Половой диморфизм размеров тела у одного из видов жужелиц (Notiophilus reitteri Spaeth)
")
Автор: Будилов П.В.
Журнал: Региональные проблемы @regionalnye-problemy
Рубрика: Биология. Экология. Геоэкология
Статья в выпуске: 1 т.16, 2013 года.
Бесплатный доступ
В условиях южного макросклона хребта Эзоп (Хабаровский край) подтверждена биотопическая приуроченность Notiophilus reitteri Spaeth к бореальным лесам. Выявлен половой диморфизм по абсолютным размерам тела: самки несколько крупнее самцов.
Половой диморфизм, размеры тела
Короткий адрес: https://sciup.org/14328823
IDR: 14328823 | УДК: 595.762.12
Sexual dimorphism in the Notiophilus reitteri Spaeth body size
At the southern macro-slope of Aesop (Khabarovsk Territory) it is investigated and confirmed the Notiophilus reitteri Spaeth biotope attachment to boreal forests. It is revealed the species sexual dimorphism in the absolute body size: females are somewhat larger than males.
Текст научной статьи Половой диморфизм размеров тела у одного из видов жужелиц (Notiophilus reitteri Spaeth)
Половой диморфизм у жужелиц чаще всего проявляется в признаках, непосредственно связанных с процессом полового размножения (строение передних лапок, длина усов и т.д.). Кроме того, как правило, самки жужелиц несколько крупнее и массивнее самцов; в длину они обычно превосходят самцов в среднем на 8-12 %, а по весу - в 1,3-1,6 и даже в 2 раза [5]. Хотя данное явление хорошо известно для жужелиц в целом, половой диморфизм размеров тела у Notiophilus reitteri рассматривается впервые.
Notiophilus reitteri Spaeth 1900 - мелкие нелетающие жужелицы размером 4,5-6 мм. Наибольшую активность проявляют в дневное время. Питаются преимущественно ногохвостками [6]. N. reitteri - транспалеарктический вид с ареалом, простирающимся от Скандинавии по северо-западу Европейской части, по северному Уралу и далее до Монголии и юга Приморского края [И]. На северо-западе своего ареала N. reitteri чаще всего встречается в увлажненных еловых лесах [12], крупнотравных и зеленомошных ельниках [4]. В Среднем Приобье - многочисленнее в приречных елово-кедровых зеленомошных лесах и отсутствует в лесах на водоразделах [3]. В Баргузинском заповеднике, помимо средней части лесного пояса и верхней части горно-лесного кедрово-стланикового пояса, был отмечен на высокогорном альпийском лугу [9].
На территории Приморского края N. reitteri обитает в поясе темнохвойной тайги. Чаще встречается в еловых или пихтовых лесах на мхах, на открытых участках [8]. В высокогорьях Южного Сихотэ-Алиня вид отмечался в субальпийском поясе кустарников, отдельные находки были сделаны в альпийском поясе, но в целом он не характерен для высокогорного комплекса и, вероятно, в альпийскую и субальпийскую зоны проникает случайно [7].
В Хабаровском крае (Верхнебуреинский р-н) N. reitteri - один из доминантных видов жужелиц темнохвойной тайги [1, 10].
Район исследований
Наши исследования проводились в северной части Верхнебуреинского района Хабаровского края. Для дан ной территории выделяется ряд типов растительного покрова, формирующих три хорошо выраженных пояса: лесной (горно-таежный), подгольцовый и гольцовый.
Горно-таежный пояс сформирован в основном светлохвойными лиственничными лесами, образованными лиственницей Гмелина, а также темнохвойными еловыми лесами из ели аянской, произрастающими на горных склонах и террасах, отдельными островками в распадках, неширокими полосами вдоль ручьев и рек до подгольцовых седловин. Верхние границы лесного пояса колеблются от 900 до 1500 м над уровнем моря.
На высотах 800-1600 м над у. м. расположен пояс кедрового стланика, среди которого встречается ольховник кустарниковый и рододендрон золотистый. Напочвенный покров состоит из зеленых мхов и лишайников.
Выше, на пологих склонах и каменистых плато, расположена лишайниковая (ягельная) тундра, где в углублениях встречаются рябинник крупноцветковый и рододендрон мелколистный [2].
Материалы и методы
Сбор материала осуществлялся на высотах от 850 до 1800 метров над у. м. в наиболее типичных биотопах южного макросклона хребта Эзоп и его отрогов, выступающих в роли водоразделов бассейнов рек Правая Бурея, Олга и Олгакан (рис. 1). Материал был собран преимущественно автором с помощью почвенных ловушек Барбера в вегетационные периоды 2006-2012 гг. Использовался также ручной сбор и отбор почвенных проб. Часть материала с отрогов хребта Эзоп любезно предоставил Л.А. Триликаускас.
Краткие характеристики участков сбора материала: 1 - ельнико-лиственничник, граничащий с поясом кедрового стланика, верховья р. Олгакан, выс. 1400 м над у. м.; 2 - ельник зеленомошный с примесью лиственницы и кедрового стланика, в верховьях реки Самыр, выс. 1200 м над у. м.; 3 - долинный лиственничник с примесью ели, в устье р. Бурейка, выс. 1000 м над у. м.; 4 -лиственничник брусничный, близ слияния рек Олгакан и Малый Олгакан, выс. 1000 м над у. м.; 5 - долинный лиственничник осоково-разнотравный, на правом берегу реки Олгакан, выс. 900 м над у. м.; 6 - лиственничник
|
i|m |
я |
w щ |
1# |
||||
|
У|№^ |
1ВЙ |
■ |
|||||
|
ill |
tw |
■ |
|||||
|
Х-ш^ш © |
Вб» ftSXWX |
я WI |
■ |
1» ■1 |
|||
|
III |
|||||||
|
в в |
■S |
й|| Ч У/ Sil |
III |
8 |
|||
|
8# |
|||||||
|
MBS |
8 |
■ |
^6 |
||||
|
|||||||
Рис. 1. Расположение участков сбора материала зеленомошно-разнотравный, вдоль ручья Агды, на окраине поселка Софийск, выс. 900 м над у. м.; 7 - долинный лиственничник бруснично-разнотравный, в районе слияния рек Олта и Самыр, выс. 1000 м над у м.; 8 -долинный лиственничник бруснично-разнотравный, на правом берегу реки Правая Бурея, выс. 850 м над у. м.; 9 - ельнико-лиственничник, в верховьях ключа - притока р. Правая Бурея, выс. 1400 м над у. м.; 10 - каменистая лишайниковая тундра на водоразделе рек Правая Бурея и Самыр, выс. 1500 м над у. м.; 11 - лишайниковая тундра, на водоразделе рек Правая Бурея и Самыр, выс. 1600 м над у. м.; 12 - лишайниковая тундра с отдельными кустами кедрового стланика, на водоразделе рек Правая Бурея и Олга, выс. 1700 м над у. м.; 13 - лишайниковая тундра, на водоразделе рек Правая Бурея и Селемджа, выс. 1800 м над у. м.
Все измерения выполнялись при помощи бинокулярного микроскопа МБ С-9 со встроенным микрометром при увеличении х1 б с точностью до 0,05 мм.
Дополнительно вычислялись: L - суммарная длина тела (HL+PLm+EL) и пропорции тела: головы (HW/HL и HL/HW); передне спинки (PA/PLm, PW/PLm, PB/PLm и PLm/PA, PLm/PW, PLm/PB); надкрылий (EL/EWh EW/EL).
Для выборок объемом более 20 экземпляров был проведен анализ и статистическая обработка морфометрических признаков (рис. 2).
Статистический анализ проводился средствами программы STATISTIC А 6.0. При сравнении выборок объемом более 30 экз. использовался тест Стьюдента для независимых групп, в остальных случаях - U критерий Манна-Уитни. Во всех случаях пороговым значением достоверности различий было принято р<0,05.
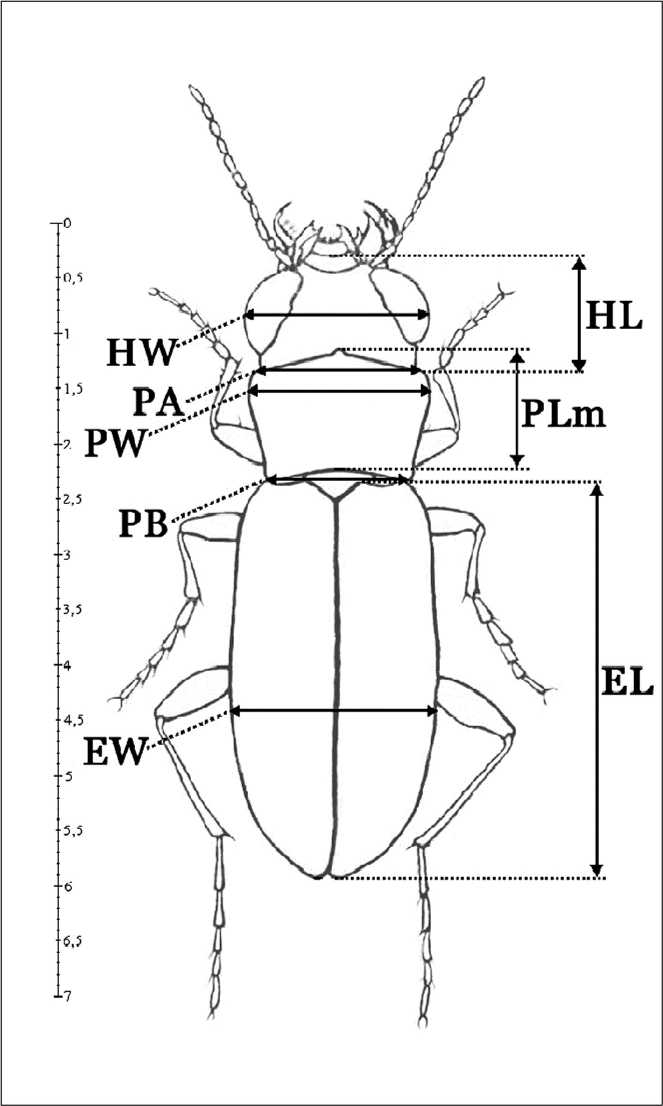
Рис 2. Схема промеров:
HW - ширина головы, включая глаза; HL - длина головы от переднего края наличника до заднего края виска; РА - ширина переднего края переднеспинки; PW - максимальная ширина переднеспинки; РВ - ширина основания переднеспинки; PLm -длина переднеспинки по средней линии; EW - максимальная ширина надкрылий; EL - длина надкрылий от плечевого зубчика до вершины
Результаты и обсуждение
В районе исследований имаго N. reitteri отмечались с середины июня до начала сентября. Вид отмечен во всех обследованных биотопах лесного пояса, но наиболее часто встречается в увлажненных моховых прирусловых ельниках и брусничных лиственничниках, а также в нижней границе кедрово-стланикового пояса. На открытых участках с тундровой растительностью (участки 10,11,12 и 13), расположенных на высотах более 1600 м над у. м., N. reitteri не обнаружен, что подтверждает его биотопическую приуроченность к бореальным лесам. Всего собрано 21 б экз. имаго жуков N. reitteri (табл. 1).
В выборке N. reitteri с участка 3, где было собрано наибольшее количество экземпляров, было отмечено проявление полового диморфизма в размерах тела- ^ несколько превосходили ^ по всем промерам (табл. 2). Применение теста Стьюдента подтвердило статистическую значимость различий между (^ и S^ по всем параметрам, кроме длины головы (HL) и длины надкрылий (EL) (табл. 4). При этом пропорции тела ^ и ^ практически не отличались (табл. 3). Статистически значимые различия были отмечены только в пропорциях надкрылий (EW/EL и EL/EW) (табл. 5).
Аналогичные результаты получены и при сравнении <5' и Ьрс участка 2. Применение U критерия Манна-Уитни подтвердило половой диморфизм по тем же признакам, что и на участке 3.
В наибольшей степени проявление полового диморфизма отмечено на участке 1 - здесь ^ и Ц^ статистически значимо различались по всем абсолютным размерам. Сходство в пропорциях тела (^ и $ также оставалось на достаточно высоком уровне, но в отличие от выборок с участков 2 и 3, ^ и $ участка 1 статистически значимо различались по пропорциям головы (HW/ HL и HL/HW) и не различались по пропорциям надкрылий.
Самцы N. reitteri, собранные на участке 3, по большинству параметров крупнее самцов с участков 1 и 2: они имели более крупную голову (HL и HW), более широкий передний край переднеспинки (РА), более длинные надкрылья (EL) и большую общую длину тела (L). Кроме того, максимальная ширина переднеспинки (PW) ^ с участка 3 больше, чем у ^ с участка 2.
При сравнении выборок ^ с участков 1 и 2 каких-либо статистически значимых различий по средним размерам тела отмечено не было.
В пропорциях тела самцов с различных участков значительных расхождений не обнаружено. Статистически значимым только различие в пропорциях переднеспинки (PW/PLm и PLm/PW) между ^ участков 2 и 3; различие по относительной ширине переднеспинки (PW/PLm) между ^ участков 2 и 1 характеризовалось значением уровня достоверности очень близкого к пороговому (р=0,0491).
Таблица 1
Количество экземпляров N. reitteri по участкам сбора
|
Номера участков |
Всего экз. |
|||||||||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
и |
12 |
13 |
||
|
Самцы |
24 |
21 |
48 |
3 |
— |
1 |
4 |
2 |
2 |
— |
— |
— |
— |
105 |
|
Самки |
25 |
25 |
52 |
1 |
1 |
1 |
3 |
2 |
1 |
— |
— |
— |
— |
111 |
|
Сумма |
49 |
46 |
100 |
4 |
1 |
2 |
7 |
4 |
3 |
— |
— |
— |
— |
216 |
Таблица 2
Средние значения измеренных параметров и ^ в мм (цифрой обозначен номер участка)
|
Параметр |
<5 1 |
<5 2 |
?з |
<5 3 |
||
|
HL |
1,06 |
1,04 |
1,06 |
1,03 |
1,07 |
1,06 |
|
HW |
1,64 |
1,58 |
1,62 |
1,57 |
1,65 |
1,63 |
|
PLm |
1,10 |
1,06 |
1,11 |
1,06 |
1,11 |
1,07 |
|
PW |
1,55 |
1,51 |
1,55 |
1,49 |
1,57 |
1,53 |
|
РА |
1,41 |
1,36 |
1,41 |
1,36 |
1,44 |
1,40 |
|
РВ |
1,23 |
1,18 |
1,24 |
1,20 |
1,24 |
1,20 |
|
EL |
3,48 |
3,41 |
3,47 |
3,42 |
3,51 |
3,49 |
|
EW |
2,01 |
1,92 |
1,98 |
1,90 |
2,00 |
1,93 |
|
L |
5,64 |
5,50 |
5,63 |
5,51 |
5,69 |
5,62 |
Таблица 3
Средние значения попорций £) и ^ (цифрой обозначен номер участка)
|
Параметр |
?! |
<5'1 |
? 2 |
<5'2 |
? 3 |
<5'3 |
|
HW/HL |
1,55 |
1,52 |
1,53 |
1,52 |
1,55 |
1,54 |
|
HL/HW |
0,65 |
0,66 |
0,65 |
0,66 |
0,65 |
0,65 |
|
PA/PLm |
1,29 |
1,29 |
1,28 |
1,28 |
1,29 |
1,30 |
|
PW/PLm |
1,41 |
1,43 |
1,40 |
1,40 |
1,41 |
1,43 |
|
PB/PLm |
1,12 |
1,12 |
1,13 |
1,13 |
1,12 |
1,12 |
|
PLm/PA |
0,78 |
0,78 |
0,78 |
0,78 |
0,77 |
0,77 |
|
PLm/PW |
0,71 |
0,70 |
0,72 |
0,71 |
0,71 |
0,70 |
|
PLm/PB |
0,89 |
0,90 |
0,89 |
0,89 |
0,90 |
0,89 |
|
EW/EL |
0,58 |
0,56 |
0,57 |
0,56 |
0,57 |
0,55 |
|
EL/EW |
1,74 |
1,78 |
1,75 |
1,80 |
1,76 |
1,81 |
Таблица 4
Уровень достоверности различий р между выборками различных участков
|
Параметр |
Сравниваемые выборки (цифра - номер участка) |
||||||||
|
d vs V 1 |
<5 vs V 2 |
d vs V 3* |
<5 1 vs <5'2 |
<5 2 vs <5'3 |
<5 3 vs <54 |
^1 vs $2 |
^2 vs ?з |
?3 vs |
|
|
HL |
0,0332 |
0,0550 |
0,2562 |
0,9275 |
0,0249 |
0,0192 |
0,6695 |
0,2889 |
0,5978 |
|
HW |
0,0001 |
0,0009 |
0,0368 |
0,6008 |
0,0001 |
0,0001 |
0,1160 |
0,0125 |
0,4496 |
|
PLm |
0,0039 |
0,0252 |
0,0011 |
0,7936 |
0,4494 |
0,2770 |
0,7785 |
0,5865 |
0,7400 |
|
PW |
0,0091 |
0,0007 |
0,0012 |
0,1795 |
0,0018 |
0,1278 |
0,5475 |
0,1237 |
0,4084 |
|
PA |
0,0009 |
0,0004 |
0,0001 |
0,7244 |
0,0020 |
0,0052 |
0,7415 |
0,0847 |
0,0344 |
|
PB |
0,0019 |
0,0096 |
0,0001 |
0,5094 |
0,5750 |
0,1323 |
0,6837 |
0,9610 |
0,6438 |
|
EL |
0,0394 |
0,0959 |
0,2714 |
0,9004 |
0,0102 |
0,0024 |
0,8767 |
0,0621 |
0,2109 |
|
EW |
0,0039 |
0,0074 |
0,0002 |
0,6409 |
0,3313 |
0,6890 |
0,2444 |
0,5388 |
0,5533 |
|
L |
0,0051 |
0,0172 |
0,0277 |
0,8378 |
0,0088 |
0,0024 |
0,9381 |
0,1622 |
0,2556 |
Примечание: * - использовался тест Стьюдента. Жирным шрифтом выделены значения /?<0,05.
Средние значения линейных размеров S^ с участка 3 в большинстве случаев несколько превосходили размеры 5^ с участков 1 и 2. Однако статистически значимые различия отмечались только по ширине головы (HW) -по сравнению с участка 2 и ширине переднего края переднеспинки (РА) - по сравнению с S^ участка 1.
При сравнении относительных параметров тела S^ можно отметить только различие в пропорциях головы (HW/HL и HL/HW), которая у Ц^’ с участка 1 относительно более широкой, чем с участка 2.
По общим размерам тела ^ во всех случаях не только несколько крупнее ^ = но У них также наблюдался более широкий диапазон колебаний этих размеров. В качестве примера на рис. 3 приведены показатели об-
Уровень достоверности различий р между выборками различных участков
Таблица 5
|
Параметр |
Сравниваемые выборки (цифра - номер участка) |
||||||||
|
д vs ? 1 |
vs "^ 2 |
vs ¥ 3* |
О 1 vs |
О 2 vs |
6 3 vs |
^ 1 vs ?2 |
^2 vs ?з |
^ 3 vs ?1 |
|
|
HW/HL |
0,0069 |
0,1932 |
0,2913 |
0,5618 |
0,0561 |
0,0650 |
0,0361 |
0,0727 |
0,9740 |
|
HL/HW |
0,0065 |
0,1970 |
0,3009 |
0,5618 |
0,0561 |
0,0650 |
0,0361 |
0,0629 |
0,9696 |
|
PAZPLm |
0,5419 |
0,5369 |
0,3835 |
0,5167 |
0,0561 |
0,3099 |
0,5737 |
0,6793 |
0,2272 |
|
PW/PLm |
0,2005 |
0,3486 |
0,0897 |
0,0491 |
0,0199 |
0,7973 |
0,7562 |
0,8576 |
0,5978 |
|
PB/PLm |
0,7642 |
0,9473 |
0,7111 |
0,5094 |
0,5018 |
0,9571 |
0,4492 |
0,5000 |
0,9480 |
|
PLm/PA |
0,5687 |
0,4873 |
0,3557 |
0,4949 |
0,0761 |
0,2640 |
0,5605 |
0,6092 |
0,1827 |
|
PLm/PW |
0,1556 |
0,4402 |
0,0744 |
0,0637 |
0,0325 |
0,8531 |
0,7269 |
0,9048 |
0,6283 |
|
PLm/PB |
0,8415 |
0,7998 |
0,6115 |
0,5167 |
0,5399 |
0,9429 |
0,4151 |
0,4367 |
0,9307 |
|
EW/EL |
0,0735 |
0,0245 |
0,0006 |
0,4736 |
0,3930 |
0,1081 |
0,3417 |
0,4177 |
0,0887 |
|
EL/EW |
0,0601 |
0,0178 |
0,0001 |
0,4325 |
0,4076 |
0,0599 |
0,2993 |
0,4661 |
0,1237 |
Примечание: * - использовался тест Стьюдента. Жирным шрифтом выделены значения /<0,05.
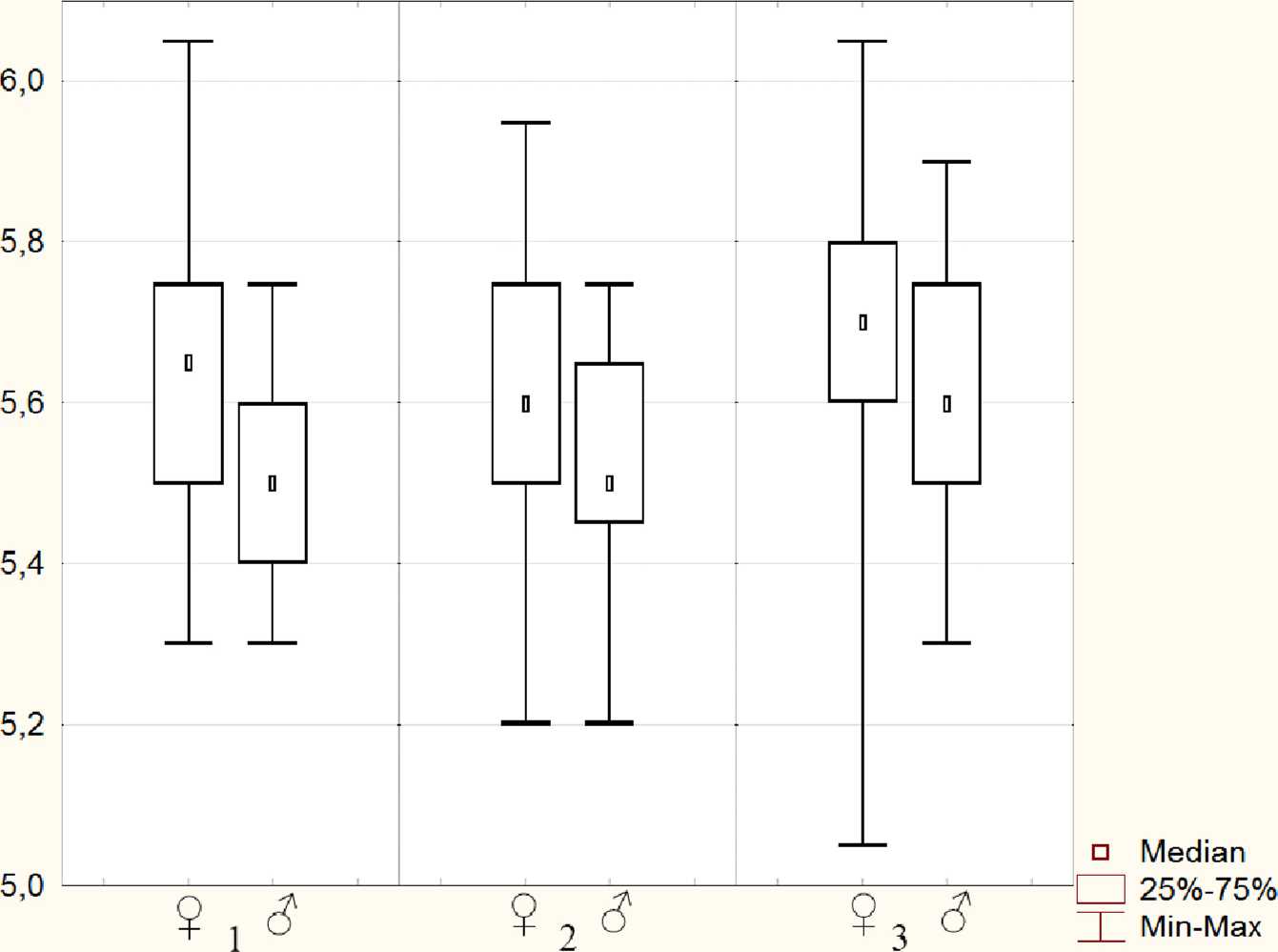
Рис 3. Диапазон колебаний показателя общей длины тела (L) для выборок с участков 1, 2 и 3: По вертикали - значения в мм, на горизонтальной оси цифрой обозначен номер участка
щей длины тела (L) самцов и самок N.reitteri с участков 1,2иЗ.
Заключение
Подтверждена биотопическая приуроченность N. reitteri к бореальным лесам.
Во всех проанализированных выборках выявлены достоверное большие абсолютные размеры тела самок.
Статистически значимые различия между выборками N. reitteri из разных участков наблюдались только по размерам (5' • Различия между вариабельны и, как правило, незначительны.
Наличие достаточно высокой вариабельности в размерах A. reitteri может косвенно свидетельствовать о его значительной экологической пластичности.
Исследование выполнено при финансовой поддержке конкурсных проектов ДВО РАН (12-1-П30-14, 12-П-СУ-06-007).
Список литературы Половой диморфизм размеров тела у одного из видов жужелиц (Notiophilus reitteri Spaeth)
- Будилов П.В. Морфометрические признаки полового диморфизма популяции Notiophilus reitteri Spaeth на территории Буреинского заповедника//Экология, эволюция и систематика животных: мат-лы Всерос. науч.-практич. конф. с междунар. участием. Рязань, 17-19 ноября 2009 г. Рязань, 2009. С. 305-306.
- Думикян А.Д., Ривкус Ю.З., Триликаускас Л.А. Буреинский заповедник -край девственной природы//Приамурское географическое общество. Хабаровск, 2005. С. 26-28.
- Зиновьев Е.В. Материалы по фауне жесткокрылых Среднего Приобья//Вестник ОГУ. Приложение. 2006. № 4. С. 44.
- Компоненты экосистем и биоразнообразие карстовых территорий Европейского Севера России (на примере заповедника «Пинежский»)/отв. ред.: Л.В. Пучнина. Архангельск, 2008. С. 252-253.
- Крыжановский О.Л. Фауна СССР. Жесткокрылые. Л.: Наука, 1983. Т. 1. Вып. 2. С. 73-74.
- Лафер Г.Ш. Семейство Carabidae -Жужелицы//Определитель насекомых Дальнего Востока СССР. Т. 3. Ч. 1. Л.: Наука, 1989. С. 71-222.
- Сундуков Ю.Н. Жужелицы (Coleoptera, Carabidae) высокогорий Южного Сихотэ-Алиня//Чтения памяти Алексея Ивановича Куренцова. Вып. 21. Владивосток: Дальнаука, 2010. С. 35-40.
- Сундуков Ю.Н. Жужелицы (Coleoptera, Carabidae) национального парка «Зов тигра» (Приморский край, Россия)//Евразиатский энтомологический журнал. 2011. № 10 (4). С. 440.
- Шиленков В.Г., Ананина Т.Л. Материалы по фауне жужелиц (Coleoptera, Carabidae) Баргузинского заповедника//Биоразнообразие Байкальского региона. Труды Биолого-почвенного факультета ИГУ. 2001. Вып. 5. С. 29.
- Barљevskis A. Biogeography of the genus Notiophilus Dumeril, 1806 (Coleoptera: Carabidae). Baltic J. Coleopterol. 2007. N 7 (1). P. 121-135.
- Kryzhanovskij O.L. et al. A checklist of the ground-beetles of Russia and ajancent lands. 1995.
- Lindroth C.H. Ground Beetles (Carabidae) of Fennoscandia A Zoogeographic Study PART I, Specific Knowledge Regarding the Species//Smithsonian Institution Libraries and The National Science Foundation Washington, D.C. 1992. 507 p.
