Половая структура популяций и семенная продуктивность Stachys palustris L. s. l. на Урале

Автор: Демьянова Е.И.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 1, 2017 года.
Бесплатный доступ
У чистеца болотного (Stachys palustris L. s.l., Lamiaceae) изучена половая структура популя-ций, антэкологические особенности и семенная продуктивность на Урале (в Приуралье и За-уралье). Это насекомоопыляемое ксеногамное растение. При изоляции семена не завязывают-ся. В Пермском крае у разноцветковых форм обнаружена различная семенная продуктив-ность. Она более высокая у белоцветковой формы по сравнению с пурпуровоцветковой. Дан-ный феномен требует дальнейшего исследования.
Половая структура популяций, антэкологические особенности, семенная продуктивность
Короткий адрес: https://sciup.org/147204803
IDR: 147204803 | УДК: 581.162
The sex structure of populations and seed productivity of Stachys palustris L. s. l. in the Urals
We studied the sexual structure of populations, anthecological features and the seed production of Stachys palustris L. s.l., Lamiaceae in the Urals (also the Priuralie and Trans-Urals). This is en-tomophilous xenogamic plant. Seeds are not appeared in isolation. In Perm region the plant forms with different flowers show different level of seed production. It is higher in plants with white flowers than in plants with purple flowers. This phenomenon should be studied more.
Текст научной статьи Половая структура популяций и семенная продуктивность Stachys palustris L. s. l. на Урале
Одной из распространенных половых форм среди цветковых растений является гинодиэция (женская двудомность), впервые исследованная Ч. Дарвином [Darwiiu 1877J. Популяции гинодиэцич-ных видов состоят из гермафродитных особей и андростерильных с функционально женскими цветками. Гинодиэция* по-видимому, возникла независимо в ходе эволюции цветковых растений. Она встречается в разных порядках и семействах, филогенетически часто не связанных друг с другом [Пономарев* Демьянова* 1975; Демьянова* Пономарев* 1979; Демьянова, 1985: Годин, Демьянова, 2012]* Подавляющее большинство из них занимает верхнее положение в системе цветковых растений [Тахтаджян* 1970; 1986]. Как правило, им свойственна зрелая трехклеточная пыльца.
Как уже указывалось [Демьянова, 1981], особенно чётко такая взаимосвязь гинодиэции со строением зрелой пыльцы просматривается у La-mi асе не, Женская двудомность свойственна в основном таксонам со зрелой трёхклеточной шестибороздной пыльцой* нежели двуклеточной трёхбороздной* В последнем случае при отсутствии или минимальном числе женских особей в популяции изучаемого вида нередко отмечается гиномоноэция - менее резкое разделение полов. При гиномоно-эции обоеполые и женские цветки находятся в пределах одной особи. Как правило* первые явно преобладают, а число андростерильных (женских) цветков сравнительно невелико. К таким растениям относятся изученные нами виды Stachys L*: S sylvatica L* У recta L., & pains tris L. У перечис-
(С Демьянова E . И * 2017
ленных видов гораздо чаще фиксируется гиномо-ноэция в сравнении с гинодиэцией* Ранее [Демьянова, 1981] нами указывалось полное отсутствие гинодиэции у Skichys paiustris L* Позднее у этого вида в 1987 г. в заказнике «Предуралье» (Пермский край) обнаружено 3 женских особи (из просмотренных 1015 растений, что составляет 0.2%), а в 1990 г. в Троицком лесостепном заказнике (Челябинская область) найдено также 3 женских особи (из 843 учтенных растений (0*4%)* Таким образом, популяции чистеца болотного, как и у ранее изученных нами видов Stachys Ц* состоят из гермафродитных, гиномоноэцичных и крайне малой доли женских особей.
Судя по литературе, многим видам Stachys L. присущ полиморфизм, проявляющийся в межвидовой и внутривидовой изменчивости [Кнорринг, 1954: Дервиз-Соколова, 1975: Mulligan at al, 1983: Кочиева и др , 2006; Русских, 2009]. В подобной ситуации таксономические отношения в пределах рода довольно запутанны и неопределённы*
Что касается семенного размножения* то у изученных видов чистеца (-S', recta L* £ siebotdii Miq*) обнаружена низкая семенная продуктивность [Старшова* 1992; Чоркина и др** 1996 а*б]. Авторы полагают, что важной причиной невысокой семенной продуктивности являются нарушения в мегаспорогенезе и развитии зародышевых мешков.
Объектом настоящего исследования явился чистец болотный (Stachys paiustris L. s.L), изученный в природных популяциях Урала (в Приуралье и Зауралье). Это лекарственное растение с терапевтическим эффектом* медоносное* жирномасличное, сорное [Костюченко, 1983; Дикорастущие.*.* 2001; Алексеева, Канев* 2014 и др*]*
Чрезвычайно полиморфный вид с очень широким ареалом, включающим Европу, Азию* Сев. Америку [Кнорринг, 1954; Mulligan at al., 1983]. Размножается семенным и вегетативным путем. В Пермском крае (Приуралье) встречается на низинных лугах и болотах* по берегам водоемов, а также как сорное [Иллюстрированный .., 2007]. В Зауралье в Челябинской области отмечается примерно в таких же условиях [Куликов, 2010].
Энтомофильное растение с обширным кругом опылителей. Благодаря короткой трубке венчика может опыляться как длиннохоботными, так и короткохоботными насекомыми* Основными опылителями являются шмели и пчелы. Судя по литературе, наиболее полный список опылителей чистеца болотного в рамках Европы приведен в монографии П. Кнута [Knuth, 1899]* Обширный перечень опылителей и посетителей этого растения включает представителей разных отрядов - Hymenoptera* Diptera, Lepidoptera, Coleoptera. По свидетельству этого автора, несмотря на разнообразие опылителей, кроме перекрестного опыления у чистеца болотного возможно и спонтанное самоопыление.
Задача нашего исследования - изучение поло вой структуры популяций чистеца болотного, антэ-кологин и семенной Продуктивности в условиях Урала. Особый интерес представляло изучение семенной продуктивности у разноцветковых форм (особей, имеющих белые или обычные пурпуровые цветки) в Пермском крае* Литературные сведения по подобной тематике нами не обнаружены.
Организация и методика исследований
Исследования по антэкологии и семенной продуктивности чистеца болотного проводились в Троицком лесостепном заказнике (1986-1990 и 2010 гг.) в ценопопуляциях ио берегу оз. Ку кай, а в Пермском крае (Нытвенский р-он) в посадках картофеля (2011-2016 гг).
Половая структура популяций (соотношение полов) определялась во время массового цветения путем визуального просмотра цветков на всех одновременно цветущих особях* Учет растений проводился методом ходовых линий. При этом просматривались все растения подряд, независимо от их обилия. Подсчеты, осуществленные в отдельных ценопопуляциях (обычно 5-7)* затем суммировались для географической популяции в целом.
Антэкологические исследования проводились по методике А.Н. Пономарева [1960]. При изучении фертильности пыльцы мы пользовались общепринятой ацетокарминовой методикой [Паушевач 1974, Справочник..., 2004].
Семенная продуктивность изучалась согласно указаниям ТА. Работнова [1960] и И.В. Вайнагия [1973* 1974]* Для анализа семенной продуктивности нами в разные годы просматривалось обычно от 500 до 2000 цветков* реже до 100* в которых учитывалось количество завязавшихся семян (эре-мов)* Раздельно подсчитывались как выполненные, так и невыполненные эремы. К последним мы относили мелкие, щуплые и поврежденные семена [Левина, 1970].
Для всех губоцветных потенциальная семенная продуктивность в расчете на цветок (ПСП - число семяпочек в завязи) постоянна и равна 4* Ежегодно определялась РСП - среднее количество завязавшихся семян* приходящихся на элементарную единицу семенной продуктивности - цветок). Кроме ПСП и РСП, устанавливался и процент семе-нификации (ПС) как отношение количества развившихся семян к количеству семяпочек. ПС свидетельствует об эффективности завязывания семян [Вайнагий* 1974]. Поскольку у этого вида женские особи встречаются чрезвычайно редко, подсчет семенной продуктивности производился лишь для обоеполых цветков.
Собранный материал подвергся статистической обработке в соответствии с методикой В.Ф. Лакина [1973, 1990] и Н А. Плохинского [1978]. Данные о метеоусловиях заимствованы на сайте «Гисметео» [].
Результаты и их обсуждение
Половая структура популяций. Как уже сообщалось, популяции чистеца болотного представлены гермафродитным IL ГИНОМОНОЭЦИЧНЫМИ и женскими особями. Последние, судя по многолетним наблюдениям, выявляются не во все годы наблюдений даже при достаточно большой выборке материала (от нескольких сотен до нескольких тысяч просмотренных особей).
Разные ценопопуляции даже в пределах одного пункта наблюдений могут быть неодинаковы по половому составу. Так, например, в Троицком лесостепном заказнике в 1989 и 1990 гг. исследовалась половая структура двух ценопопуляции: первая находилась по берегу пресноводного оз. Кукай, а вторая - в Солёном логу (по берегу солоноватого озера). Во второй ценопопуляции в оба года исследований нс было обнаружено ни женских, ни гиномоноэцичных особей. Возможно, что подобная ситуация может быть объяснена формированием данной ценопопуляции в основном с помощью вегетативного размножения. Здесь растения произрастали очень скученно, в то время как для цено-популяцни по берегуг оз. Кукай было характерно разреженное произрастание особей. В состав последней ценопопуляции входили гермафродитные, гиномоноэцичные и крайне немногочисленные женские особи.
Гиномоноэцичные и гермафродитные особи отличаются по строению соцветий. У гермафродитных растений соцветие более рыхлое, мутовки дальше отставлены друг от друга, чем у гиномоноэцичных особей. У последних верхняя часть соцветия, где по большей части расположены женские цветки, сильно уплотнена по сравнению с серединой соцветия с преимущественной локализацией обоеполых цветков. Женские особи имели очень плотные и короткие соцветия,
Динамика цветения. Для чистеца болотного характерно длительное цветение. Так, в условиях Зауралья цветение этого вида наблюдается с середины июля до конца сентября. Примерно в такие же сроки (в зависимости от погодных условий) оно отмечается и в Пермском крае.
Анализ динамики цветения, изученной в 1990 г. у ценопопуляции по берегу оз. Кукай, позволяет утверждать, что в начале цветения у этого вида преобладают гермафродитные особи. Затем (примерно через 2 недели) начинают активно цвести гиномоноэцичные растения. После массового цветения гермафродитных особей увеличивается и становится максимальным число цветущих гино-моноэцнчных растений. Цветение женских особей отмечено во время максимального цветения гермафродитных. Последние цветут примерно 15-19 дней (табл. 1.).
Таблица 1
Динамика половой структуры ценопопуляции чистеца болотного в 1990 г, (по берегу оз, Кукай)
|
Дата наблюдений1 |
Половая форма |
||
|
гермафродитные, шт. |
гиномоноэцичные. шт. |
женские, шт. |
|
|
15.07.90 |
зо |
4 |
О |
|
17,07,90 |
102 |
52 |
0 |
|
20.07.90 |
174 |
50 |
1 |
|
22,07,90 |
155 |
101 |
2 |
|
24.07.90 |
89 |
125 |
0 |
Примсчашк, 1 В каждый срок наблюдет ши вес растения ценопопуляции пересчитывались заново и определялся их половой статус.
У гиномоноэцичных особей наблюдается определенная последовательность в распускании обоеполых и женских цветков в пределах соцветия. Сначала распускаются обоеполые цветки, расположенные в нижних и средних мутовках. Женские цветки, обычно находящиеся в верхних мутовках.
как правило, раскрываются ко времени окончания цветения особи. Их число на ГИНОМОНОЭЦИЧНЫХ особях весьма неодинаково. Так. при анализе 30 модельных экземпляров в 1990 г. их численность колебалась от 15 до 70 цветков (табл. 2).
Таблица 2
Динамика цветения гиномоноэцичных особен чистеца болотного в Троицком заказнике в 1990 г, (п=30)
|
Дата наблюдений |
Число цветков на одной особи |
|
|
обоеполые |
женские |
|
|
15.07.90 |
20.10 ± 3.46 |
0 |
|
1607.90 |
28.30 ±201 |
5 .10 ± 1 02 |
|
18,07,90 |
25,60 ± 1,29 |
12.40 ±0.63 |
|
20.07.90 |
17.50 ±2.23 |
48.70± 135 |
|
22,07,90 |
5.00 ± 3,21 |
68.20 ± 9.63 |
|
24.07.90 |
единичны |
72.10 ± 10.22 |
Суточная ритмика распускания обоеполых цветков Она изучалась неоднократно в Троицком заказнике в 1987-1990 гг. Во все годы наблюдений зафиксирован утренний тип раскрывания цветков В отдельные июльские дни с высокими дневными температурами раскрывание цветков начиналось на рассвете и продолжалось до 14-16 ч. Массовое распускание цветков наблюдалось между 9-12 ч. (рисунок)
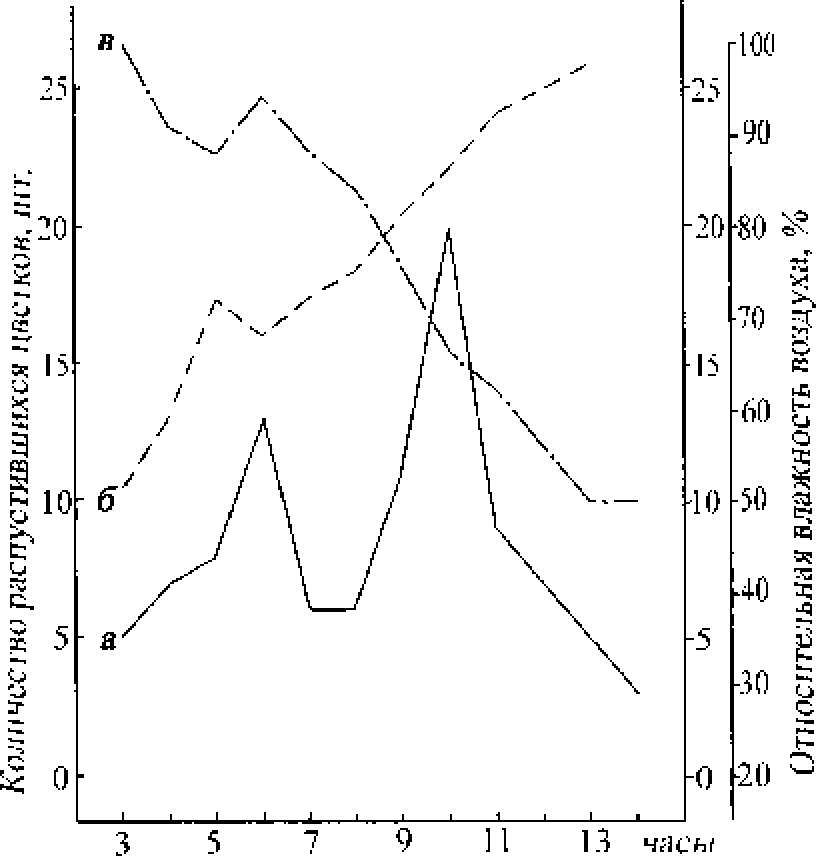
Суточный ритм распускания цветков Siachys paluslns L (2 5 07.1987):
а - количество распустившихся цветков; б -температура воздуха, в - относительная влажность воздуха
Антэ кол огня обоеполых цветков. Некрупные цветки чистеца болотного явно энтомофильного облика. Нижняя губа венчика с темным рисунком («указатели» нектара) Она шире и длиннее верхней и служит посадочной площадкой для опылителей. В стадии рыхлого бутона светло-коричневые пыльники обоеполых цветков расположены выше рыльца, лопасти которого еше сомкнуты. Ко времени полного раскрывания венчика начинается тычиночная стадия Пыльники приобретают красноватый оттенок. На первом этапе тычиночной стадии активно выделяет пыльцу нижняя пара тычинок (длинные тычинки). Пыльники вскрываются продольной шелью интрорзно. Они выдвигаются из трубки околоцветника и раздвигаются в стороны. Когда эти тычинки начинают подсыхать, экспонировать пыльцу начинает верхняя пара тычинок (короткие тычинки), знаменуя начало второго этапа тычиночной стадии. Через сутки нижняя пара тычинок окончательно завядает, а еще через 6-8 ч. засыхает и верхняя пара тычинок Общая про- должительность тычиночной стадии - 30—32 ч. Во время тычиночной стадии рыльце находится внутри трубки венчика ниже тычинок. Столбик рыльца начинает расти только через 29—30 ч, после открывания цветка. Через 50-52 ч, после распускания цветка оно выдвигается из околоцветника и рас-крывает лопасти (вильчатая стадия). К концу третьих суток цветения околоцветник начинает подвядать, но рыльце остается еще жизнеспособным. На четвертые сутки венчик окончательно завядает, а рыльце, судя по его тургору, еще 1-2 ч, остается жизнеспособным.
Следовательно, хотя дихогамия выражена сравнительно слабо, тычиночная и рыльцевая стадии обоеполого цветка почти не налагаются друг на друга, причем в распустившемся цветке репродуктивные органы бывают пространственно разделен ы
Тем не менее, отмечено, что последовательность в прохождении стадий цветения обоеполых цветков чистеца болотного может быть и иной Вскрывание пыльников длинных тычинок может происходить в одних цветках в стадии рыхлого бутона, у других позднее - ко времени распускания цветка (см. выше), у трет ьих — через 1—2 ч. после раскрывания цветка. Таким образом, длительность тычиночной стадии в цветках чистеца болотного неодинакова у разных особей.
Бутоны женских цветков более плотные по сравнению с обоеполыми. При распускании женского цветка нижняя губа лишь слегка отходит от верхней Их цветение начинается сразу же с рыль-цевой стадии. При этом рыльце слегка вытягивается за пределы околоцветника в узкую щель между нижней и верхней губами, после чего его лопасти раскрываются В этом состоянии цветок находится на протяжении всего периода цветения Венчик начинает засыхать спустя 50-52 ч. после распускания цветка, но рыльце сохраняет свою жизнеспособность еще 2-3 ч, Женский цветок у гиноди-Эцичных видов, будучи функционально однополым, является потенциально обоеполым [Кордюм, Глущенко, 1976] В женских цветках чистеца болотного обнаружены тычинки разной степени дегенерации
Фертильность пыльцы У обоеполых цветков чистеца болотного выявлена высокая фертильность пыльцы (табл. 3). Особи, имеющие низкую фертильность пыльцы, единичны. Высокая фертильность пыльцы (70-90%) была отмечена и в другие годы наблюдений в Троицком заказнике (в 1987, 1988, 2010 гг.). Аналогичные данные получены и в Пермском крае Так, в 2012 г. фертильность пыльцы у пурпуровоцветковой формы оказалась равной 90.7, а у белоцветковой 91.4%. в 2016 г соответ- ственно 78.8 и 83*4% (из 20 измерений той и другой формы)*
Семенная продуктивность обоеполых цветков при свободном опылении и изоляции. В Зауралье во все годы исследований (1986-1990 и 2010 гт.) семенная продуктивность при свободном опылении была крайне неодинаковой, о чем свидетельствуют ее показатели (табл. 4). Обнаружена определенная зависимость между ними и погод ными условиями. В сухие по сумме осадков годы (1986* 1989 гг*) процент семенификации (ПС) и число эремов на плод были гораздо ниже* чем во влажные 1987 и 1990 гг. Данные результаты хорошо согласуются с показателями структуры семенной продуктивности (табл* 5); число цветков* завязавших 4 эрема* в сухие годы оказывается наименьшим. При изоляции растений семена не завязываются*
Таблица 3
Варьирование фертильности пыльцы у обоеполых цветков чистеца болотного в Троицком лесостепном заказнике
|
Год наблюдений |
Число исследованных растений |
Показатели |
||
|
фертильность пыльцы, % |
число растений с данной фертильностью, шт |
средняя фертильность пыльцы в группе, % |
||
|
1989 |
10 |
100-90 |
8 |
99,5 |
|
89-80 |
1 |
83.4 |
||
|
79-70 |
1 |
72.2 |
||
|
1990 |
50 |
100 - 90 |
43 |
98.3 |
|
89-80 |
3 |
81.7 |
||
|
79 - 70 |
2 |
75.6 |
||
|
69-60 |
1 |
61.7 |
||
|
59-50 |
1 |
57.6 |
||
Таблица 4
Показатели семенной продуктивности обоеполых цветков чистеца болотного при свободном опылении в Троицком лесостепном заказнике
|
Год наблюдений |
Выборка* п |
ПП*% |
ПСП, семяпочек* шт |
РСП, эре-МОВ* шт |
ПС, % на плод |
Эремов на плод* шт М ± ш |
Cv, % |
Р*% |
|
1986 |
500 |
92.8 |
2000 |
1050 |
52.5 |
2ЛО±О*ОО5 |
5.06 |
0,24 |
|
1987 |
500 |
79.2 |
2000 |
1390 |
69.5 |
2,78±0.005 |
3.7 |
0.18 |
|
1988 |
500 |
98.6 |
2000 |
1591 |
79.5 |
3.18±0.005 |
3 68 |
0.16 |
|
1989 |
500 |
37.4 |
2000 |
474 |
23.7 |
О.95±О.ОО8 |
18.0 |
0.84 |
|
1990 |
500 |
89.0 |
2000 |
1464 |
73.2 |
2,93±0.005 |
3.66 |
0.17 |
|
2010 |
500 |
93.8 |
2000 |
1551 |
77.6 |
3.10±0.005 |
3.69 |
0.16 |
Примечания; ПП, % - процент цветов, давших плоды. ПС, % - доля семяпочек, развившихся в семена (эремы);
Cv, % - коэффициент варьирования признаков; Р, % - точность определения средней опыта.
Таблица 5
Структура семенной продуктивности обоеполых цветков чистеца болотного при свободном опылении в Троицком лесостепном заказнике
|
Год наблюдений |
Число исследова иных цветков |
Число эремов в одном плоде |
||||
|
0 |
1 |
2 |
3 |
4 |
||
|
Число цветков, давших указанное число эремов. шт. (%) |
||||||
|
1986 |
500 |
36(7,2) |
136 (27.2) |
147 (29.4) |
104 (20.8) |
77(15.4) |
|
1987 |
500 |
104 (20.8) |
23 (4.6) |
32 (6.4) |
61 (12.2) |
280(56.0) |
|
1988 |
500 |
7(1.4) |
15 (3.0) |
78(15.6) |
180 (36.0) |
220 (44,0) |
|
1989 |
500 |
313(62.6) |
41 (8.2) |
50(10.0) |
51 (10.2) |
45(9.0) |
|
1990 |
500 |
55 (11.0) |
19(3.8) |
55(11.0) |
149 (29.8) |
222 (44,4) |
|
2010 |
500 |
31 (6.2) |
28(5.6) |
61 (12.2) |
119(23.8) |
261 (52.2) |
При изучении семенного размножения чистеца болотного в Пермском крае обращает на себя внимание существенная разница в показателях семенной продуктивности обычной пурпуровоцветковой формы и гораздо реже встречающейся белонветко-вой (табл* 6). Особенно отчетливо эти различия проявились в 2012 и 2016 гг. при одновременном исследовании обеих форм, произрастающих в одинаковых условиях (как сорное в посадках картофеля). В оба года наблюдений более высокая семенная продуктивность была свойственна белоцветковой форме (больше, чем в 2 раза). Структура се- менной продуктивности обеих форм (табл. 7) также свидетельствует о преимуществах белоиветко-вой формы; число цветков с 3-4 эремами у этой формы несравненно выше, чем у пурпуровоцветковой, а число цветков, не завязавших плоды, напротив, выше у последней* Зависимость уровня семенной проективности от погодных условий во время цветения и завязывания семян у обеих разноцветковых форм особенно явственно проявилась в 2012 и 2016 гг** резко различных по погодным условиям* Август 2012 г, согласно сводке метеоусловий, оказался с большим числом пасмурных и даже дождливых дней* Напротив, в августе и даже сентябре 2016 г. погода была солнечной и сухой. Показатели семенной продуктивности и ее структура у обеих цветковых форм были несколько выше в 2012 г. по сравнению с засушливым 2016 г*
Таблица 6
Показатели семенной продуктивности разноцветковых форм чистеца болотного при свободном опылении в Пермском крае
|
Год наблюден ий |
Выборка, п |
пп, % |
ПСП* семяпочек, шт |
РСП. эремов, шт |
ПС, % на плод |
Эремов на плод, шт, М ± m |
Cv. % |
р, % |
Невыполненные семена, шт. |
|
Белоцветковая 4 |
юрма |
||||||||
|
2011 |
500 |
78.6 |
2000 |
1155 |
57.8 |
2,31±О.ОО5 |
439 |
0,22 |
142 |
|
Пурпуровоцветковая форма |
|||||||||
|
2012 |
500 |
65.6 |
2000 |
529 |
26.1 |
1.06±0.007 |
15*44 |
0,69 |
31 |
|
Белоцветковая 4 |
юрма |
||||||||
|
2012 |
500 |
86.4 |
2000 |
1235 |
61.8 |
2,47±0.005 |
4,05 |
0,20 |
70 |
|
Белоцветковая (| |
Юрма |
||||||||
|
2013 |
100 |
800 |
400 |
248 |
62.0 |
2.484:0 02 3 |
9.06 |
0.93 |
|
|
Белоцветковая 4 |
юрма |
||||||||
|
2014 |
500 |
69.0 |
2000 |
1014 |
50.7 |
2.03±0.005 |
535 |
0.25 |
|
|
Пурпуровоцветковая форма |
|||||||||
|
2015 |
100 |
260 |
400 |
27 |
6.8 |
0.27±0050 |
1.86 |
0.19 |
|
|
Пурпуровоцветковая форма |
|||||||||
|
2016 |
500 |
52.6 |
2000 |
434 |
21.8 |
О.87Ю.ОО8 |
20.33 |
0.9 |
95 |
|
Белоцветковая (|юрма |
|||||||||
|
2016 |
500 |
79.0 |
2000 |
1158 |
58.0 |
2,32±0.005 |
4,38 |
0,22 |
174 |
Таблица 7
Структура семенной продуктивности разноцветковых форм чистеца болотного при свободном опылении в Пермском крае
|
Год наблюдений |
Число исследо-ванных цветков |
Число эремов Б одном плоде |
||||
|
0 |
1 |
2 |
3 |
4 |
||
|
Число цветков, давших указанное число эремов, шт* (%) |
||||||
|
Белоцветковая форма |
||||||
|
2011 |
500 |
107 (21.4) |
37(7.4) |
89(17.8) |
128(25.6) |
139 (27.8) |
|
Пурпу |
ровоцветковая |
форма |
||||
|
2012 |
500 |
172(34,4) |
171 (34.2) |
121 (24,2) |
28(5,6) |
8(1,6) |
|
Белоцветковая форма |
||||||
|
2012 |
500 |
68(13.6) |
66(13.2) |
92(18.4) |
111 (22.2) |
163(32.6) |
|
Белоцветковая форма |
||||||
|
2013 |
100 |
20 (20.0) |
6(6.0) |
12(12,0) |
30 (30.0) |
32 (32.0) |
|
Белоцветковая форма |
||||||
|
2014 |
500 |
155 (31.0) |
36(7.2) |
75(15.2) |
108(21.6) |
126 (25.2) |
|
Пурпуровоцветковая форма |
||||||
|
2015 |
100 |
74 (74.0) |
25 (25.0) |
1 (1.0) |
0(0) |
0(0) |
|
Пурпуровоцветковая |
форма |
|||||
|
2016 |
500 |
237 (47.4) |
138 (52.5) |
86 (32.7) |
32(12.2) |
7(2.7) |
|
Белоцветковая форма |
||||||
|
2016 |
500 |
105 (21.0) |
46(11.7) |
78 (19.7) |
128(32,4) |
143 (36.2) |
Более эффективное завязывание семян у белоцветковой формы чистеца болотного в сравнении с обычной пурпуровоцветковой, отмеченное в разные по погодным условиям годы, пока не поддает- ся достоверному объяснению. Нужны дополнительные исследования, касающиеся опылителей этого растения* Известно, что пчелиные (наиболее эффективные опылители) хорошо различают как пурпуровые, так и белые окраски [Мазохин-Поршняков, 1965]* причем первые для них наиболее привлекательны. Возможно, что обе формы в пределах одного вида опыляются разными насекомыми, предпочитающими определенные цветки, но различных окрасок [Кау, 1978: цит* по Насимовичу* 1986]* При изоляции растении от насекомых ни одна из форм семена не завязывала.
Так же необходимы и цитоэмбриологические исследования* поскольку внешние экологические факторы для обеих цветковых форм были идентичны* Возможно* что причиной различной семенной продуктивности разноцветковых форм может быть неодинаковая степень нарушений в мегаспорогенезе и развитии зародышевых мешков* как это отмечено у других видов Stachys L. [Старшова* 1992; Чоркина и др * 1996а]. Косвенно об этом свидетельствует и разное количество невыполненных (неполноценных) семян (табл. 6) у обеих цветковых форм* По-видимому* нарушения Б ЦИТОЭМ-брИОЛОГИ ческих процессах у белоцветковой формы наступают на более поздней стадии формирования диаспор. Возможно, что различия в семенной продуктивности у них является проявлением внутривидовой конкуренции*
Подводя ИТОГИ изучения семенной продуктивности чистеца болотного* можно констатировать невысокую семенную продуктивность этого энтомофильного перекрестноопыляемого вида в условиях Урала. В этом отношении он близок к ранее исследованным другим видам Stachys L.
Список литературы Половая структура популяций и семенная продуктивность Stachys palustris L. s. l. на Урале
- Алексеева Л.И., Канев В.А. Динамика содержания фенольных соединений в Stachys palustris L. (Lamiaceae)//Растительные ресурсы. 2014. Т. 50, № 2. С. 242-249
- Вайнагий И.В. Методика статистической обработки материала по семенной продуктивности растений на примере Potentilla aurea L.//Растительные ресурсы. 1973. Т. 9, вып. 2. С. 287-296
- Вайнагий И.В. О методике изучения семенной продуктивности растений//Ботанический журнал. 1974. Т. 59, № 6. С. 826-831
- Годин В.Н., Демьянова Е.И. О распространении гинодиэции у цветковых растений//Ботанический журнал, 2013. Т. 98, № 12. С. 1465-1487
- Демьянова Е.И. Об особенностях распространения гинодиэции в семействе губоцветных//Научные доклады высшей школы. Биол. науки. 1981. № 9. С. 69-73
- Демьянова Е.И. Распространение гинодиэции у цветковых растений//Ботанический журнал. 1985. Т. 70, № 10. С. 1289-1301
- Демьянова Е.И., Пономарев А.Н. Половая структура природных популяций гинодиэцичных и двудомных растений лесостепи Зауралья//Ботанический журнал, 1979. Т. 64, № 7. С. 1017-1024
- Дервиз-Соколова Т.Г. Stachys palustris L. (систематика и география)//Бюллетень Главного ботанического сада. 1975. Т. 80, вып. 3. С. 85-93
- Дикорастущие полезные растения России/под ред. А.Л. Буданцева и Е.Е. Лесиовской. СПб.: Изд-во СПХФА, 2001. 663 с
- Иллюстрированный определитель растений Пермского края/под ред. С.А. Овеснова. Пермь: Кн. мир, 2007. 740 с
- Кнорринг О.Э. Род чистец -Stachys L.//Флора СССР. М.; Л., 1954. Т. 21. С. 200-237
- Костюченко О.И. Химический состав и фармакологические свойства Stachys L.//Растительные ресурсы. 1983. Т. 19, вып. 3. С. 407-413
- Кочиева Е.З. и др. RAPD и ISSR-анализ видов и популяций Stachys//Генетика. 2006. Т. 42, № 7. С. 887-892
- Кордюм Е.Л., Глущенко Г.И. Цитоэмбриологические аспекты проблемы пола покрытосеменных. Киев: Наукова думка, 1976. 197 с
- Куликов П.В. Определитель сосудистых растений Челябинской области. Екатеринбург, 2010. 968 с
- Лакин Г.Ф. Биометрия. М.: Высш. шк., 1973. 291 с
- Лакин Г.Ф. Биометрия. М.: Высш. шк., 1990. 351 с
- Левина Р.Е. К изучению ритма плодоношения некоторых губоцветных (Salvia pratensis L. и Stachys recta L.)//Бюллетень МОИП. Отд. биол. 1970. Т. 75, № 3. С. 53-65
- Левина Р.Е., Старшова Н.П. Семенная продуктивность двух видов чистеца (Stachys recta L. и S. neglecta Klok.) в природных популяциях и в питомнике//Экология цветения и опыления растений. Пермь, 1989. С. 37-46
- Мазохин-Поршняков Г.А. Зрение насекомых. М.: Наука, 1965. 211 с
- Насимович Ю.А. Биологическое значение окраски цветка//Бюллетень МОИП. Отд. биол. 1986. Т. 91, № 5. С. 82-93
- Паушева З.П. Практикум по цитологии растений. М.: Колос, 1974. 288 с
- Плохинский Н.Ф. Математические методы в биологии. М.: Изд-во МГУ, 1978. 263 с
- Пономарев А.Н. Изучение цветения и опыления растений//Полевая геоботаника. М.; Л., 1960. Т. 2. С. 9-19
- Пономарев А.Н., Демьянова Е.И. К изучению гинодиэции у растений//Ботанический журнал. 1975. Т. 60, № 1. С. 3-15
- Работнов Т.А. Методы изучения семенного размножения травянистых растений в сообществах//Полевая геоботаника. М.; Л., 1960. Т. 2. С. 20-40.
- Русских И.В. Эколого-биологические особенности представителей родов Lamium L. и Stachys L. (Lamiaceae) северо-западного Кавказа: автореф. дис. … канд. биол. наук. Краснодар, 2009. 24 с
- Справочник по ботанической микротехнике. Основы и методы/сост. Р.П. Барыкина и др.. М.: Изд-во МГУ, 2004. 311 с
- Старшова Н.П. Эмбриология как метод при изучении причин дефектности диаспор на примере Stachys recta L.//Embryology and seed reproduction: XI International symposium. St-Peterburg: Nauka, 1992. P. 535-536
- Тахтаджян А.Л. Происхождение и расселение цветковых растений. М.; Л.: Наука, 1970. 144 с
- Тахтаджян А.Л. Система магнолиофитов. Л.: Наука, 1987. 438 с
- Чоркина Н.Г., Чиботару А.А., Челак В.Р. Микрои макроспорогенез и оплодотворение у Stachys sieboldii Miq.//Проблемы репродуктивной биологии растений: тез. докл. симпозиума. Пермь, 1996а. С. 223-224
- Чоркина Н.Г., Чиботару А.А., Челак В.Р., Маринеску М.Ф. Цветение, опыление и семенная продуктивность у стахиса//Проблемы репродуктивной биологии растений: тез. докл. симпозиума. Пермь, 1996б. С. 224-226
- Darwin C. The different forms of flowers on plants on the same species. London: John Murray, 1877. 352 p
- Knuth P. Handbuch der Blütenbiologie. Leipzig: Verlag von Wilchelm Engelmann, 1899. Bd.II, T. 2. 705 S
- Mulligan G.A., Munro D.B., McNeil J. The status of Stachys palustris (Labiatae) in North America//Can. J. Bot., 1983. Vol. 61, № 3. P. 679-682
