Половая структура популяций Juniperu s polycarpos C. Koch в Дагестане

Автор: Садыкова Гульнара Алиловна, Амирханова Надежда Алиевна
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 4, 2021 года.
Бесплатный доступ
Изучение половой структуры Juniperus polycarpos C. Koch проводили в популяциях Предгорного (Талгинская, Дубкинско-Миатлинская и Губденская) и Высокогорного (Тляратинская и Цумадинская) Дагестана. В общей сложности исследовано 1 009 особей в пределах всего ареала вида на территории Дагестана, у которых измерены биометрические показатели. В популяциях выявлены многобрачно-двудомные особи J. polycarpos, установлено количественное соотношение женских, мужских и многобрачно-двудомных особей. Определены биометрические показатели особей разных полов, их индексные показатели. Соотношение женских и мужских особей в дагестанских популяциях J. polycapos, как и для других видов Juniperus в целом, может быть связано с экологическими условиями среды (увеличение высоты над уровнем моря места произрастания и более засушливые условия). Вместе с тем изменение соотношения высоты и диаметра кроны у деревьев разных половых групп может зависеть от возраста особей в популяции, сроков вступления их в генеративное состояние и другими процессами.
Juniperus polycarpos c. koch, популяции, двудомные виды, многобрачно- двудомные растения, соотношение полов, полидийокисты, биоморфология
Короткий адрес: https://sciup.org/147236784
IDR: 147236784 | УДК: 581.8 | DOI: 10.17072/1994-9952-2021-4-249-255
Sexual structure of Juniperus polycarpos C. Koch populations in Dagestan
The study of the sexual structure of Juniperus polycarpos C. Koch was carried out in the populations of Piedmont (Talginskaya, Dubkinsko-Miatlinskaya, and Gubdenskaya) and highmountain (Tlyaratinskaya and Tsumadinskaya) Dagestan. In total, 1009 individuals were studied within the entire range of the species in the territory of Dagestan, in which biometric parameters were measured. In populations, polygamous-dioecious individuals of J. polycarpos were identified, and a quantitative ratio of female, male, and polygamous-dioecious individuals was established. Biometric indicators of individuals of different sexes and their index indicators have been determined. The ratio of females and males in the Dagestan populations of J. polycapos, as well as for other Juniperus species in general, may be related to the ecological conditions of the environment (an increase in the altitude above sea level of the place of growth and more arid conditions). At the same time, the change in the ratio of the height and diameter of the crown in trees of different sex groups may depend on the age of individuals in the population, the timing of their entry into the generative state, and other processes.
Текст научной статьи Половая структура популяций Juniperu s polycarpos C. Koch в Дагестане
При решении задач охраны редких видов растений популяционные исследования становятся незаменимыми. Знание биологии видов и структуры их популяций является основой, с помощью которой можно прогнозировать как дальнейшее развитие популяции, так и реакцию растений на неблагоприятные воздействия среды обитания [Пушкарь, Майоров, 2003].
Одним из основных показателей структуры популяций видов является половая структура, отражающая определенное соотношение особей разного пола в популяции. Половая структура популяции напрямую связана с её воспроизводством и устойчивостью и имеет практическую значимость для сохранения биоразнообразия.
При изучении двудомных растений исследование лабильности половой структуры необходимо для оценки динамических процессов в популяциях. Соотношение полов связывают с условиями внешней среды, с историей развития видов и т.д. В более стабильных условиях для некоторых видов отмечается видоспецифичность половой структуры, хотя у других видов соотношение полов колеблется от почти полного преобладания одного пола до равных количеств мужских и женских особей [Демьянова, Пономарев, 1979].
Для видов можжевельника изменение соотношения полов связывают с разнообразием факторов: в Пиренеях причиной преобладания мужских особей J. thurifera считают вторичное заселение ксерофиль-ных пастбищ, в Альпах отмечается увеличение доли мужских особей J. thurifera на террасированных полях [Borel, Polidori, 1983], уменьшение доли женских особей на больших высотах [Ortiz, Arista, Talavera, 2002], по краю диапазона [Marion, Houle, 1996]. Только женские особи J. communis подвержены влиянию биотических факторов (клещи, насекомые) [Ward, 2007].
У J. excelsa с факторами среды связывают разнообразие и изменчивость половых типов и форм. Отмечают изменчивость моноэцичности и диэцичности особей с высотой над уровнем моря, засушливостью и старением особей [Douaihy et al., 2013]. Для данного вида моноэцичность оценивалась как средство обеспечения самоопыления ценой увеличения инбридинга. Тогда как двудомность, характерная для Juniperus polycarpos C. Koch [Fisher, Gardner, 1995], нередко рассматриваемого в качестве подвида J. excelsa , приписывалась генетически пластичным видам [Ortiz, Arista, Talavera, 2002].
Таким образом, половая структура популяции лишь в самом обобщенном виде может быть представлена средним численным соотношением мужских и женских особей. Реально она характеризуется соотношением полов в различных возрастных группах и отражает не только интенсивность размножения, но и общий потенциал популяции: ход численности, продукцию биомассы, уровень популяционных энергозатрат и др. [Шилов, 2003].
Цель настоящего исследования – изучение половой структуры в различных популяциях Juniperus pol-ycarpos в Дагестане, выявление соотношения разнополых особей в различных условиях среды и зависимости от возрастных состояний особей для оценки устойчивости популяции.
Материал и методика
Изучение половой структуры можжевельника многоплодного (Juniperus polycarpos C. Koch) проводили в популяциях Предгорного (Талгинская, Дубкинско-Миатлинская и Губденская) и Высокогорного (Тляратинская и Цумадинская популяции) Дагестана.
Дубкинско-Миатлинская популяция расположена на западном склоне хр. Надырбег, прилегающего к р. Сулак. Участок находится между Чиркейским и Миатлинским водохранилищами, близ селений Дубки и Миатли. J. polycarpos произрастает на высотах от 150 м (у Миатлинского вдхр.) до 600 м (у Чиркейско-го вдхр.). Талгинская популяция (Талгинское ущелье или ущелье Истису-кака) расположена на юго-западе в 20 км от г. Махачкалы у основания г. Кукуртбаш. Здесь можжевеловые редколесья расположены на высотах от 400 до 600 м. Губденская популяция расположена в Центрально-предгорном Дагестане в 5 км от с. Губден. J. polycarpos произрастает на южных отрогах хр. Чонкатау и северных отрогах хр. Шам-халдаг с высоты от 700 до 1 000 м.
Тляратинская популяция (Чадоколобо-Анцухский участок) расположена на южных, юго-западных, западных, юго-восточных и восточных склонах отрогов Богосского хр. вдоль р. Аварское Койсу, близ с. Чадоколоб и Анцух Тляратинского р-на. Можжевеловые редколесья здесь произрастают на высотах от 1 342 до 1 589 м над ур. моря; Цумадинская популяция (Гадаичи-Эчединский участок) расположена вдоль р. Андийское Койсу, на отрогах Кад Богосского хр., близ с. Гадаичи и на склонах г. Омар Вахуна-убетер Снегового хр. близ с. Эчеда Цумадинского р-на на высотах 1 168–1 257 м (рисунок).
В общей сложности нами было исследовано 1 009 особей в пределах всего ареала вида на территории Дагестана. У каждой особи измерены высота и диаметр ствола деревьев.
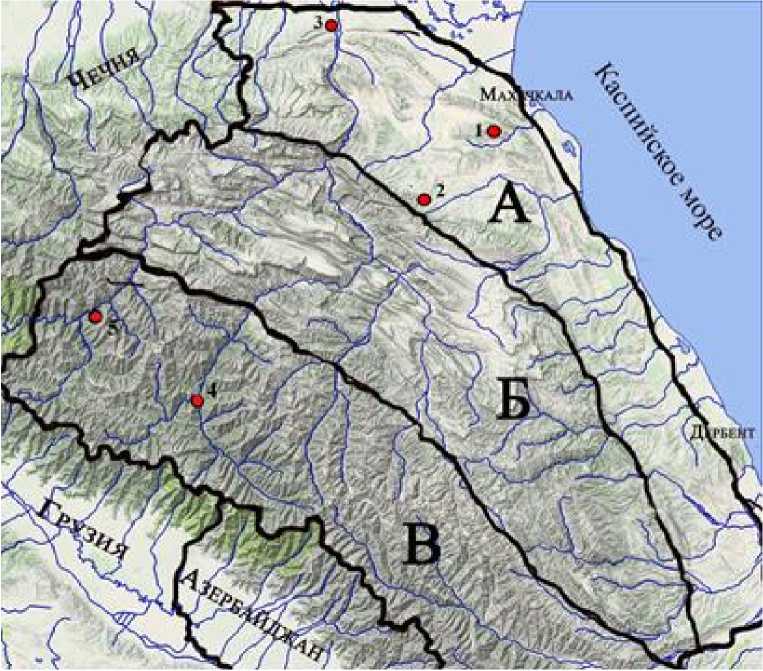
Районы распространения Juniperus polycarpos в Дагестане:
-
1 – Талгинское ущелье, 2 – Губденский участок, 3 – Дубкинско-миатлинский участок, 4 – Чадоколобо-Анцухский участок, 5 – Гадаичи-Эчединский участок
[Distribution areas of Juniperus polycarpos in Dagestan:
1 – Talginsky gorge, 2 – Gubdensky section, 3 – Dubkinsko-miatlinsky section, 4 – Chadoko-lobo-Antsukhsky section, 5 – Gadaichi-Echedinsky section]
Результаты исследований
Juniperus polycarpos считается двудомным растением, однако в дагестанских популяциях нами выявлены особи с микро- и макростробилами на одном дереве. Такие особи по классификации Е.И. Демьяновой [Эмбриология…, 2000] отнесены к многобрачно-двудомным растениям или полидиойкистам.
Из более 1 000 проанализированных особей J. polycarpos 25.2% составили женские, 22.4 % – мужские, 1.5% – полидиойкисты. Более половины (50.3%) особей были без половых признаков. Наибольшее количество женских особей выявлено в губденской (39.3%) и тляратинской (33.3%) популяциях, где их количество превышает мужские в 1.4 и 1.2 раза соответственно. В целом количество женских особей в популяциях больше в 1.1–1.4 раза, в среднем в 1.1 раза. Исключение составляет талгинская популяция, где количество мужских особей больше женских в 1.5 раза (табл. 1).
Таблица 1
Количественный состава особей J. polycarpos разного пола в популяциях Дагестана [Quantitative composition of J. polycarpos individuals of different sexes in Dagestan populations]
|
Популяция |
Половое соотношение особей |
|||
|
женские |
мужские |
полидиойкисты |
неопределенного пола |
|
|
Талгинская |
49/ 15.0 |
74/ 22.7 |
5 /1.5 |
198/ 60.7 |
|
Губденская |
46/ 39.3 |
32/ 27.4 |
- |
39/ 33.3 |
|
Дубкинско-миатлинская |
76/ 25.2 |
55/ 18.2 |
12/ 4.0 |
159/ 52.6 |
|
Тляратинская |
36/ 33.3 |
31/ 28.7 |
- |
41/38.0 |
|
Цумадинская |
47/ 30.1 |
34/ 21.8 |
4/ 2.6 |
71/ 45.5 |
|
Общее |
254/ 25.2 |
226/ 22.4 |
21/ 2.1 |
508/ 50.3 |
Примечание. Абсолютный / относительный показатели.
Средние значения высоты женских особей в различных популяциях варьируют от 2.8 м (дубкинско-миатлинская) до 4.0 м (цумадинская), у мужских – от 2.7 м (дубкинско-миатлинская) до 3.8 м (тляратин-ская). Средние значения диаметра крон женских особей варьирует от 2.6 (дубкинско-миатлинская) до 3.3 м (губденская), у мужских – от 2.6 м (дубкинско-миатлинская) до 3.2 м (цумадинская).
В целом, во всех популяциях женские особи выше и шире мужских (табл. 2). Наибольшие биометрические показатели отмечены у многобрачно-двудомных особей цумадинской популяции (5.2 м и 5.6 м высота и диаметр кроны, соответственно).
Многобрачно-двудомные особи выше женских в 1.3 раза, мужских – в 1.4 раза в цумадинской популяции, в дубкинской популяции – в 1.1 раза. В талгинской популяции, напротив, полидийкисты в 1.1 раза ниже женских особей.
Индексные значения для женских и мужских особей в целом одинаковы и составляют 1.1–1.4, для многобрачно-двудомных особей индексные показатели деревьев несколько ниже: от 0.9 (цумадинская популяция) до 1.1 (дубкинско-миатлинская популяция).
Таблица 2
Биометрические показатели особей J. polycarpos разного пола в популяциях Дагестана [Biometric indicators of J. polycarpos individuals of different sexes in Dagestan populations]
|
Популяция |
Пол |
Высота особи, см |
Диаметр кроны, см |
Индекс особи |
||
|
x ± sx |
CV, % |
x ± s x |
CV, % |
|||
|
Талгинская |
женские |
3.1±0.23 |
51.3 |
3.0±0.25 |
58.2 |
1.1 |
|
мужские |
2.8±0.16 |
47.4 |
2.7±0.21 |
63.4 |
1.2 |
|
|
полидиойкисты |
2.9±0.47 |
35.9 |
3.2±0.43 |
29.5 |
1.0 |
|
|
Губденская |
женские |
3.4±0.22 |
42.9 |
3.3±0.26 |
52.3 |
1.2 |
|
мужские |
3.5±0.28 |
45.1 |
2.8±0.28 |
52.9 |
1.4 |
|
|
Дубкинско-миатлинская |
женские |
2.8±0.13 |
39.8 |
2.6±0.12 |
40.3 |
1.1 |
|
мужские |
2.7±0.14 |
37.3 |
2.6±0.12 |
34.6 |
1.1 |
|
|
полидиойкисты |
3.0±0.24 |
27.1 |
3.0±0.24 |
27.9 |
1.1 |
|
|
Тляратинская |
женские |
3.9±0.21 |
32.4 |
3.2±0.18 |
34.0 |
1.1 |
|
мужские |
3.8±0.25 |
36.3 |
3.1±0.23 |
41.6 |
1.3 |
|
|
Цумадинская |
женские |
4.0±0.16 |
27.8 |
3.1±0.17 |
36.7 |
1.4 |
|
мужские |
3.7±0.25 |
38.7 |
3.2±0.23 |
42.1 |
1.2 |
|
|
полидиойкисты |
5.2±0.23 |
8.8 |
5.6±0.38 |
13.3 |
0.9 |
|
Наименьшие значения биометрических показателей по популяциям у особей дубкинско-миатлинской популяции, наибольшие – у женских особей в цумадинской популяции, мужских – в тляратинской.
Обсуждение результатов
В исследуемой популяции Juniperus polycarpos , расположенной на северном пределе своего распространения, доля многобрачно-двудомных особей столь же незначительна, как и у других популяций этого вида, расположенных в центре своего ареала. Так, например, в Омане и Пакистане (провинция Белуджистан) в популяциях Juniperus excelsa subsp . polycarpos многобрачно-двудомные особи также встречаются крайне редко. Они составляют 2.3% от общего количества особей [Moinuddin, Shahid, Hafeez, 1990]. Соотношение же мужских и женских особей Juniperus excelsa близко к 1.
Для вида в целом считается оптимальным равное соотношение женских и мужских особей. Изменение соотношения полов связывают с экологическими условиями и генетическими факторами, которые могут обусловливать пол дерева. Преобладание женских особей связывают с внутрипопуляционным механизмом сохранения вида; с адаптацией к конкретным условиям среды обитания [Ainsworth, 1999]. В засушливых условиях отмечают уменьшение числа женских особей [Демьянова, 2013; Douaihy et all, 2013] и увеличение доли мужских деревьев, то же наблюдается и с увеличением высоты места произрастания популяции (например, у Juniperus communis subsp. alpina ) [Ortiz, Arista, Talavera, 2002].
В нашем исследовании только для талгинской популяции выявлено преобладание мужских особей. Сравнительный анализ условий этого ущелья в связи с геоморфологией указывает на более засушливые здесь условия по сравнению с условиями произрастания других дагестанских популяций. Климат резко континентальный, характерны резкие перепады летних и зимних температур. Средняя температура в летний период здесь составляет 20–21ºС, а в зимний – 2.5ºС. Это ущелье изолировано от морских воздушных потоков горными массивами Тарки-тау и Нарат-тюбе, что способствует сдерживанию облаков и, соответственно, сухости климата.
Условия произрастания двух других ближайших популяций (губденская и дубкинско-миатлинская) несколько мягче. Оба ущелья направлены к морю и не изолированы крупными хребтами и массивами, в связи с чем более подвержены проникновению влажных северно-восточных каспийских воздушных масс.
Таким образом, можно говорить о том, что половая структура талгинской популяции с большей долей в составе мужских особей связана с возможным влиянием более засушливых экологических условий.
Кроме того, ряд авторов связывают разницу в соотношении полов у можжевельника с возрастным составом особей в популяции [Falinski 1980; Ward, 1982; Gauquelin et all, 2002].
Так, средний возраст особей во всех предгорных популяциях почти одинаковый: в талгинской популяции составляет 135 лет, в дубкинско-миатлинской – 168 лет, в губденской – 152 года. Однако в талгин-ской популяции количество особей предгенеративного возраста в 1.5 раза превышает количество особей генеративного и постгенеративного состояний.
В то же время биоморфологическая конусовидная конструкция кроны как женских, так и мужских растений J. polycarpos во всех популяциях, являясь результатом их функциональной адаптации, находящейся в состоянии подвижного равновесия с условиями произрастания вида и с онтогенетическим развитием особей, характерна для молодых женских и мужских особей (I>1) в связи с активным их ростом в благоприятных условиях среды. Индексные значения кроны многобрачно-двудомных видов ниже 1. Крона туповершинная ширококонусная.
В целом, преобладание размеров многобрачно-двудомных растений над размерами раздельнополых особей J. polycarpos и преобладание размеров женских над мужскими особями, выявленное для наших популяций, подтверждается и на примере других популяций.
В популяциях Juniperus excelsa в Пакистане деревья имели различные значения высоты в разных районах, но во всех случаях минимальные показатели имели мужские особи [Fisher, Gardner, 1995], при том, что годовой прирост у мужских особей может быть и немного больше [Ortiz, Arista, Talavera, 2002; Асадулаев, Садыкова, 2011].
Возраст особей и пол деревьев влияют на ежегодный вегетативный прирост. Средний годовой прирост у мужских особей больше. Эта продолжающаяся разница в росте приводит к большему кумулятивному росту мужских особей и, следовательно, изменению биометрических показателей деревьев разного пола с возрастом [Григорян, 1977; Ward, 2007].
Таким образом, биометрические показатели и биоморфологическая структура особей разного пола отражают возраст особей в популяциях и возраст популяции в целом. Наименьшие биометрические параметры особей дубкинско-миатлинской популяции указывают на особенности условий произрастания популяции, а высокая пластичность, характерная для видов рода Juniperus в целом [Иванова, 1946; Dou-aihy et al., 2013], приводит к изменению соотношения полов в различных условиях среды.
Заключение
Соотношение женских и мужских особей в дагестанских популяциях J. polycapos , как и для других видов Juniperus в целом, может быть связано с экологическими условиями среды. В нашем случае это увеличение высоты над уровнем моря места произрастания и более засушливые условия, например, на южных склонах гор. Вместе с тем изменение соотношения высоты и диаметра кроны у деревьев разных половых групп может зависеть от возраста особей в популяции, сроков вступления их в генеративное состояние и другими процессами.
Список литературы Половая структура популяций Juniperu s polycarpos C. Koch в Дагестане
- Асадулаев З.М., Садыкова Г.А. Структурная и ресурсная оценка природных популяций можжевельника продолговатого (Juniperus oblonga Bieb.) в Дагестане. Махачкала: Наука ДНЦ, 2011. 216 с.
- Григорян А.А. Некоторые итоги интродукции можжевельников Кавказа в Ереванском ботаническом саду // Бюллетень ботанического сада Академии наук Армянской ССР. 1977. № 24. С. 5-48.
- Демьянова Е.И. Половая структура природных популяций некоторых двудомных и гинодиэцичных растений окского заповедника // Вестник Пермского университета. Сер. Биология. 2013. Вып. 2. С. 1119.
- Демьянова Е.И., Пономарев А.Н. Половая структура природных популяций гинодиэцичных и двудомных растений лесостепи Зауралья // Ботанический журнал. 1979. Т. 64, № 7. С. 1017-1024.
- Иванова А.В. Можжевеловые редколесья Южной Армении // Труды Ботанического ин-та АН Армянской ССР. 1946. Т. 4. С. 109-155.
- Пушкарь В.С., Майоров И.С. Экология. Владивосток, 2003. 188 с.
- Шилов И.А. Экология. М.: Высш. шк., 2003. 512 с.
- Эмбриология цветковых растений. Терминология и концепции / под ред. Т.Б. Батыгиной. СПб.: Мир и семья, 2000. Т. 3. Системы репродукции. 640 с.
- Ainsworth C.C. Sex determination in plants. Oxford: Bios. Scientific Publishers, 1999.
- Borel A., Polidori J.-L. Le genévrier thurifère (Juniperus thurifera L.) dans le Parc National du Mercan-tour (Alpes-Maritimes) // Bulletin de la Société Botanique de France. Lettres Botaniques. 1983. Vol. 130, № 3. P. 227-242. DOI: 10.1080/01811797.1983.10824590
- Douaihy Ch.B. et al. Ecological characterization of the Juniperus excelsa stands in Lebanon // Ecologia mediterranea. 2013. Vol. 39, № 1.
- Falinski J.B. Vegetation dynamics and sex structure of the population of pioneer dioecious woody plants // Vegetation. 1980. Vol. 43. P. 23-38.
- Fisher M., Gardner A.S. The status and ecology of a Juniperus excelsa subsp. polycarpos. Woodland in the northern mountains of Oman // Vegetation. 1995. Vol. 119. P. 33-51.
- Gauquelin Th. et al. Sex ratio and sexual dimorphism in mountain dioecious thuriferous juniper (Juniperus thurifera L., Cupressaceae) // Botanical Journal of the Linnean Society. 2002. Vol. 138. P. 237-244.
- Marion C., Houle G. No differential consequences of reproduction according to sex in Juniperus communis var. depressa (Cupressaceae) // American Journal of Botany. 1996. Vol. 83. P. 480-488.
- Moinuddin A., Shahid Sh.S., Hafeez B.A. Population structure and dynamics of Juniperus excelsa in Balouchistan // Pakistan Journal of Vegetation Science. 1990. № 1. P. 271-276.
- Ortiz P.L., Arista M., Talavera M. Sex ratio and reproductive effort in the Dioecious Jiniperus communis subsp. alpine (Suter) Celak. (Cupressaceae) along an altitudinal gradient // Annals of botany. 2002. Vol. 89. P. 205-211.
- Ward L.K. Lifetime sexual dimorphism in Juniperus communis var. communis // Plant Species Biology. 2007. Vol. 22. P. 11-21. Doi: 10.1111/j.1442-1984.2007.00171.x
- Ward L.K. The conservation of juniper: longevity and old age // Journal of Applied Ecology. 1982. Vol. 19. P. 917-928.
