 0,5 Мб от целевого SNP на 26 хромосоме также расположены гены BTG2 , FMOD и PREPL . Ген BTG2 кодирует антипроли-феративный белок, участвующий в процессе транскрипции и ее регуляции. В исследовании Park et al. (2004) на мышах выявлено, что BTG2 усиливает транскрипционную активность передачи сигналов BMP (костный морфогенетический белок), а обнаруженные динамические паттерны экспрессии BTG2 в развивающихся сомитах доказывают его участие в формировании паттерна позвонков [Park et al., 2004]. Ген FMOD кодирует фибромодулин и предположительно играет основную роль в фибриллогенезе коллагена. FMOD детектируется в эмбриональных склеротомах уже на 10 день после оплодотворения, а также экспрессируется хондроцитами и остеобластами плода во время эндохондрального и внутримембранозного окостенения [Gori, Schipani, Demay, 2001; Tevlin et al., 2023]. Особое значение для формирования и прочности кости FMOD имеет благодаря его способности опосредованно облегчать сшивание коллагена [Hudson et al., 2017]. PREPL кодирует пропилэндопептидазу и представляет собой матричный белок, экспрессируемый в базальных мембранах, хрящах и костном матриксе [Rucci et al., 2013]. Он обладает мощным антирезорбтивным действием, ингибируя образование остеокластов, в результате чего был рекомендован в качестве нового агента для лечения заболеваний костей, вызванных повышенной активностью остеокластов [Rucci et al., 2013].
SNP на 9 хромосоме был локализован в интронной части гена DCUN1D1, который обеспечивает связывающую активность белков семейства куллинов. DCUN1D1 играет важную роль в убиктинировании и неддилировании белков – процессах пострансляционной модификации, которые регулируют стабильность и функцию целевых белков [Paccez et al., 2024]. В пределах 0,3 Мб от целевого SNP расположен ген FETUB – член семейства фетуинов, которые участвуют в остеогенезе и резорбции кости, регуляции рецепторов инсулина и фактора роста гепатоцитов, а также реакции на системное воспаление. Отмечено, что фетуины включаются в костную ткань через сыворотку крови при кальцификации, при этом пониженные концентрации фетуина в плазме крови характерны для больных болезнью Педжета, а высокие уровни – у больных с несовершенным остеогенезом [Kubota et al., 2012]. Вся группа генов, ассоциированных с ДП, задействована либо в остеогенезе, либо в костном гомеостазе, и наличие высоких корреляционных связей между показателями ДП, ДГ и ДБ подтверждает влияние выявленных локусов на скелет конечностей.
Для последнего изучаемого показателя – УГ – был идентифицирован SNP rs316245165 (2,27E-05) на 1 хромосоме, локализованный в интроне РНК-кодирующего гена с неустановленной функцией (рис. 2 Г). Для установления функциональной характеристики и влияния данного SNP на показатель УГ необходимы дополнительные исследования.
Заключение
В результате проведения настоящего исследования для царскосельской породы кур было получено 6 предположительно значимых SNP, ассоциированных с показателями экстерьерной оценки – обхватом груди (1 SNP), длиной голени (2 SNP), длиной плюсны (2 SNP) и углом груди (1 SNP). Ген FOXK2 , ассоциированный обхватом груди, включен в группу высоко консервативных сигнальных путей Wnt. Согласно существующим данным, пути Wnt задействованы в процессах эмбриогенеза и миогенеза, а также развития и регенерации костных тканей, что объясняет как полученную ассоциацию гена FOXK2 с обхватом груди, так и положительную корреляцию последнего с показателями развития конечностей (обхват плюсны, длины плюсны и голени). Гены IGF2BP1, GIP, PHOSPHO1 и PHB1 были ассоциированы с длиной голени. Известно, что все перечисленные гены вовлечены в процессы костеобразования, а ген IGF2BP1 ранее был ассоциирован с ростовыми показателями как представителей класса птиц, так и других сельскохозяйственных животных. Эти данные позволяют предположить, что локус на 27 хромосоме, содержащий перечисленные гены, играет важную роль в формировании размерно-ростовых показателей особей, в особенности конечностей. С показателем длина плюсны ассоциированы два региона на 26 и 9 хромосомах. На хромосоме 26 идентифицированы гены TAF8, BTG2, FMOD и PREPL . Согласно данным, мутации в TAF8 вызывают нейродегенеративные заболевания, сопровождающиеся задержкой роста, тогда как BTG2, FMOD и PREPL задействованы в процессах костеобразования и гомеостаза костной ткани. На 9 хромосоме расположены гены DCUN1D1 и FETUB. Влияние гена DCUN1D1 на длину плюсны может обуславливаться способностью первого участвовать в процессах посттрансляционной модификации белков. Ген FETUB же достоверно участвует в процессах остеогенеза и резорбции костей, и, более того, отклонения концентраций фетуинов от нормы наблюдаются у пациентов с костной болезнью Педжета и с несовершенным остеогенезом. Весь кластер генов, ассоциированных с длиной плюсны, задействован либо в остеогенезе, либо в костном гомеостазе, и наличие высоких положительных корреляционных связей между показателями длина плюсны, длина голени и длина бедра подтверждает влияние выявленных локусов на скелет конечностей. Также выявлена ассоциация между углом груди и РНК-кодирующим геном с неизвестной функцией, в связи с чем установление функциональных характеристик данного гена требует дополнительного изучения.
Проведенное исследование позволило выявить ряд локусов, ассоциированных с экстерьерными характеристиками царскосельской породы кур. Бо́льшая часть генов в идентифицированных локусах была вовлечена в процессы костеобразования и костного гомеостаза, которые косвенно регулируют биологический потенциал особи к росту. Выявленные гены-кандидаты могут быть рекомендованы к использованию в маркерной селекции для царскосельской породы кур. Для подтверждения фундаментальной роли выявленных генов в формировании генетического потенциала размерно-весовых характеристик необходимы исследования и на других породах птиц.
