Популяционная экология большой синицы Parus major в таежных лесах Карелии. Часть 1. Структура населения и особенности гнездования

Автор: Артемьев Александр Владимирович
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 2 (92), 2008 года.
Бесплатный доступ
На основе исследований 1979-2004 гг. подробно описана популяционная экология большой синицы в Карелии. Приведены сведения по структуре гнездового населения, биологии гнездования и динамике численности птиц. Выявлены существенные отличия исследованной популяции от популяций центральной и западной Европы, в первую очередь по характеру действия зависящих от плотности механизмов регуляции численности, степени воздействия на птиц погодных условий и ряду других параметров. Выявлены и проанализированы основные факторы, определяющие динамику обследованной популяции.
Большая синица, таежные леса, плотность населения, биология гнездования, механизмы регуляции численности
Короткий адрес: https://sciup.org/14749398
IDR: 14749398 | УДК: 575.857:598.841(470.22)
Текст научной статьи Популяционная экология большой синицы Parus major в таежных лесах Карелии. Часть 1. Структура населения и особенности гнездования
Ареал номинального подвида большой синицы включает в себя почти всю Европу и значительную часть Азии, по широте он растянут от Турции до севера Скандинавии, а по долготе – от Атлантиче ского океана до побережья Охотского моря [1]. Условия обитания птиц на такой обширной территории самые разнообразные, и поэтому ответ разных популяций на изменение внешних факторов может существенно различаться. В этой связи большой интерес представляет сравнительный анализ экологии и динамики популяций из разных частей ареала для выявления как общих закономерностей, так и особенностей, связанных с локальными условиями. Большинство детальных исследований по биологии большой синицы проведено на птицах номинального и британского подвидов в европейских широколиственных и сме- шанных лесах [2, 3, 4, 1, 5 и др.]. Настоящая работа посвящена подробному анализу экологии популяции в лесах таежной зоны Европейской Ро ссии, в условиях, существенно отличающихся от центральных частей видового ареала по климату, характеру растительности и ряду других параметров.
МАТЕРИАЛ И МЕТОДИКА
Работа выполнена в 1979–2004 гг. на стационаре Маячино Института биологии Карельского научного центра РАН (60° 46 ′ с. ш., 32° 48 ′ в. д.), расположенном на юго-восточном берегу Ладожского озера (Олонецкий район Карелии). Район исследований относится к подзоне средней тайги. Ландшафт здесь сильно заболоченный: древостои с сухими почвами занимают 8 % площади, а основная их часть расположена на свежих и влажных или сырых и мокрых поч-
вах. Среди лесов преобладают сосняки, они занимают 65 % лесопокрытой площади, ельники занимают 16 %, осинники – 10 %, березняки – 8 % и ольшанники – 1 %. Сильная антропогенная трансформация территории привела к существенному изменению возрастной структуры древостоев, при этом их породный состав за последние 125–135 лет претерпел меньшие изменения, т. к. на местах рубок сосновых лесов обычно вырастают сосновые и сосново-лиственные молод-няки, а ельники восстанавливаются через стадию лиственного леса [6].
Для обследуемой территории и всего региона характерна продолжительная, но относительно мягкая зима и прохладное лето. Снежный покров лежит около 125 дней, температуры выше 0ºС держатся в среднем 205 дней в году (с 10 апреля по 1 ноября), а выше +5ºС – 160 дней (с 29 апреля по 6 октября). Апрель довольно прохладный, средняя многолетняя температура этого месяца на широте Петрозаводска всего 1.2ºС, что не сколько ниже температуры самого холодного месяца года на юге Англии, в Голландии, в равнинных частях Швейцарии и некоторых других областях ареала большой синицы [7]. Весенняя погода неустойчивая, что обусловлено чередованием вторжений воздушных масс из Атлантики и Арктики [8, 9]. В связи с этим здесь отмечается повышенная, по сравнению с центральными областями России, нестабильность сроков многих фенологических явлений [10].
Для привлечения птиц использовались дощатые синичники обычных размеров [11]. Они были вывешены в характерных для региона типах леса, основная масса в спелых сосновых (7 С, 2 Е, 1 Б) и сосново-березовых лесах разного возраста (5 С, 5 Б), часть в спелых ельниках (7 Е, 2 С, 1 Б), елово-сосновых (4 Е, 3 С, 2 Б, 1 Ол) и лиственно-хвойных лесах (3 Б, 2 Ол, 2 Ос, 2 Е, 1 С) и черноольшанниках (8 Ол, 1 Е, 1 Б). Первые 150 синичников вывешены в 1979 г., еще 200 штук – в 1980, в последующие годы число пригодных для синиц гнездовий изменялось от 270 до 400 (рис. 1). Большинство из них было размещено линиями вдоль дорог, квартальных просек и мелиоративных канав, с интервалом в 20–100 м, в среднем около 40 м. Кроме того, были заложены 3 площадки по 15, 25 и 50 гнездовий с плотностью развески около 10 синични-ков на 1 га. Участки развески располагались на территории около 10 км2 в сплошном лесном массиве в 25 км от небольшого города Олонца. Для оценки плотности гнездового населения учитывали только площадь ближайших окрестностей гнездовий. При развеске линией в расчет включали 100 м полосу вдоль нее (по 50 м с каждой стороны), а к ее протяженности добавляли 100 м (по 50 м от крайних синичников). При развеске площадками в расчет включали 50 м полосу, прилегающую к крайним рядам гнездовий. С 1980 г. площадь, занятая гнездовьями, составляла 120–170 га, в среднем – 145 га. Данные о числе гнездовий по биотопам и их заселенности большой синицей приведены в табл. 1.
Ежегодно прослеживали судьбу всех гнезд, фиксировали сроки начала и величину кладки, успешность инкубации и выкармливания, проводили практически сплошное мечение птенцов. Взрослых птиц (87 % самок и 84 % самцов) отлавливали на гнездах во время выкармливания 7–14 - дневных птенцов и метили стандартными кольцами. Их возраст определяли по контрасту кроющих перьев крыла [12, 13].
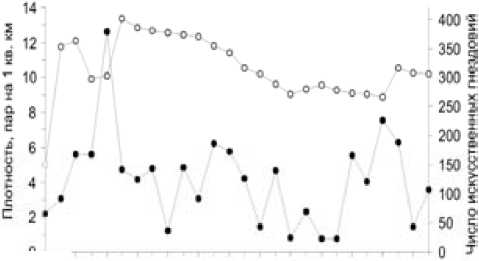
-*- плотность гнездового населения -о- число ИГ
Рис. 1.Число искусственных гнездовий и плотность гнездового населения большой синицы в Приладожье в 1979–2004 гг.
Таблица 1
Заселенность искусственных гнездовий и плотность гнездового населения большой синицы в разных биотопах Приладожья в 1979–2004 гг.
|
Биотопы |
о S |
is ° и С w 2 |
л о ч к & U с |
§. с? я с К о о 5 н 2 d о сз со к к |
н о r'U н ^ 2 С с |
|
Черноольшанники |
664 |
318.8 |
11 |
1.66 |
3.45 |
|
Ельники |
909 |
636.9 |
6 |
0.66 |
0.94 |
|
Елово-сосновые леса Лиственно-хвойные |
542 |
239.4 |
9 |
1.66 |
3.76 |
|
леса |
1991 |
720.6 |
54 |
2.71 |
7.49 |
|
Сосняки |
871 |
563.6 |
9 |
1.03 |
1.60 |
|
Сосново-лиственные |
1931 |
713.6 |
48 |
2.49 |
6.73 |
|
Молодняки сосноволиственные |
348 |
154.8 |
6 |
1.72 |
3.88 |
|
Приспевающие сосново-лиственные |
655 |
240.1 |
8 |
1.22 |
3.33 |
|
Всего за 1979–2004 гг. |
7911 |
3587.8 |
151 |
1.91 |
4.21 |
В 1979–2004 гг. в синичниках гнездилось 159 пар птиц (от 1 до 18 пар в год), у них была прослежена судьба 235 гнезд. Ежегодные сроки начала размножения определяли по появлению 1 яйца в наиболее раннем гнезде. Средние ежегодные даты начала размножения популяции рассчитывали по принятой у западноевропейских орнитологов методике [14] – по кладкам, начатым в течение 30 дней после появления наиболее ранней в сезоне, без учета повторных и вторых гнезд. Успешность размножения оценивали по относительному числу гнезд, сохранившихся до вылета птенцов, а также по соотношению числа слетков, покинувших синични-ки, и суммарного числа отложенных яиц. Отдельно определяли эмбриональную смертность (отход яиц без учета погибших кладок) и частичный отход птенцов (без учета погибших выводков). Общую успешность насиживания и выкармливания оценивали с учетом гибели кладок и выводков. Среднюю величину выводка рассчитывали только для гнезд, из которых вылетел по крайней мере 1 птенец. Продуктивность размножения определяли по соотношению числа слетков, покинувших синичники, и числа гнездившихся в них самок. Предварительные итоги исследований гнездовой биологии опубликованы ранее [15].
Принято считать, что изменения численности птиц в искусственных гнездовьях отражают реальное состояние популяции на сопредельных участках леса. Для исследуемого региона характерна очень низкая плотность населения большой синицы (табл. 1), из-за чего объем собранного материала по годам был невелик, несмотря на значительную площадь контролируемой территории и большое число синичников, и не всегда мог служить основой для полноценного статистического анализа. Поэтому некоторые заключения относительно отдельных факторов, влияющих на динамику местного населения, основаны на обнаруженных тенденциях, и их следует рассматривать как предварительные, требующие дополнительной проверки.
Для характеристики погоды использованы данные метеостанции Олонец, расположенной в 25 км к северо-востоку от центра территории, где были развешаны гнездовья: среднемесячные и суточные температуры воздуха. Следует отметить, что эти показатели не дают точного представления о микроклимате в гнездовых стациях, а лишь отражают общую тенденцию изменений погоды в районе исследований.
При обработке данных пользовались обычными методами вариационной статистики [16, 17]. Для оценки связи и зависимости переменных применяли корреляционный анализ (коэффициент Браве-Пирсона) и пошаговый многофакторный регрессионный анализ. При оценке различий признаков, выраженных в процентах, пользовались критерием Фишера с фи-преобразованием.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Плотность гнездового населения
На участках развески искусственных гнездовий плотность населения большой синицы колебалась по годам от 0.8 до 12.6 пары/км2 и в среднем составляла 4.2 пары/км2 (рис. 1). Наиболее охотно птицы заселяли синичники, вывешенные в спелых хвойно-лиственных лесах со значительной примесью или преобладанием (40–70 %) мелколиственных пород: березы, осины, ольхи (табл. 1). Максимальная для района исследований плотность населения зарегистрирована в спелых сосново-лиственных лесах: во время пика численности в 1983 г. она составила 24.4 пары/км2 (8 пар на 94 гнездовья). Несколько реже птицы заселяли чистые черноольшанники, сосново-лиственные приспевающие древостои и молодняки, а также хвойные леса с небольшой примесью лиственных пород. Наименее привлекательными для них были чистые ельники и сосняки, в которых они в годы низкой численности (менее 3 пар/км2) не селились вовсе.
Как и в других частях ареала [2, 18, 4, 19, 1 и др.], в Приладожье прослеживается характерное для большой синицы предпочтение древостоев с преобладанием лиственных пород. Исключение со ставляют спелые черно-ольшанники, в которых птицы селятся менее охотно, чем в березово-хвойных лесах, что связано с поздними сроками распускания листьев на ольхе и малой привлекательностью этого биотопа ранней весной.
В условиях северо-запада России большая синица предпочитает гнездиться в лесах, расположенных поблизости от зимовочных стаций. Поэтому максимальная плотность гнездового населения в Ленинградской области (30– 40 пар/км2) и в Карелии (28 пар/км2) отмечается в смешанных древостоях в окрестностях населенных пунктов или в старых городских парках [20, 21]. Известно, что в исследуемом регионе в удаленных от жилья человека лесах синицы обычно не зимуют и с невысокой плотностью заселяют их в гнездовой период, поэтому в оптимальных для птиц стациях, в окрестностях крупных городов, они занимают 80–90 % искусственных гнездовий, а в глубине лесных массивов – только 2–10 % [22, 23]. В естественных условиях в лесах Приладожья плотность населения этого вида невысока, в преобладающих здесь сосняках разного типа она колеблется от 0.4 до 6.3 пар/км2 [6]. Судя по данным табл. 1, развеска гнездовий в таких местообитаниях не приводит к заметному увеличению численности птиц. Несмотря на то что синицы охотно селятся в искусственных гнездовьях, с равной вероятностью они выбирают для размножения и свободные естественные дупла. В отличие от мухоловки-пеструшки, плотность населения которой во многом определяется наличием пригодных для гнездования дупел и резко возрастает при развеске синичников, у большой синицы этот фактор не оказывает существенного влияния на динамику популяции. В удаленных от населенных пунктов лесах таежной зоны плотность гнездового населения этих птиц невысока, поэтому они не испытывают недостатка в свободных дуплах, и только в молодняках развеска гнездовий ведет к росту численности синиц [19].
Структура гнездового населения
Анализ возрастного состава гнездового населения показывает, что его основа формируется из птиц-первогодков, птицы старших возрастных групп составляют 48.4 % среди самцов и 24.4 % среди самок (рис. 2). Сходное соотношение возрастных групп наблюдается на побережье Ладожского озера и в предгнездовой период в марте–апреле, когда доля первогодков составляет 53 % среди самцов и 72 % среди самок [22]. Очевидно, наши данные отражают реальное распределение синиц по возрасту в лесах Приладожья. По происхождению большинство гнездящихся в синичниках птиц были иммигрантами, впервые поселяющимися здесь и родившимися за пределами контролируемой территории. Особи, обитавшие на контролируемой территории ранее, – резиденты – составили всего 8.3 %, а птицы местного происхождения – лишь 2.5 % населения (рис. 3). По этому соотношению обследуемая популяция сильно отличается от оседлых популяций европейских широколиственных лесов, где иммигранты составляют от 20 % до 57 % гнездового населения, а на долю особей местного происхождения обычно приходится более 25 % [2, 4, 24, 25].
Сроки размножения
В Приладожье большие синицы обычно приступают к размножению в конце апреля – начале мая. Начало первой в сезоне кладки в разные годы регистрировалось с 22 апреля (1983 г.) по 25 мая (1997 г.), в среднем – 7 мая. Весь период начала размножения растянут почти на 3 месяца, как за счет наличия второго цикла гнездования у значительной части птиц, так и за счет кладок возобновления после гибели гнезд (рис. 4).
На большей части ареала для большой синицы характерно бицикличное размножение [26, 1]. В районе исследований ежегодно от 0 до 100 %, в среднем 47,8 % (n=116), пар после успешного выкармливания первого выводка приступало ко второй кладке.
По нашим данным сроки начала кладок первого цикла в разные годы колеблются с 22 апреля по 26 мая (средняя дата – 8 мая, n=65), вторых – с 6 июня по 15 июля (средняя дата – 23 июня, n=55). Повторные кладки отмечались в период с 1 мая по 25 июля, после гибели как первых, так и вторых гнезд. В сходные сроки птицы приступают к гнездованию и в других пунктах юга
Карелии: в районе 61.5–62.5o с. ш. начало размножения длится с 30 апреля по 18 июля [19].
Средняя за 1979–2004 гг. дата начала кладок, появившихся в течение 30 дней после появления наиболее ранней в сезоне, – 10 мая (n=145).
Данные по срокам гнездования вида в южной Карелии укладываются в общую схему, описанную Гопкинсом [27, 28], сроки фенологических явлений при продвижении к северу в целом запаздывают.
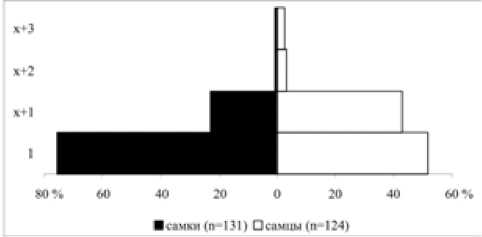
Рис. 2. Возрастная пирамида гнездового населения большой синицы в Приладожье в 1979–2004 гг.
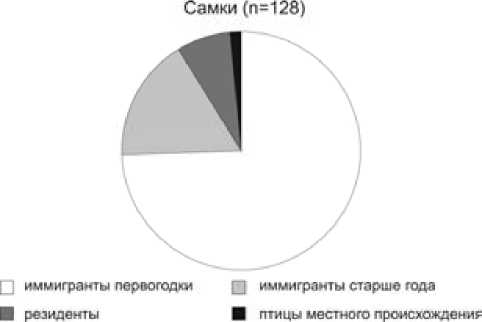
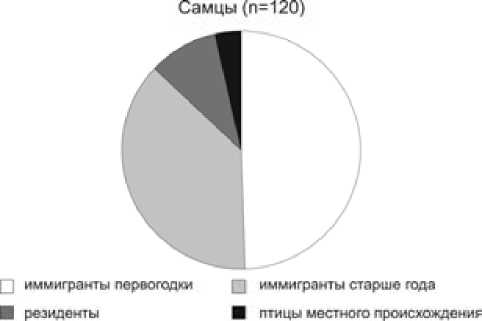
Рис. 3. Структура гнездового населения большой синицы в Приладожье в 1981–2004 гг.
В то же время анализ материалов по изменени-° ям средних дат гнездования птиц на широте 60–61° показывает, что существенных сдвигов времени размножения по долготе нет (табл. 2), несмотря на более мягкий климат и высокие весенние температуры в западных областях ареала.
Известно, что в разных частях ареала сроки гнездования синиц во многом определяются ходом температур воздуха, а также развитием весенних фенологических явлений [2, 27]. В При-ладожье наиболее значимая корреляция сроков гнездования птиц выявлена со среднемесячной температурой апреля (табл. 3), тогда как связи с температурами других месяцев не обнаружено. Весной на вегетацию растительности умеренных широт наиболее сильно действуют так называемые «эффективные» температуры воздуха – превышающие +5ºС [31]. Поэтому нами предпринята попытка определить связь сроков размножения птиц с ходом температур, превышающих +5ºС. Наиболее тесная корреляция выявлена между сроками появления первой в сезоне кладки и средней даты начала гнездования популяции с датой накопления суммы эффективных температур 50ºС, хотя интервал между этими событиями сильно варьировал в разные годы. Чуть более слабая связь названных параметров выявлена и с датой устойчивого перехода минимальных температур через 0ºС, и с датой накопления суммы положительных температур 100ºС.
Известно, что сроки гнездования птиц умеренных и высоких широт контролируются эндогенной программой, которая запускается при пороговом для каждого вида изменении фотопериода [32, 33]. У большой синицы выявлены популяционные различия фотопериодических
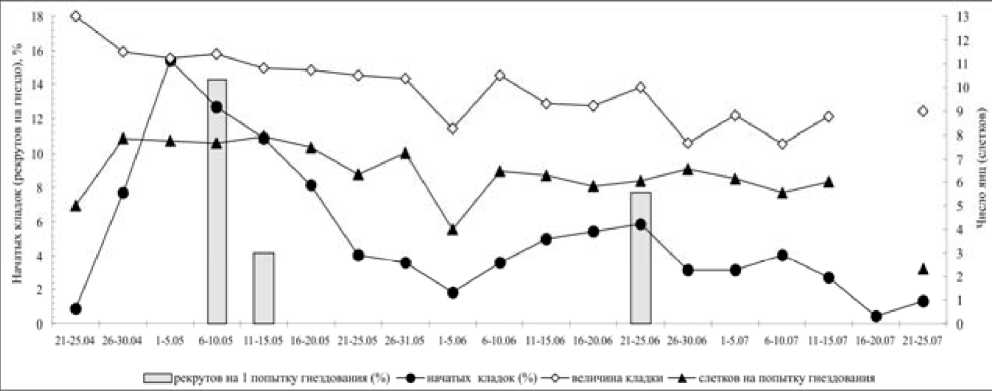
Рис. 4. Влияние сроков гнездования на величину кладки, выводка и число рекрутов у большой синицы в Приладожье в 1979–2004 гг.
Таблица 2
Основные параметры размножения большой синицы в европейской части ареала на широте 60–61o
|
Место и годы исследований |
Координаты и высота над ур. моря |
Первые и ранние кладки |
Число вторых кладок, % |
Автор |
|
|
средняя дата начала (1–1 мая) |
среднее число яиц |
||||
|
Норвегия, Серкедал 1968– 1974 |
60° с.ш., 11° в.д.; 200 м |
11.7 (35)* |
– |
– |
Slagsvold, 1976 [27] |
|
Швеция, Тэрнсйо 1952– 1963, 1972–1974. |
60°19 ′ с.ш., 16° 56 ′ в.д.; 200 м |
– |
8.2 (400) |
18.5 |
Johansson, 1974 [29] |
|
Финляндия, Лемсйохолм 1953–1967 |
60° 30 ′ с.ш., 22° в.д.; 50 м |
10.7 (225) |
9.9 (225) |
20.2 |
Haartman, 1969 [30] |
|
Россия, Карелия, Маячино 1979–2004 |
60° 46 ′ с.ш., 32° 48 ′ в.д.; 20 м |
10.1 (141) |
11.1 (122) |
47.8 |
Наши данные |
* в скобках – число гнезд
Таблица 3
Связь сроков гнездования большой синицы и некоторых показателей весенней погоды в Приладожье в 1980–2002 гг.
О влиянии температуры на сроки гнездования известно давно, но до сих пор нет общего мнения относительно того, как она действует на птиц: непосредственно на физиологию и развитие репродуктивного состояния или на бюджет энергии через развитие растительности и кормовой базы. Ван Бален [38] установил, что температура действует опосредованно, и синицы реагируют не на нее, а на сроки вылупления из яиц и обилие мелких гусениц. Позднее в экспериментах на скворцах Мейер с соавторами [39] подтвердили предположение Клюйвера [2] о прямом действии температуры на сроки начала кладки. По мнению Нагера [40], в природе этот фактор оказывает на птиц как прямое, так и опосредованное влияние. При этом ответная реакция на него особей из разных популяций существенно различается. В Голландии и Великобритании на сроки размножения больших синиц влияли температуры за 2 месяца до начала кладки, в Швейцарии и Германии – за 1.5 месяца, а в Скандинавии и Карелии лишь за предшествующий месяц [2, 38, 4, 30, 27, 40, 41, наши данные]. Везде гнездование обычно отмечалось после кратковременного потепления, но температурные пороги начала размножения в разных точках ареала различались, понижаясь в направлении с юго-запада на северо-восток [42, 43, 27, 44]. Даже в одной местности они сильно варьируют по годам, так что в условиях сурового и нестабильного климата практически невозможно выделить строгий температурный показатель старта размножения популяции [26, 5, 36, наши данные]. В Приладожье в разные годы в день начала 1 в сезоне кладки среднесуточная температура воздуха колебалась от 2.2 до 14.3ºС, а в среднем составила 9.0ºС. «Дата детерминанта» (determinant date) – показатель, предложенный Клюйвером [2] как условное время старта быстрого развития ооцитов у большинства самок (день за 4 суток до средней даты начала кладки популяции), – в районе исследований в разные годы отмечалась с 27 апреля по 21 мая (в среднем – 8 мая). В этот де°нь температура воздуха составила в среднем 8.7°С, однако пороговой ее назвать нельзя, так как по годам она колебалась от –0.4 до 14ºС. Срок накопления суммы эффективных температур 50ºС совпадал с «датой детерминанта» и в среднем наступал 8 мая с вариациями по годам от 23 апреля до 19 мая. И хотя различия во времени наступления этих событий в отдельные годы достигали 9 дней, данный фенологический показатель наиболее значимо влиял на птиц исследуемой популяции.
Анализ данных по Карелии не выявил значимых долгосрочных изменений сроков гнездования синиц. Глобальное потепление климата, существенно влияющее на сроки размножения птиц Западной Европы [45, 46, 14], пока еще не сказывается на биологии гнездования большой синицы в Приладожье. Это связано с тем, что в нашем регионе за последние 50 лет более теплой стала зима и повысились температуры воздуха в марте [47], в то время как апрельские и майские температуры, наиболее сильно влияющие на весеннюю фенологию, изменились незначительно. Так за период 1980–2002 гг. среднемесячная температура апреля имела незначимую тенденцию к росту (r= 0.35 n. s.), а температура мая – такую же обратную (r= –0.28 n. s.). Соответственно и сроки гнездования птиц за этот период практически не изменились (r= –0.21 n. s.). Подобное явление отмечено в Подмосковье и в северной Финляндии, где у большой синицы, по сравнению с более западными частями ареала, предгнездовой период также не стал заметно теплее [48].
Сроки размножения большой синицы в разных биотопах Приладожья довольно близки, поэтому на наших материалах значимых различий между лесами разного типа не выявлено (табл. 4). Однако суммарно птицы, селящиеся в практически чистых ельниках и сосняках, приступали к гнездованию в среднем на 7 дней позже, чем в смешанных и лиственных древостоях (F=6.5, p<0.05). Вероятно, это связано с тем, что ранней весной хвойные леса менее проницаемы для солнечных лучей, снег здесь сходит несколько позднее, почва прогревается медленнее и фенологическое развитие растительности запаздывает по сравнению с другими стациями. В других пунктах южной Карелии В. Б. Зимин
(1978) [19] обнаружил значимые различия только между спелыми лиственными древостоями и сосново-лиственными молодняками. Следует отметить, что биотопические различия в сроках размножения не всегда отчетливо проявляются и в других частях ареала. В центральной Европе более раннее гнездование отмечено в городских парковых насаждениях по сравнению с лесами и древостоями в сельской местности [1]. В Англии в садах синицы начинают гнездиться раньше, чем в лесах и парках, а в хвойных и лиственных древостоях сроки размножения не различаются [49]. Такую же тенденцию выявил Клюйвер (1951) [2] в Голландии, однако позднее там были обнаружены различия в сроках размножения птиц в дубовых и сосновых лесах [38]. В Средиземноморье в листопадных дубовых лесах птицы начинают гнездиться раньше, чем в вечнозеленых [50]. В целом подобные различия чаще всего проявляются в контрастно различающихся местообитаниях, причем в каждом случае они обусловлены действием разных локальных факторов, таких как искусственное освещение в городах, дополнительная подкормка птиц в садах или особенности фенологии и динамики кормовой базы в естественных стациях.
В Приладожье у самок большой синицы слабо выражены возрастные различия сроков размножения (табл. 5). Как и многие другие воробьиные птицы таежной зоны [36], синицы разного возраста приступают к гнездованию практически одновременно, и основная часть популяции ежегодно начинает размножение в сжатые сроки. Известная закономерность раннего гнездования старых самок по сравнению с первогодками в Карелии и Финляндии не подтверждается, более того, имеет место обратная тенденция: самки-первогодки приступают к откладке яиц наиболее рано [36, 30, наши данные].
Разница в датах начала кладки годовалых и более старых птиц в Приладожье и южной Финляндии (Лемсйохолм) в среднем составляла 1 день, но из-за небольшого объема материалов не была значимой. В Западной Европе самки старших возрастных групп, как правило, гнездятся раньше первогодков, причем в южных частях ареала эта разница в сроках увеличивается. Так в Швейцарии она достигала 4.2 дней [37], в Голландии отмечалась не ежегодно и составляла 2.1 дня [2], в южной Англии около 1 дня [4], а в южной Швеции и в Польше небольшие различия проявлялись лишь в отдельные годы [51].
На сроки размножения птиц в районе исследований существенное влияние оказывал и возраст самца: пары с самцами-первогодками обычно гнездились на 1–4 дня позже, чем с более старыми партнерами (табл. 5). И хотя в Карелии эта тенденция прослеживалась во всех комбинациях возраста партнеров, значимые различия выявлены только между группами «старый самец – самка-первогодок» и «самец-первогодок – самка любого возраста» (t=2.01; p<0.05). По-видимому, возрастные различия сроков гнездования, отчетливо проявляющиеся в более южных частях ареала [52, 53], в Приладо-жье сглаживаются и затушевываются на фоне действия внешних факторов, в первую очередь нестабильной весенней погоды.
Известно, что сроки размножения синиц приурочены ко времени максимального обилия корма для птенцов, но они также зависят и от обеспеченности пищей самки в период кладки [3, 49, 4]. На размножение птиц влияют качество гнездового участка и индивидуальные характеристики самца: его половая активность, стимулирующая развитие репродуктивной системы партнерши, и интенсивность кормлений ухаживания, служащих дополнительным источником энергии для самки в период формирования кладки. Обычно самцы
Таблица 4
Основные показатели первого цикла размножения и частота вторых кладок в разных биотопах Приладожья в 1979–2004 гг.
|
Биотопы |
Дата начала кладки (1 – 1 мая) |
Число гнезд |
Величина первой кладки |
Величина первого выводка |
Число успешных первых кладок |
Число вторых кладок, % |
|
Черноольшанники |
9.8±2.1 |
10 |
10.9±0.7 |
8.8±0.5 |
8 |
25 |
|
Ельники |
14±3.7 |
5 |
11±0.6 |
8.5±0.5 |
4 |
50 |
|
Елово-сосновые леса |
6±4.5 |
6 |
11.6±0.2 |
11.2±0.2 |
5 |
20 |
|
Лиственно- хвойные леса |
9.4±1.3 |
50 |
11.1±0.2 |
9.4±0.3 |
39 |
46.2 |
|
Сосняки |
16.4±3.1 |
9 |
10.9±0.5 |
8.9±0.5 |
7 |
42.9 |
|
Сосново-лиственные леса |
7.7±1.2 |
46 |
11±0.2 |
8.9±0.4 |
42 |
54.8 |
|
Молодняки сосноволиственные |
10.3±3.8 |
6 |
10. 7±0.4 |
9.2±0.4 |
5 |
80 |
|
Приспевающие сосново-лиственные |
8.8±1.5 |
8 |
11.5±0.4 |
9.5±0.9 |
6 |
50 |
Таблица 5
Возраст партнеров и основные показатели первого цикла размножения
|
Возраст |
Число пар |
Дата начала кладки |
Величина полной кладки |
Величина выводка |
||||
|
самки |
самца |
пределы |
средняя |
пределы |
средняя |
пределы |
средняя |
|
|
1 год |
все |
87 |
25.04-1.06 |
8.90* ±0.92 |
7-13 |
11.19 ±0.14 |
4-13 |
9.42 ±0.21 |
|
x+n |
все |
29 |
22.04-29.05 |
9.97 ±1.58 |
6-15 |
10.86 ±0.33 |
4-11 |
8.33 ±0.39 |
|
все |
1 год |
58 |
27.04-1.06 |
10.45** ±1.07 |
6-15 |
10.91 ±0.21 |
4-12 |
9.09 ±0.26 |
|
все |
x+n |
56 |
25.04-29.05 |
8.11 ±1.16 |
7-13 |
11.15 ±0.17 |
4-13 |
9.22 ±0.28 |
|
1 год |
1 год |
43 |
28.04-1.06 |
10.44 ±1,26 |
7-13 |
11.07 ±0.19 |
4-12 |
9.41 ±0.28 |
|
1 год |
x+n |
40 |
25.04-27.05 |
6.95 ±1.37 |
7-13 |
11.33 ±0.22 |
4-13 |
9.43 ±0.33 |
|
x+n |
1 год |
13 |
27.04-28.05 |
11.15 ±2.27 |
6-15 |
10.54 ±0.64 |
4-12 |
8.08 ±0.62 |
|
x+n |
x+n |
12 |
2.05-22.05 |
9.92 ±2.12 |
8-11 |
10.58 ±0.26 |
6-11 |
8.55 ±0.56 |
* за 1 принято1 мая
** значимые различия выделены жирным шрифтом старших возрастных групп занимают лучшие территории и отличаются от первогодков более ранним началом половой активности. Вероятно, в условиях холодной и нестабильной весенней погоды в Приладожье самки испытывают определенные трудности в поддержании положительного энергетического баланса во время формирования кладки, и поэтому возраст самца, его индивидуальные качества и способность выбрать хорошую территорию оказывают существенное влияние на сроки начала гнездования. В условиях мягкого климата и теплых весен южной Голландии возраст самца не сказывался на сроках размножения [2]. Подобное явление наблюдалось и в южной Англии в 1947–1964 гг. [4], однако позднее связь возраста самца со сроками гнездования там проявилась [52]. В южной Швеции и в Польше в отдельные годы такая взаимосвязь также имела место [51].
Тот факт, что на наших широтах старые самки гнездятся несколько позже первогодков, может свидетельствовать о более точном определении ими сроков будущего обилия корма для птенцов. Известно, что в нестабильных условиях весны и раннего лета в таежной зоне раннее гнездование не всегда бывает наиболее продуктивным [36]. В литературе есть сведения о том, что с возрастом птицы способны корректировать сроки размножения, так как на основе собственного опыта они точнее определяют время появления пика гусениц [37]. Возможно, что самки-первогодки приступают к размножению максимально рано, как только им позволяет физиология и состояние кормовой базы, а птицы старших возрастных групп начинают гнездиться в наиболее благоприятные для будущего потомства сроки.
Величина кладки
Снижение величины кладки в течение сезона размножения, отмеченное во многих частях ареала, до недавнего времени считалось правилом для большой синицы [2, 4, 54, 45 и др.]. Позднее было обнаружено, что в некоторых биотопах кладка увеличивается к середине или к концу сезона, и тренд ее изменений, как и у ряда других видов птиц, зависит от внешних условий и от особенностей популяции [38, 1, 55].
В условиях Карелии величина полной кладки у большой синицы в течение сезона уменьшалась (рис. 4). Эта зависимость удовлетворительно описывалась уравнением линейной регрессии y=11.51-0.043x (R2=33.5 %), где y – число отложенных яиц, а x – дата начала кладки (1 – 1 мая). При этом величина кладок, начатых в апреле-мае в меньшей степени зависела от календарных сроков (r= -0.28, p<0.001), чем поздних, отложенных в июне-июле (r= -0.46, p<0.001), что можно объяснить нестабильностью погодных и кормовых условий в начале сезона размножения.
У пар, дающих два выводка за сезон, первые гнезда содержали 7–13, в среднем 11.2±0.1 яиц (n=61), вторые – 6–12, в среднем 9.3±0.3 яиц (n=41). Ранние кладки птиц с одним циклом размножения состояли из 6–15, в среднем из 11.0±0.2 яиц (n=64), а повторные, начатые в разные сроки взамен утраченных, – из 1–14, в среднем из 9.6±0.5 яиц (n=23). Годовые вариации среднего числа яиц в первых и ранних гнездах были связаны с датой начала первой в сезоне кладки (r=0.43 , p<0.05), но связь этого параметра со средней датой начала размножения популяции оказалась незначимой (r=0.32, n.s.).
Зависимости величины первых и ранних кладок от биотопа в Приладожье не выявлено: в лесах разного типа этот показатель менялся, но небольшие отличия во всех случаях оказались незначимыми (табл. 4). В отличие от западной и южной Европы, где биотопические различия величины кладки отчетливо проявляются [2, 4, 38, 50, 56], на севере они сглаживаются и могут, как показывают данные Л. Хаартмана [30], вовсе отсутствовать. В Карелии такие различия отмечены только между маргинальными и оптимальными для вида стациями: сосново-лиственными молодняками и спелыми лиственными лесами [19], а в Финляндии они обнаружены лишь в пригородах Оулу, на участках, существенно отличающихся по составу древостоев и плотности гнездового населения синиц [57].
Плотность гнездового населения в разных биотопах Приладожья не влияла на число яиц в первых и ранних гнездах (r=0.01, n.s.). Средняя величина кладки была прямо связана с численностью большой синицы на контролируемой территории (r=0.42, p <0.05). Как отмечалось ранее, снижения величины кладки при повышении плотности гнездового населения не наблюдается ни в южной Финляндии, ни в Карелии [30, 58, 36, наши данные]. Это связано с тем, что в лесах таежной зоны, плотность населения большой синицы значительно меньше критического уровня, при котором включаются популяционные механизмы регуляции численности, в том числе и сокращения числа откладываемых яиц. Как показал Клюйвер [2] в Голландии эти механизмы начинают действовать, когда плотность гнездового населения превышает 40 пар/км2, но в нашем регионе она не достигает этой величины даже в оптимальных стациях [21].
В западноевропейских популяциях взрослые самки продуцируют кладки большей величины, чем первогодки [2, 4, 49]. Такая тенденция сохраняется и на юге Финляндии, однако различия уменьшаются и становятся незначимыми [30]. В Приладожье самки первогодки при сходных сроках гнездования откладывали чуть больше яиц, чем старые особи, и хотя различия в величине кладки не были значимыми, в величине выводка они проявились отчетливо. При этом возраст самца на величину кладки и выводка не влиял вовсе (табл. 5). Годовые изменения возрастного состава гнездового населения не сказывались на динамике величины кладки: от доли самцов и самок старше года она практически не зависела (r=0.31 и r= -0.01, n.s.). По-видимому, в северных широтах на птиц более сильно, чем возраст, действуют другие факторы, и на их фоне возрастные различия исчезают или существенно трансформируются. В свете гипотезы оптимизации величины кладки [59, 60, 61], можно предположить, что взрослые самки более точно прогнозируют будущее состояние кормовой базы и откладывают оптимальное для текущего сезона, в то время как первогодки – максимально возможное число яиц.
Факторы, влияющие на частоту вторых кладок
В южной Карелии один из наиболее изменчивых показателей биологии этого вида, сильно варьирующий как по годам, так и по районам исследований, это – доля вторых кладок в популяции. На контролируемой территории в Прила-дожье она составляла в разные годы от 0 до 100 %, в среднем 47.8 % (n=116).
В целом в ареале прослеживается общая тенденция сокращения частоты вторых кладок при продвижении на север [28], хотя широтные различия часто сглаживаются под влиянием других факторов: особенностей биотопа, плотности гнездового населения, возрастного состава популяции и т. д. Так, например, в меридианаль-ном разрезе между 28o и 41o в. д. такой закономерности не наблюдается, а заметное снижение частоты бициклии имеет место только на крайнем севере (табл. 6). В Фенноскандии на широте наших исследований число вторых кладок увеличивается в восточных популяциях, что, возможно, связано с изменениями местных условий, в том числе характера биотопов (табл. 2).
Одним из наиболее значимых факторов, с которым связывают частоту вторых кладок, является породный состав древостоев. В западной Европе птицы чаще приступают ко 2 циклу гнездования в хвойных и вечнозеленых лесах и реже – в листопадных [1]. При этом доля вторых кладок в лиственных лесах ниже, чем в смешанных, а в смешанных – ниже, чем в сосновых [2, 4, 49]. В Карелии в заповеднике Кивач в спелых лесах с преобладанием хвойных пород деревьев, ко второму циклу гнездования приступает около 18 % птиц, в сосноволиственных молодняках Прионежья – 55.8 %, в спелых мелколиственных лесах Заонежья – около 82 % [19]. В Приладожье два цикла гнездования птицы чаще имели в лесах с разнообразным по составу пород древостоем (табл. 4). Наиболее высока частота вторых кладок в лесах разного возраста с равным соотношением сосны и березы, чуть ниже – в смешанных лесах с преобладанием лиственных пород и почти чистых сосняках, минимальна – в практически чистых черноольшанниках и ельниках, а также в еловососновых лесах с малым числом лиственных деревьев. В разнообразных по составу пород древостоях, со сходной долей хвойных и лиственных деревьев, ко вторым кладкам приступало 57 % птиц (n=53), а в однородных или бедных по составу пород лесах (черноольшанник, ельник, елово-сосновый лес) – всего 24 % пар (n=17) (F=6.2; p<0.05). Исключение составили чистые сосняки, которые, несмотря на практически монопородный состав древостоя, по частоте бициклии заняли среднее положение.
Таблица 6
Широтная изменчивость частоты вторых кладок у большой синицы в полосе между 28o и 41o в.д.
|
Место исследований |
Координаты |
Число вторых кладок, % |
Автор |
|
Мурманская обл., Лапландский заповедник |
67°54 ′ с.ш., 32°50 ′ в.д. |
единичны |
Семенов-Тян-Шанский, Гилязов, 1991 [62] |
|
Мурманская обл., Кандалакшский заповедник |
67° ′ с.ш., 32° 20 ′ в.д. |
единичны |
Бианки, Шутова, 1978 [63] |
|
Карелия, Прионежье |
61° 30 ′ с.ш., 34° 58 ′ в.д. |
55.8 |
Зимин, 1978 [19] |
|
Карелия, Маячино |
60° 46 ′ с.ш., 32°48 ′ в.д. |
47.8 |
Наши данные |
|
Ленинградская обл. пригороды Ленинграда |
59° 55 ′ с.ш., 30°30 ′ в.д. |
27.6 |
Смирнов, Тюрин, 1981 [23] |
|
Московская обл., Приокско-Террасный заповедник |
54° 50 ′ с.ш., 37° 40 ′ в.д. |
63.4 |
Лихачев, 2002 [5] |
|
Рязанская обл., Окский заповедник |
54° 46 ′ с.ш., 40° 45 ′ в.д. |
28.1 |
Нумеров, 1987 [64] |
|
Украина, Житомирская обл. |
50° 15 ′ с.ш., 28°40 ′ в.д. |
37 |
Яремченко, Болотников, 1988 [65] |
В западноевропейских популяциях большой синицы на частоту вторых кладок существенно влияет плотность гнездового населения [2 и др.]. В районе исследований подобного не наблюдалось. Несмотря на существенную разницу в плотности населения в разных биотопах, ее связи с долей птиц, имеющих 2 кладки, не было (r=0.1, n.s.). Годовые колебания численности птиц на контролируемой территории также не влияли на частоту бициклии (r= -0.1, n.s.), даже в годы, когда плотность населения была выше средней многолетней.
Известно, что участие во втором цикле размножения зависит от возраста птиц, – особи старших возрастных групп чаще приступают ко вторым кладкам, чем первогодки [2, 4, 49]. Такая тенденция имеет место и в Приладожье, однако здесь отчетливой связи частоты бициклии с возрастом птиц не прослеживается. Ежегодные колебания доли старых птиц в составе местного населения были прямо связаны с частотой вторых кладок, но коэффициент корреляции оказался незначимым (r=0.39, p=0.06). Пары, участвующие в двух циклах гнездования, чаще состояли из старых птиц, чем моноцикличные: в первых самцы старше года составляли 57 % (n=64), во вторых – 42 % (n=60), а самки – 28 % и 23 % соответственно. Но и в этом случае различия были статистически незначимы. По-видимому, в исследуемом регионе на частоту вторых кладок воздействует большее число внешних факторов, и влияние возраста проявляется здесь не столь отчетливо, как в западной и южной Европе, где условия существования птиц более стабильные.
На частоту бициклии не влияли ни сроки начала размножения популяции (r= -0.14, n.s.), ни весенняя погода, во многом определяющая эти сроки. Примечательно, что в годы с большой величиной первых и ранних кладок, число пар, приступающих ко второму циклу размножения, возрастало (r=0.57, p<0.05). Это свидетельствует о том, что и величина кладки, и частота бицик-лии, очевидно, были связаны с состоянием кормовой базы в весенний период.
Определенное влияние на долю вторых кладок оказывали погодные условия зимы. В При-ладожье после теплых зим их число обычно возрастало. Подобную тенденцию отмечал Г. Н. Лихачев [26] в Московской области. Наши материалы показывают связь на грани значимости частоты бициклии с суммой среднемесячных температур декабря, января и февраля (r=0.35, p=0.09) и значимую связь этого параметра со среднемесячной температурой февраля (r=0.41, p<0.05). В феврале у синиц начинается развитие репродуктивного состояния, и можно предположить, что погода действует на его ход. Экспериментально установлено, что физиологическое развитие репродуктивной системы у синиц северных популяций не зависит от низких температур и ускоряется при высоких [34]. Из этого следует, что в теплые сезоны птицы будут готовы к размножению раньше, чем в холодные, и соответственно изменятся сроки гнездования. Наши данные показывают, что так и происходит, но в Приладожье на сроки размножения влияют только температуры апреля (табл. 3) и не влияют ни мартовские, ни февральские (r= -0.1 n.s.). Температуры воздуха этих месяцев остаются низкими и, по-видимому, не достигают пороговых значений, ускоряющих развитие репродуктивного состояния. Кроме того, в районе исследований зимует лишь незначительная часть местного населения, а большинство птиц перемещается в зимовочные стации, иногда удаленные на сотни километров, где погодные условия совсем другие [66, 67, наши данные].
Более вероятно, что зимние температуры воздействуют на частоту бициклии опосредованно, сказываясь на состоянии кормовой базы синиц. Известно, что погодные условия зимовки существенно влияют на динамику численности насекомых [68, 69]. А состояние кормовой базы – один из важных факторов, определяющих частоту вторых кладок [70]. Вероятно, низкие зимние температуры снижают выживаемость беспозвоночных, уменьшая их обилие весной, и поэтому после холодных зим меньшее число синиц имеет два цикла размножения. Косвенным подтверждением этому служит динамика выживаемости потомства в первых и ранних гнездах – частичный отход птенцов находится в обратной зависимости от суммы среднемесячных температур декабря, января и февраля (r= -0.44, p<0.05).
В Приладожье дистанция между первым и вторым гнездами одних и тех же пар колебалась от 0 до 1350 м и в среднем составляла 228 м (n=55). Масштаб перемещений птиц в районе исследований был несколько большим, чем в других частях ареала [71, 26, 53], этому способствовала низкая плотность гнездового населения и отсутствие конкуренции за территории.
Отдельные пары перемещались на значительное расстояние, переселяясь из одного биотопа в другой, очевидно, в связи с динамикой кормовой базы в разных стациях в течение лета.
Интервал между откладкой 1 яйца в первой и второй кладках составлял от 33 до 63 дней, в среднем 46 дней (n=55), а между датами вылупления птенцов в первой и началом второй кладки – 12–38, в среднем 22 дня. Считается, что длительность промежутка между двумя циклами гнездования точнее отражает последний показатель [70]. По нашим данным, сроки начала первой кладки на него не влияли (r=0.1, n.s.), зато прямо влияла ее величина (r=0.5, p <0.001), а также дата вылупления птенцов (r=0.3, p <0.05) и величина первого выводка (r=0.35, p <0.01). Наиболее сильно этот интервал был связан со сроками начала второй кладки (r=0.75, p <0.001). Прямая связь длительности промежутка между первой и второй кладками с величиной первого выводка известна из результатов полевых экспериментов [72, 70]. Его увеличение при поздних сроках размножения служит косвенным свидетельством обеднения кормовой базы во второй половине лета. Помимо обилия и доступности корма, важным фактором, определяющим этот интервал, по-видимому, является физиологическое состояние самки, ее готовность к продуцированию новой кладки.
P. 601–647.
Список литературы Популяционная экология большой синицы Parus major в таежных лесах Карелии. Часть 1. Структура населения и особенности гнездования
- Cramp S., Perrins C. M. Flycatchers to Shrikes. The Birds of the Western Palearctic. Oxford university press,1993. V. VII. 577 p.
- Kluyver H. N. The population ecology of the Great Tit Parus m. major L. Ardea 39. 1951. P. 1-135.
- Лэк Д. Численность животных и ее регуляция в природе. М.: Изд-во иностр. лит, 1957. 404 с.
- Perrins C. M. Population fluctuations and clutch-size in the Great Tit, Parus major L. J. Animal Ecology. 1965. V. 34. P. 601-647.
- Лихачев Г. Н. Размножение и численность большой синицы (Parus major) на юге Московской области//Сибирский экологический журнал. 2002. № 6. С. 757-773.
- Волков А. Д., Громцев А. Н., Еруков Г. В., Караваев В. Н. и др. Экосистемы ландшафтов запада средней тайги (структура и динамика). Петрозаводск, 1990. 284 с.
- Алисов Б. П., Берлин И. А., Михель В. М. Курс климатологии. Ч. 3. Климаты земного шара. Л.: Гидрометеоиздат, 1954. 320 с.
- Агроклиматический справочник по Карельской АССР. Л.: Гидрометеоиздат, 1959. 184 с.
- Романов А. А. О климате Карелии. Петрозаводск: Госиздат КАССР, 1961. 140 с
- Минин А. А. Пространственно-временная изменчивость дат начала некоторых фенологических явлений у птиц на Русской равнине//Бюлл. МОИП.. Отд. биологии. 1992. Т. 97. Вып. 5. С. 28-34.
- Благосклонов К. Н. Гнездование и привлечение птиц в сады и парки. М.: Изд-во МГУ, 1991. 251 с.
- Виноградова Н. В., Дольник В. Р., Ефремов В. Д., Паевский В. А. Определитель пола и возраста воробьиных птиц фауны СССР. Справочник. М.: Наука, 1976. 189 с.
- Svensson L. Identification Guide to European Passerines. Naturhist. Riksmusset, Stockholm, 1975. 153 p.
- Both C., Artemyev A. V., Blaauw B., Cowie R. J. et. al. Large-scale geographical variation confirms that climate change causes birds to lay earlier. Proc. R. Soc. Lond. B. 2004. V. 271. P. 1657-1662.
- Артемьев А. В. Биология гнездования большой синицы Parus major в юго-восточном Приладожье//Русский орнитол. журнал. 1993. Т. 2. С. 201-207.
- Лакин Г. Ф. Биометрия. М.: Высшая школа, 1973. 343 с.
- Ивантер Э. В., Коросов А. В. Введение в количественную биологию. Петрозаводск, 2003. 304 с.
- Щербаков И. Д. Требования мухоловки-пеструшки и большой синицы к гнездовой стации и искусственному гнездовью//Пути и методы использования птиц в борьбе с вредными насекомыми. М., 1956. С. 81-93.
- Зимин В. Б. Материалы по гнездованию большой синицы (Parus major L.) в Карелии. Фауна и экология птиц и млекопитающих таежного Северо-Запада СССР. Петрозаводск: Изд. КФ АН СССР, 1978. С. 17-31.
- Мальчевский А. С., Пукинский Ю. Б. Птицы Ленинградской области и сопредельных территорий. Л.: Изд ЛГУ, 1983. Т. 2. 504 с.
- Зимин В. Б., Сазонов С. В., Лапшин Н. В., Хохлова Т. Ю., и др. Орнитофауна Карелии. Петрозаводск: Изд. КНЦ РАН, 1993. 220 с.
- Носков Г. А., Смирнов О. П. Территориальное поведение и миграции большой синицы (Parus m. major L.)//Экология птиц Приладожья. Труды БИНИИ ЛГУ. 1981. Вып. 32. С. 100-130.
- Смирнов О. П., Тюрин В. М. К биологии размножения большой синицы в Ленинградской области. Орнитология. М.: Изд. МГУ, 1981. Вып. 16. С. 185-188.
- Balen J. H. van, Noordwijk A. J. van, Visser J. Liefetime reproductive success and recruitment in two Great Tit populations. Ardea. 1987. V. 75. P. 1-11.
- McCleery R. H., Clobert J. Differences in recruitment of young by immigrant and resident Great Tts in Wytham Wood. Population biology of passerine birds. NATO ASI Series. V. G 21. Berlin: Springer, 1990. P. 423-439.
- Лихачев Г. Н. Материалы по биологии птиц, гнездящихся в искусственных гнездовьях. Тр. Приокско-Террасного гос. заповедника. 1961. Вып. 4. С. 82-146.
- Slagsvold T. Annual and geographical variation in the time of breeding in the Great Tit Pamus major and the Pied Flycatcher Ficedula hypoleuca in relation to environmental phenology and spring temperature. Ornis Scandinavica. 1976. V. 7. P. 127-145.
- Sanz J. J. Effects of geographic location and habitat on breeding parameters of Great Tits. Auk. 1998. V. 115. P. 1034-1051.
- Johansson H. Kullstorlek och hackningsframgang hos visssa holkhackande smafaglar i centrala Sverige 1972-1974 (1952-1963). Fauna och flora. 1974. V. 69. 212-218.
- Haartman L. von. The Nesting Habits of Finnish birds 1. Passeriformes. Commentationcs Biologicae Soc. Sci. Fenn. 1969. V. 32. l-187.
- Шульц Г. Э. Общая фенология. Л.: Наука, 1981. С. 188 с.
- Gwinner E. Circannual clocks in avian reproduction and migration. Ibis. 1996. V. 138. P. 47-63.
- Dawson A., King V. M., Bentley G. E., Ball G. Photoperiodic Control of Seasonality in Birds. J. of Biological rhythms. 2001. V. 16. P. 365-380.
- Silverin B. Reproductive Adaptations to Breeding in the North. American Zoologist. 1995. V. 35. P. 191-202.
- Perrins C. M. Eggs, egg formation and the timing of breeding. Ibis. 1996. V. 138. P. 2-15.
- Зимин В. Б. Экология воробьиных птиц северо-запада СССР. Л.: Наука, 1988. 184 с.
- Nager R. G., Noordwijk A. J. van. Proximate and ultimate aspects of phenotypic plasticity in timing of great tit breeding in a heterogeneous environment. American Naturalist. 1995. V. 146. P. 454-474.
- Balen J. H. van. A comparative study of the breeding ecology of the great tit (Parus major) in different habitats. Ardea. 1973. V. 61. P. 1-93.
- Meijer T., Nienaber U., Lancer U., Trillmich F. Temperature and timing of egg-laying of european starlings. Condor 1999. V. 101. P. 124-132.
- Nager R. On the effects of small scale variation in temperature and food availability on laying date and egg size in Great Tits (Parus major). Population biology of passerine birds. NATO ASI Series. V. G 21. Berlin: Springer, 1990. P. 187-197.
- Winkel W., Hudde H. Long-term trends in reproductive traits of tits (Parus major, P. caeruleus) and Pied Flycatcher (Ficedula hypoleuca). J. of Avian Biol. 1997. V. 28. P. 187-190.
- Slagsvold T. Breeding time of birds in relation to latitude. Norw. J. Zool. 1975. V. 23. P. 213-218.
- Slagsvold T. Hypotheses on breeding time and clutch-size in birds. Norw. J. Zool. 1975. V. 23. P. 219-222.
- Veistola S., Lehikoinen E., Iso-Iivari L. Breeding biology of the Great Tit Parus major in a marginal population in Northernmost Finland. Ardea. 1995. V. 83. P. 419-420.
- Crick H. Q. P., Dudley C., Glue D. E., Thomson D. L. UK birds are laying eggs earlier. Nature. 1997. V. 388. 526 p.
- McCleery R. H., Perrins C. M. Temperature and egg-laying trends. Nature. 1998. V. 391. P. 30-31.
- Назарова Л. Е., Филатов Н. Н. Изменчивость климата по данным метеорологических наблюдений//Климат Карелии: изменчивость и влияние на водные объекты и водосборы. Петрозаводск: КарНЦ РАН, 2004. С. 12-34.
- Visser M. E., Adriaensen F., Balen J. H. van, Blondel J. et. al. Variable responses to large-climate change in European Parus populations. 2003. Proc.of the Royal Soc. of London Ser. B. P. 02PB0699.1-02PB0699.5.
- Lack D. Population studies of birds. Oxford. Clarendon Press. 1966. 341 p.
- Blondel, J., Clamens A., Cramm P., Gaubert H., Isenmann P. Population studies on tits in the Mediterranean region//Ardea. 1987. V. 75. P. 21-34.
- Cichon M., Linden M. The timing of breeding and offspring size in Great Tits Parus major. Ibis. 1995. V. 137. P. 364-370.
- Perrins C. M., McCleery R. H. The effect of age and pair bond on the breeding success of Great Tit Parus major. Ibis. 1985. V. 127. P. 306-315.
- Glutz von Blotzheim U. N., Bauer K. M. Muscicapidae -Paridae. Handbuch der Vogel Mitteleuropas. Bd.13. Teil 1. Wiesbaden, Aula-Verlag, 1993. 808 s.
- Klomp H. Fluctuations and stability in Great Tit populations. Ardea 1980. V. 68. 205-224.
- Dhondt A. A., Kast T. L., Allen P. E. Geographical differences in seasonal clutch size variation in multi-brooded bird species. Ibis. 2002. V. 144. P. 646-651.
- Beldal E. J., Barba E., Gil-Delgado J. A., Iglesias D. J., Lorez G. M., Monros J. S. Laying date and clutch size of Great Tit (Parus major) in the Mediterranean region: a comparison of four habitat types. J. fur Ornithologie. 1998. V. 139. P. 269-276.
- Orell M., Ojanen M. Effect of habitat, date of laying and density on clutch size of the Great Tit Parus major in northern Finland. Holarctic Ecology. 1983.V. 6. P. 413-423.
- Haartman L. von. Population dynamics. Farner D. S., King J. R. (eds). Avian biology. V. 1. London: Academic Press, 1971. P. 391-459.
- Perrins C. M., Moss D. Reproductive rates in the Great Tit. J. Animal Ecology. 1975. V. 44. P. 695-706.
- Pettifor R. A., Perrins C. M., McCleery R. H. Individual optimization of clutch size in Great Tits. Nature. 1988. V. 336. P. 160-162.
- Godfray H. C. J., Partridge L., Harvey EH. Clutch sice. Annual Review of Ecology and Systematics. 1991. V. 22. P. 409-429.
- Семенов-Тян-Шанский О. И., Гилязов А. С. Птицы Лапландии. М.: Наука, 1991. 288 с.
- Бианки В. В., Шутова Е. В. К экологии большой синицы в Мурманской области. Бюлл. МОИП. Отд. биологии. Т. 83. Вып. 2. 1978. С. 63-70.
- Нумеров А. Д. Популяционная экология большой синицы в Окском заповеднике. Орнитология. М.: Изд. МГУ. 1987. Вып. 22. С. 3-21.
- Яремченко О. А., Болотников А. М. Биология размножения большой синицы. Орнитология. М.: Изд. МГУ. 1988. Вып. 23. С. 81-93.
- Добрынина И. Н. Предварительные результаты кольцевания больших синиц (Parus major) в северо-западных районах СССР. Результаты кольцевания и мечения птиц 1985 г. М.: Наука, 1991. 68-75.
- Резвый С. П., Носков Г. А., Гагинская А. Р. и др. Атлас миграций птиц Ленинградской области по данным кольцевания//Тр. С-Пб общества естествоиспыт. С-Пб., 1995. Т. 85. Вып. 4. 232 с.
- Викторов Г. А. Проблемы динамики численности насекомых на примере вредной черепашки. М.: Наука, 1967. 270 с.
- Воронцов А. И. Лесная энтомология. М.: Высшая школа, 1975. 368 с.
- Tinbergen J. M. Costs of reproduction in the Great Tit: intraseasonal costs associated with brood size. Ardea. 1987. V. 75. P. 111-122.
- Езерскас Л. И. О биологии птиц-дуплогнездников семейств синицевых и мухоловковых в Литовской ССР. Экология и миграции птиц Прибалтики (Тр. 4-й Прибалт. орнитол. конф.). Рига: Изд-во АН Латв. ССР, 1961. С. 115-122.
- Smith H. G., Kallander H., Nilsson J-A. Effects of experimentally altered brood size on frequency and timing of second clutches in the Great Tit. Auk. 1987. V. 104. P. 700-706.
