Популяционная структура соболя (Martes zibellina Linnaeus, 1758) Среднего Приамурья: кластерный анализ субпопуляций право- и левобережья реки Амур
 Среднего Приамурья: кластерный анализ субпопуляций право- и левобережья реки Амур")
Автор: Брыкова А.Л., Родимцева Д.В., Фрисман Л.В.
Журнал: Региональные проблемы @regionalnye-problemy
Рубрика: Математическое моделирование. Биология
Статья в выпуске: 4 т.26, 2023 года.
Бесплатный доступ
Проведенный ранее мониторинг аллельного состава субпопуляций соболя Приамурья в охотничьи сезоны 2011-2021 гг. показал присутствие аллелей, маркирующих различия соболиного населения левобережья и правобережья р. Амур. При попарном сравнении сезонных сборов субпопуляций лево- и правобережья был обнаружен разброс уровня генетической дифференциации от практически нулевых значений до промежуточного уровня. Стало понятно, что для определения количества различающихся генетических групп соболя в данном регионе необходимо привлечение дополнительных математических методов работы с полученными данными. Целью настоящей работы является определение и визуализация наиболее вероятного числа различающихся генетических групп соболей рассматриваемой территории. Были изучены три субпопуляции соболя Буреинского нагорья (левобережье р. Амур) и для сравнения рассмотрена одна субпопуляция c западных склонов Сихотэ-Алиня (правобережье р. Амур). Анализ данных по двум микросателлитным локусам Ma3 и Mer041 на материале охотничьих зимних сезонов с 2011 г. по 2022 г. проведен с помощью программ Structure 2.3.4 и Structure Harvester. По результатам, полученным с помощью этих программ, можно предположить, что на рассматриваемой территории данный вид представлен двумя генетическими группами (кластерами) - «левобережной» и «правобережной». Структура сохраняется как для материала сборов рассматриваемых зимних охотничьих сезонов 2012-2013 гг. и 2013-2014 гг., так и для суммарных сборов 2011-2022 гг. Формирование такой структуры, несомненно, - результат наложения истории становления ареала вида и генетической специфики различающихся антропогенно обусловленных интродукций из двух географических районов, проводимых здесь в середине прошлого века.
Соболь, кластерный анализ, генетические группы, микросателлитные локусы, субпопуляции, буреинское нагорье
Короткий адрес: https://sciup.org/143181149
IDR: 143181149 | УДК: 575.2:599.742.4 | DOI: 10.31433/2618-9593-2023-26-4-24-32
Genetic structure of the sable (Martes zibellina Linnaeus, 1758) at the Amur region: cluster analysis of subpopulations
The Amur region sable sub-populations monitoring of the allelic composition in the hunting seasons of 2011-2021 showed the presence of alleles marking the differences between the sable population at the Amur River left and right banks. A pairwise comparison of seasonal collections of left- and right-bank sub-populations revealed a spread in the level of genetic differentiation from almost zero values to intermediate ones. It became clear that in order to determine the number of different genetic groups of sable in this region, we need additional mathematical methods to work with the obtained data. The purpose of this work is to determine and visualize the most likely number of different genetic groups of sables in the territory under consideration. It was studied three sub-populations of the Bureinsky Highland sable (left bank of the Amur River). For comparison, it was taken a species sub-population from the Sikhote-Alin western slopes (the Amur River right bank). The analysis of data for two microsatellite loci Ma3 and Mer041, on the material of hunting winter seasons from 2011 to 2022, was carried out using the programs Structure 2.3.4 and Structure Harvester. Based on the results obtained with these programs, it can be assumed that in the territory under consideration, this species is represented by two genetic groups (clusters) - “the left-bank” and “the right-bank”. The structure remains the same for both the collection material of the 2012-2013 winter hunting seasons and those of 2013-2014, as well as for the total collections of 2011-2022. The formation of such a structure is undoubtedly the result of both the historical formation of the species habitat and the genetic specifics of anthropogenic introductions from two different geographical areas.
Текст научной статьи Популяционная структура соболя (Martes zibellina Linnaeus, 1758) Среднего Приамурья: кластерный анализ субпопуляций право- и левобережья реки Амур
Институт комплексного анализа региональных проблем ДВО РАН, ул. Шолом-Алейхема 4, г. Биробиджан, 679016, e-mail: , ; e-mail: , ; e-mail: ,
российской территории. Централизованный запрет на охоту и массовые интродукции зверьков из двух географических районов – окрестностей оз. Байкал и севера Буреинского хребта – восстановили численность. Возникла необходимость описания структуры вновь сформированных по- пуляций в различных частях ареала. Развитие методов молекулярно-генетического анализа и обработки полученных данных стимулировало работу. В последние десятилетия в качестве генетических маркеров популяционной структуры активно и успешно привлекаются микросателлиты – адаптивно нейтральные и кодоминантно наследуемые гены ядерной ДНК [2, 10, 11].
Уже первые исследования маркеров митохондриальной ДНК соболя привлекли внимание к обитателям правобережья Амура, а именно хребта Сихотэ-Алинь. Сравнение пространственно разобщенных популяций территории России показало, что уровень аллельного разнообразия популяции Сихотэ-Алиня значительно превышает подобный показатель в исследованных популяциях Сибири, Камчатки и Урала. Для ряда локусов (среди них Ма3 и Mer041) были обнаружены аллели, встречающиеся только у соболей Сихотэ-Алиня [3].
Сравнение аллельного разнообразия трех микросателлитных локусов (Ма2, Ма3 и Mer041) в субпопуляциях соболей с Буреинского нагорья и западной части Сихотэ-Алиня по сборам одного охотничьего сезона 2012–2013 гг. позволило обнаружить более высокий уровень аллельного разнообразия в субпопуляции правобережья (Сихотэ-Алинь) по сравнению с левобережными [5]. Для локусов Ма3 и Mer041 были обнаружены аллели, маркирующие субпопуляции по обе стороны р. Амур. Однако частота встречаемости таких аллелей оказалась низкой. Полученные значения попарных FST между пространственно разобщенными группами обитателей северной, центральной частей и южных отрогов Буреинского хребта не противоречили нулевой гипотезе отсутствия их дифференциации, что позволило рассматривать эти географические субпопуляции в рамках единой популяции. Однако попарное сравнение субпопуляции соболей Сихотэ-Алиня с субпопуляциями Буреинского хребта показало, что результат неоднозначен. Определялись как нулевой, так и слабый, но статистически достоверный уровни дифференциации.
Наиболее перспективные для исследования локусы Ма3 и Mer041, характеризующиеся присутствием уникальных аллелей в субпопуляциях лево- либо правобережья Амура в сезон 2012–2013 гг., были включены нами в мониторинговое исследование, проведенное на материале, собранном в охотничьи сезоны c 2011–2012 гг. по 2020–2021 гг. [9]. Мониторинг аллельного состава в трех субпопуляциях соболя Буреинского хребта и одной субпопуляции соболя Сихотэ-Алиня по- казал присутствие четырех аллелей, маркирующих различия соболиного населения левобережья и правобережья р. Амур. Однако эти аллели, как правило, относятся к категории редких и встретились в выборках только одного либо двух сезонов. Оценки наблюдаемой и ожидаемой гетерозиготности, соответствия распределению Харди-Вайнберга и F-статистик, проведенные с помощью программы Arlequin, подтвердили обнаруженную в сезон 2012–2013 гг. относительную однородность популяционной структуры соболя Буреинского хребта в рассматриваемый отрезок времени. Однако при попарном сравнении сезонных сборов географических субпопуляций лево- и правобережья Амура обнаружен разброс уровня генетической дифференциации от практически нулевых значений до промежуточного уровня. Стало понятно, что для определения количества различающихся генетических групп соболя в данном регионе необходимо привлечение дополнительных математических методов работы с полученными данными.
Цель настоящего исследования – определить и визуализировать наиболее вероятное число различающихся генетических групп у соболей рассматриваемой территории.
Задача – по данным аллельного разнообразия локусов Ma3 и Mer041 провести оценку популяционной структуры соболя Буреинского нагорья и западной части Сихотэ-Алиня с помощью программ Structure 2.3.4 [7] и Structure Harvester [6].
Рассматривается суммарный материал 2011–2022 гг. и отдельно материал сезонов 2012– 2013 гг. и 2013–2014 гг., наиболее полно представленных в нашем исследовании.
Материалы и методы
В работе использован биологический материал от 633 соболей из четырех географических районов (северной, центральной и южной частей Буреинского хребта и западной части Сихотэ-Алиня), охватывающий охотничьи сборы 2011– 2022 гг. Ни одно животное не было отловлено специально для данной работы. Материал предоставлен охотниками, имеющими лицензии на отлов соболей.
Исследуемые субпопуляции левобережья р. Амур, локализованные на Буреинском нагорье, условно обозначены нами как «Маган» (охотничьи сборы в северной части Буреинского хребта в верховьях р. Бурея), «Сутырь» (охотничьи сборы в средней части Буреинского хребта у притоков среднего течения р. Бурея) и «Каменушка» (охотничьи сборы в бассейне р. Бира, в южной части
Буреинского хребта и северной части хребта Малый Хинган). Субпопуляция правобережья Амура обозначена как «Сихотэ-Алинь» (охотничьи сборы около рек Манома и Хар). Ниже представлено географическое происхождение (рис. 1), посезонное и численное распределение собранного материала (табл.). Подробно этапы работы с материалом и методы выявления микросателлитных локусов представлены в [9].
Оценка популяционной структуры соболя проведена в программе Structure 2.3.4 [8], в которой используются методы Монте-Карло, в ее основе лежит схема марковской цепи (Markov Chain Monte Carlo, MCMC). Это дает возможность минимизировать неравновесное состояние соответствия распределению Харди-Вайнберга в кластерном анализе особей [12]. Данная программа также применялась для визуализации результатов и их математического подтверждения методами G. Evanno [7]. Особенность ввода исходных дан- ных в Structure 2.3.4 дает возможность расширить объем выборки за счет того, что можно включать особей, по которым был обработан всего один из двух рассматриваемых маркеров (данные недостающего локуса указываются как «Missing data -1»).
Для предварительного выбора стартовой точки марковской цепи («burnin») проводили 500 000 итераций с последующим построением марковской цепи с 1 000 000 итераций для гипотетического числа различающихся генетических групп (К) от 1 до 6 в пятикратной повторности для каждой величины К.
Для интерпретации результатов использовано максимальное значение ΔK, полученное с помощью веб-программы Structure Harvester [6], в которой для определения наиболее вероятного числа генетических групп или, как их называют авторы программы, «кластеров» используется метод G. Evanno [7].
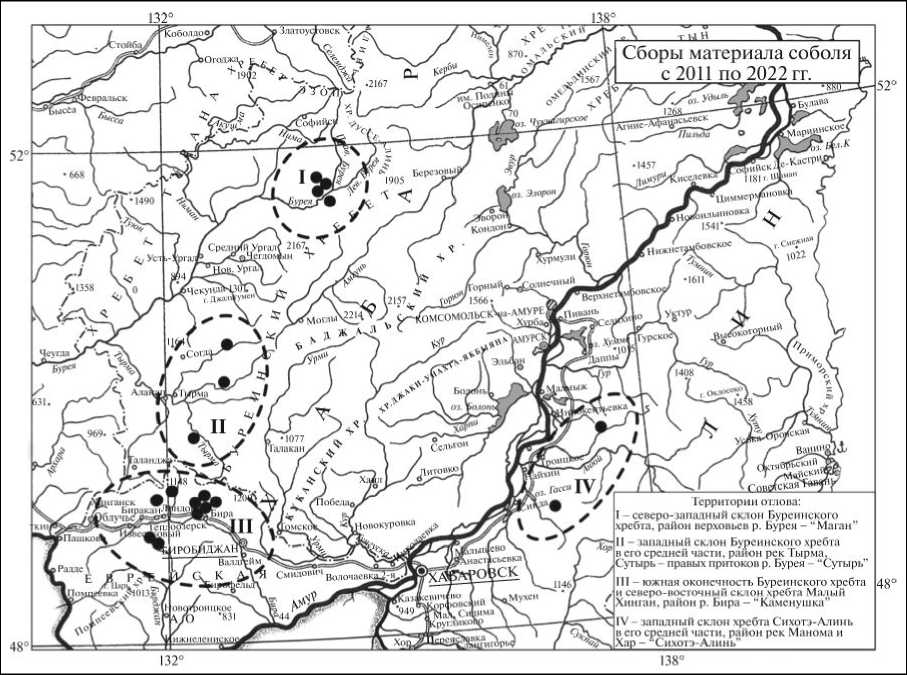
Рис. 1. Географическая локализация районов сбора материала соболя на территории Среднего Приамурья
-
Fig. 1. Geographical localization of the areas of collecting sable material in the Middle Amur Region
Таблица
Географическая локализация и количество исследуемого материала соболя по сезонам
Table
Geographical localization and quantity of the studied sable material by season
|
Субпопуляции |
Общее число животных |
Число животных в отдельные охотничьи сезоны |
||||||||
|
2011– 2012 |
2012– 2013 |
2013– 2014 |
2014– 2015 |
2015– 2016 |
2017– 2018 |
2019– 2020 |
2020– 2021 |
2021– 2022 |
||
|
Левобережье р. Амур |
||||||||||
|
I «Маган» |
150 |
– |
14 |
21 |
52 |
– |
– |
43 |
20 |
– |
|
II «Сутырь» |
187 |
– |
52 |
30 |
35 |
33 |
– |
– |
– |
37 |
|
III «Каменушка» |
173 |
19 |
15 |
54 |
– |
34 |
– |
– |
31 |
20 |
|
Правобережье р. Амур |
||||||||||
|
IV «Сихотэ-Алинь» |
123 |
– |
30 |
28 |
– |
16 |
23 |
26 |
– |
– |
|
Суммарно |
633 |
19 |
111 |
133 |
87 |
83 |
23 |
69 |
51 |
57 |
Результаты
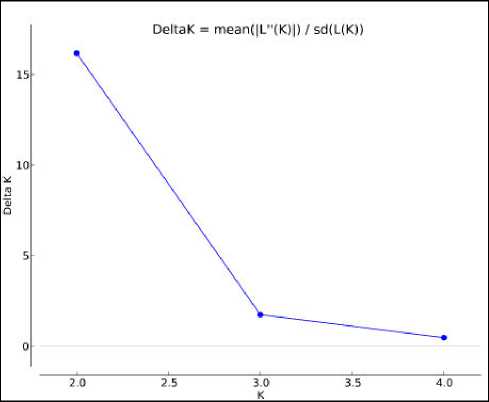
Моделирование популяционной структуры в программе Structure 2.3.4 определило возможное число генетических групп в выборке. Согласно данным, полученным с помощью Structure Harvester, их наиболее вероятное число в выборках сезонов 2012–2013 гг., 2013–2014 гг. и суммарной выборке 2012–2022 гг. одинаково и равно двум, так как для каждой из них наибольшее значение параметра ΔK установлено при К=2. На рис. 2 в качестве примера показан график значений ΔK для суммарной выборки 2011–2022 гг. (рис. 2). Выявленные характеристики принадлежности к группам представлены во всех рассматриваемых субпопуляциях, но с различающимися коэффициентами принадлежности.
Сезон 2012–2013 гг. Для материала этого сезона во вкладке «Столбчатая диаграмма» программы Structure 2.3.4 при К=2 получено следующее изображение распределения генотипов в рассматриваемых субпопуляционных выборках (рис. 3). Каждая особь на этой диаграмме представлена вертикальной линией, разделенной на K цветовых сегментов, где длина каждого пропорциональна расчетному коэффициенту принадлежности к группе.
Полученная столбчатая диаграмма отображает наибольшую генетическую близость двух из трех буреинских субпопуляций, центральной – «Сутырь» и южной – «Каменушка». Несколько отличается от них субпопуляция северной части Буреинского хребта – «Маган». Наибольшим образом выделяется субпопуляция «Сихотэ-Алинь».
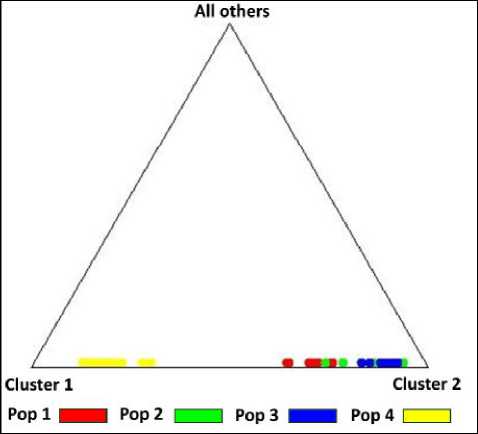
По треугольной диаграмме программы Structure 2.3.4 (рис. 4) можно визуально оценить, как субпопуляции распределяются на два кластера.
Из треугольной диаграммы (рис. 4) видно, что субпопуляции левобережья р. Амур объединяются в один кластер. Субпопуляция «Сихотэ-
Рис. 2. График значений ΔK для суммарной выборки 2011–2022 гг., максимальное значение при К=2
-
Fig. 2. Graph of ΔK values for the total sample of 2011–2022, the maximum value at K = 2
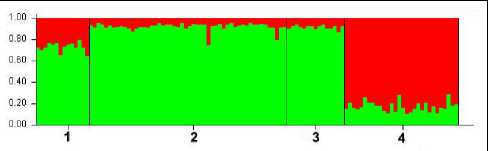
Рис. 3. Столбчатая диаграмма распределения генотипов субпопуляций соболя охотничьего сезона 2012–2013 гг. при К=2; по вертикали – доля частот аллелей соответствующей генетической группы (апостериорная вероятность), по горизонтали – обозначения субпопуляций: 1 – «Маган», 2 – «Сутырь», 3 – «Каменушка», 4 – «Сихотэ-Алинь»
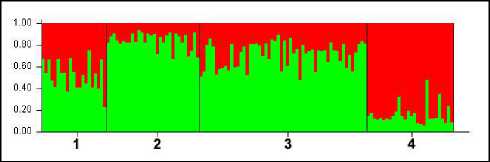
Рис. 5. Столбчатая диаграмма распределения генотипов субпопуляций соболя охотничьего сезона 2013–2014 при К=2; по вертикали – доля частот аллелей соответствующей генетической группы (апостериорная вероятность), по горизонтали – обозначения популяций: 1 – «Маган», 2 – «Сутырь», 3 – «Каменушка», 4 – «Сихотэ-Алинь»
-
Fig. 3. Bar chart of genotype distribution of the 2012–2013 sable sub-populations hunting season at K=2; vertically – the proportion of allele frequencies of the corresponding genetic group (a posteriori probability), horizontally – the designations of sub-populations: 1 – «Magan», 2 – «Sutyr», 3 – «Kamenushka»,4 – «Sikhote-Alin»
Fig. 5. Bar chart of sable sub-populations genotype distribution for the 2013–2014 hunting season at K=2; vertically – the proportion of allele frequencies of the corresponding genetic group (a posteriori probability), horizontally – population designations: 1 – «Magan», 2 – «Sutyr», 3 – «Kamenushka», 4 – «Sikhote- Alin»
Р ис. 4. Треугольная диаграмма распределения субпопуляций в выборке соболей охотничьего сезона 2012–2013 гг.
1 – «Маган», 2 – «Сутырь», 3 – «Каменушка», 4 – «Сихотэ-Алинь»
-
Fig. 4. Triangular diagram of sub-populations distribution in the sample of sables for the 2012– 2013 hunting season.
1 – «Magan», 2 – «Sutyr», 3 – «Kamenushka», 4 – «Sikhote-Alin»
Алинь» в этот сезон представляет отдельный кластер, не пересекающийся с левобережным.
Сезон 2013–2014 гг. Для материала этого сезона столбчатая диаграмма (Structure 2.3.4) при К=2 выглядит следующим образом (рис. 5).
Судя по ней, субпопуляции Буреинского хребта относятся к одной генетической группе, в которой субпопуляции «Сутырь» и «Каменушка» так же, как и в сезон 2012–2013 гг., наиболее близки друг к другу. Отдельно представлена субпопуляция «Сихотэ-Алинь». Треугольная диаграмма программы Structure 2.3.4 (рис. 6) для сезона 2013–2014 гг. показывает значительное взаимное наложение состава левобережных субпопуляций. В этом сезоне наблюдается пересечение сихотэ-алинской группы (правобережного кластера) с северной буреинской субпопуляцией «Маган».
Суммарный материал сборов 2011–2022 гг. Для суммарной выборки при К=2 получено следующее изображение (рис. 7). Судя по столбчатой диаграмме, картина, полученная при анализе материала по девяти охотничьим сезонам с 2011 г. по 2022 г., аналогична с той, что представлена в двух рассмотренных выше сезонах: единая генетическая группа субпопуляций Буреинского хребта и отдельно – субпопуляция Сихотэ-Алиня.
По треугольной диаграмме (Structure 2.3.4) (рис. 8) можно визуально оценить, как субпопуля-
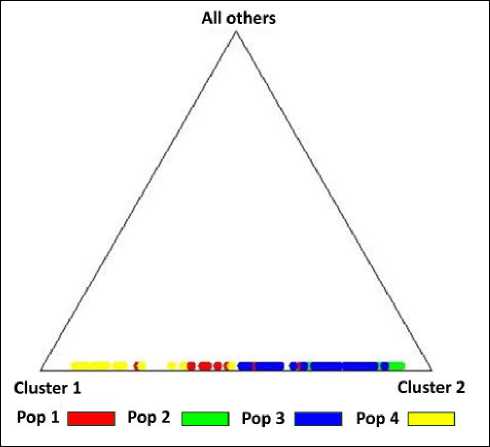
Рис. 6. Треугольная диаграмма распределения субпопуляций в выборке соболей охотничьего сезона 2013–2014 гг.
1 – «Маган», 2 – «Сутырь», 3 – «Каменушка», 4 – «Сихотэ-Алинь»
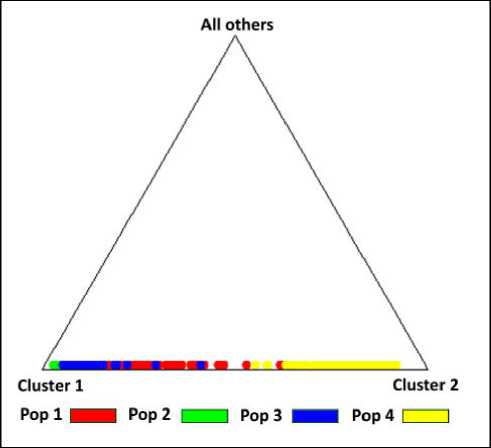
Рис. 8. Треугольная диаграмма распределения субпопуляций в суммарной выборке соболей 2011–2022 гг.
1 – «Маган», 2 – «Сутырь», 3 – «Каменушка», 4 – «Сихотэ-Алинь»
Fig. 6. Triangular diagram of sub-populations distribution in the sample of sables for the 2013–2014 hunting season.
-
1 – «Magan», 2 – «Sutyr», 3 – «Kamenushka», 4 – «Sikhote-Alin»
ции, представленные суммарными сборами, разделяются на два кластера.
По данным суммарных выборок субпопуляции Буреинского хребта также объединены в одну генетическую группу (кластер). Привлекает внимание частичное наложение субпопуляций севера Буреинского хребта «Маган» с сихотэ-алинской субпопуляцией (правобережным кластером).
Fig. 8. Triangular diagram of sub-populations distribution in the total sample of sables 2011–2022.
-
1 – «Magan», 2 – «Sutyr», 3 – «Kamenushka», 4 – «Sikhote-Alin»
Обсуждение
Результат кластерного анализа четырех субпопуляций, полученный с помощью алгоритма Structure, показывает существование на рассматриваемой территории двух кластеров – левобережного и правобережного по отношению к реке Амур. Формирование такой структуры, несомненно, результат наложения истории становле-
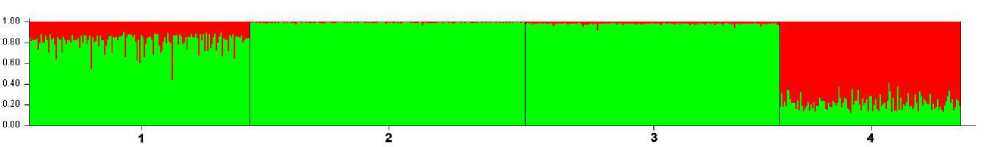
Рис. 7. Столбчатая диаграмма распределения генотипов субпопуляций соболя по суммарным сборам 2011–2022 гг., при К=2; по вертикали – доля частот аллелей соответствующей генетической группы (апостериорная вероятность), по горизонтали – обозначения субпопуляций: 1 – «Маган», 2 – «Сутырь», 3 – «Каменушка», 4 – «Сихотэ-Алинь»
Fig. 7. Bar chart of sable sub-populations genotypes distribution by total collections of 2011–2022, at K=2; vertically – the proportion of allele frequencies of the corresponding genetic group (a posteriori probability), horizontally – the designations of subpopulations: 1 – «Magan», 2 – «Sutyr», 3 – «Kamenushka», 4 – «Sikhote-Alin»
ния ареала вида на данной территории и генетической специфики различающихся антропогенно обусловленных интродукций, проводимых здесь в середине прошлого века.
По современным представлениям северный Сихотэ-Алинь, особенно его восточные склоны, с сохранявшимися здесь в ледниковые периоды плейстоцена лесными рефугиумами [4] – это территория, где соболь мог пережить плейстоценовое оледенение и по его окончанию расселиться по Северной Евразии [1]. Ближайшими районами такого расселения являлись леса левобережья Амура, в том числе Буреинского нагорья. Это означает, что автохтонные популяции левобережья и правобережья Амура исходно должны быть близки при несколько меньшем аллельном разнообразии на левобережье, обусловленном эффектом основателя. После антропогенно организованного падения численности начала XX века очаги автохтонных соболей сохранились на Сихотэ-Алине лишь в недоступных человеку горных участках. Сохранился автохтонный соболь и на севере Буреинского хребта, откуда зверьки были расселены в различные регионы России, в том числе и в южную часть Буреинского нагорья. Таким образом, современные субпопуляции Буреинского хребта являются потомками северной автохтонной формы этой территории, с несколько меньшим аллельным разнообразием за счет эффекта основателя. Историческая общность происхождения и единый источник интродукции объясняет их объединение в единую группу.
В то же время генетическая структура си-хотэ-алинской субпопуляции, выделяемая с помощью алгоритма программы Structure в отдельную группу, потенциально должна отличаться от буреинской, так как может иметь в своем составе характеристики трех его составляющих: автохтонной сихотэ-алинской и двух реинтродуцирован-ных – северо-буреинской и иркутской. Именно в этой генетической группе выявлено наибольшее аллельное разнообразие среди четырех представленных субпопуляций [5]. Изменяющаяся структура сихотэ-алинской генетической группы может быть обусловлена существованием на Сихотэ-Алине пространственно неоднородной популяционной структуры, сформировавшейся из двух интродуцированных и автохтонной форм. Данная неоднородность проявляется в динамике аллельного разнообразия, в разбросе генетических дистанций как при сравнении сихотэ-алинских сезонных сборов, так и сезонных сборов лево- и правобережья [9], что отражено в структуре столбчатых и треугольных сезонных диаграмм.
Немаловажным фактором, изменяющим популяционную структуру соболя, может быть антропогенное воздействие, которое, судя по полученным данным мониторинга 2011–2022 гг., не изменяет значимо генетическую структуру буре-инских субпопуляций. Для выделения фактора, определяющего изменение популяционной структуры в генетической группе Сихотэ-Алиня, необходимо расширение географии собираемого материала. Дальнейший мониторинг уже известных субпопуляций, поиск наиболее информативных микросателлитных маркеров и подбор программ для обработки полученных данных позволят дополнить исследование популяционной структуры соболя Приамурья, оценить и понять причины ее изменения в пространстве и во времени.
Исследование выполнено при поддержке гранта департамента образования Еврейской автономной области (ЕАО) в соответствии с распоряжением губернатора ЕАО Р.Э. Гольдштейна от 10.05.2023 № 104-рг.
Список литературы Популяционная структура соболя (Martes zibellina Linnaeus, 1758) Среднего Приамурья: кластерный анализ субпопуляций право- и левобережья реки Амур
- Абрамов К.Г. Соболь в охотничьем хозяйстве Дальнего Востока. М.: Наука, 1967. 116 с.
- Каштанов С.Н., Свищева Г.Р., Лазебный О.Е., Колобков Д.С., Пищулина С.Л., Мещерский С.Л., Рожнов В.В. Влияние антропогенных факторов на генетическое разнообразие вида соболь (Martes zibellina L.) // Молекулярная биология. 2015. Т. 49, № 3. С. 449-454. DOI: 10.7868/S002689841503009X.
- Каштанов С.Н., Свищева Г.Р., Пищулина С.Л., Лазебный О.Е., Мещерский И.Г., Симакин Л.В., Рожнов В.В. Географическая структура генофонда соболя (Martes zibellina L.): данные анализа микросателлитных локусов // Генетика. 2015. Т. 51, № 1. С. 78-88. DOI: 10.7868/ S001667581501004X.
- Крестов П.В., Баркалов В.Ю., Омелько А.М., Якубов В.В., Накамура Ю., Сато К. Реликтовые комплексы растительности современных рефугиумов северо-восточной Азии // Ко-маровские чтения. Владивосток: Дальнаука, 2009. Вып. 56. С. 5-63.
- Фрисман Л.В., Шлюфман К.В., Брыкова А.Л. Генетическая структура популяции соболя (Martes zibellina L.) Буреинского нагорья по сборам одного охотничьего сезона // Региональные проблемы. 2019. Т. 22, № 4. С. 46-55. DOI: 10.31433/2618-9593-2019-22-4-46-55.
- Earl Dent A., VonHoldt B.M. STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method // Conservation Genetics Resources. 2012. Vol. 4. P. 359-361. DOI: 10.1007/s12686-011-9548-7.
- Evanno G., Regnaut S., Goudet J. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study // Molecular ecology. 2005. Vol. 14, N 8. P. 2611-2620. DOI: 10.1111/j.1365-294X.2005.02553.x.
- Falush D., Stephens M., Pritchard J.K. Inference of population structure using multilocus genotype data: linked loci and correlated allele frequencies // Genetics. 2003. Vol. 164, N 4. P. 1567-1587. PMID: 12930761.
- Frisman L.V., Brykova A.L. Monitoring of the Genetic Structure of the Sable Population (Martes zibellina Linnaeus, 1758) in the Middle Amur Region // Russian Journal of Genetics. 2023. Vol. 59, N 4. P. 375-384. DOI: 10.1134/ S1022795423040038.
- Modorov M., Monakhov V., Mikryukov V., Erokhin N., Tkachenko I., Polezhaeva M., Ranyuk M. Microsatellite multiplex assay for sable (Martes zibellina) and pine marten (Martes martes) // Mammal Research. 2020. Vol. 65. P. 855-862. DOI: 10.1007/s13364-020-00529-4.
- Ranyuk M., Modorov M., Monakhov V., Devyat-kin G. Genetic differentiation of autochthonous sable populations in Western and Eastern Siberia // Journal of Zoological Systematics and Evolutionary Research. 2021. Vol. 59. P. 2539-2552. DOI: 10.1111/jzs. 12565.
- Smulders M.J.M., Van Der Schoot J., Arens P., Vosman B. Trinucleotide repeat microsatellite markers for black poplar (Populus nigra L.) // Molecular Ecology Notes. 2001. Vol. 1, N 3. P. 188190. DOI: 10.1046/j .1471-8278.2001.00071.x.
