Популяционная структура Thalictrum minus L. на Южном Урале

Автор: Самойлова Л.Ю., Федоров Н.И., Михайленко О.И., Хасбутдинова И.Р.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биологические ресурсы: флора
Статья в выпуске: 1-4 т.13, 2011 года.
Бесплатный доступ
Методом кластерного анализа по средневыборочным значениям морфологических параметров растений проанализирована популяционная структура Thalictrum minus L. на Южном Урале. Выявлено 6 групп локальных популяций, четыре из которых представлены 3 подвидами этого вида - T. minus subsp. minus, T. minus subsp. macrophyllum и T. minus subsp. flexuosum, а 2 - гибридными популяциями этих подвидов.
Василистник малый, популяционная структура, южный урал
Короткий адрес: https://sciup.org/148199762
IDR: 148199762 | УДК: [582.675:581.55]:470.55/.58
Population structure of Thalictrum minus L. in South Urals
By method of cluster analysis using the average selected values of morphological parameters of plants it was analysed the population structure of Thalictrum minus L. in South Urals. 6 groups of local populations were revealed, four of which are presented by 3 subspecies of this kind - T. minus subsp. minus, T. minus subsp. macrophyllum and T. minus subsp. flexuosum, and 2 - by hybrid populations of these subspecies.
Текст научной статьи Популяционная структура Thalictrum minus L. на Южном Урале
Вид Thalictrum minus L. (сем. Ranuncu-laceae) является одним из перспективных источников изохинолиновых алкалоидов, обладающих болеутоляющей, противовоспалительной, гипотензивной и противоопухолевой активностью, на основе которых уже получены или находятся в стадии разработки высокоэффективные медицинские препараты [1]. Вид достаточно широко распространен в Южноуральском регионе в остепненных разнотравно-злаковых лугах союза Trifolion montani , в степных сообществах союза Festucion valesia-cae , в остепненных дубняках союза Lathyro-Quercion, мезофитных кострово-злаковых лугах союза Festucion pratensis и ряде других растительных сообществ [2]. Вариабельность состава и содержания алкалоидов в растениях может зависеть от внутривидовой структуры [3].
Цель работы: анализ популяционной структуры T. minus на Южном Урале.
Материалы и методы. Материал для исследования собран на 19 пробных площадях в частично изолированных локальных популяциях. 10 пробных площадей заложены в Пред-уралье: 3 в северной части в зоне распространения хвойных и хвойно-широколиственных лесов («Аскинская» ( АС ), «Караидельская» (КА), «Салаватская» (СА)), одна – в Меся-гутовской лесостепи («Дуванская» (ДУ)), 3 – в зоне распространения широколиственных лесов
Самойлова Лилия Юлаевна, аспирантка
Хасбутдинова Ильнара Рамзиловна, студентка
(«Кушнаренковская» (КУ), «Уфимская» (УФ), «Бирская» (БИ)), 3 – в степных районах («Туй-мазинская» (ТУ), «Кугарчинская» (КГ), «Зиан-чуринская» (ЗИ)) и одна – на Белебеевской возвышенности в лесостепном районе («Давле-кановская» (ДА)). В горной части на Южном Урале заложены пробные площади – «Бурзян-ские» (БУ1, БУ2, БУ3), «Белорецкие» (БЕ1, БЕ2) и «Зилаирская» (ЗЛ). 2 пробные площади заложены в Зауралье: «Баймакская» (БА) и «Учалинская» (УЧ). На пробных площадях выполняли геоботанические описания и отбирали случайным образом, но не ближе, чем в 20 м друг от друга, 20-25 генеративных растений. У растений учитывали 23 вариабельных параметра стебля, листа, соцветия и цветка, используемых в систематике таксономических единиц рода Thalictrum. Для анализа популяционной структуры Т. minus на Южном Урале проводили кластерный анализ 19 локальных популяций по средневыборочным значениям морфологических параметров растений. Кластерный анализ проводили по программе SYN-TAX IV [4].
Результаты и обсуждение. Результаты кластерного анализа различия южно-уральских локальных популяций T. minus по средневыборочным значениям морфологических параметров приведены на рис. 1. На дендрограмме анализируемые локальные популяции объединились в 2 крупных кластера, каждый из которых, в свою очередь, включает по 3 группы локальных популяций. Популяции первого кластера (группы I-III) приурочены к зонам распространения горных хвойных и смешанных лесов, а также смешанных широколиственно-темнохвойных и сосново-березовых лесов северной части РБ. Во второй кластер вошли популяции, приуроченные, преимущественно, к степным и лесостепных районам
Предуралья, Зауралья и Зилаирского плато, лесная растительность в которых представлена широколиственными лесами (группы IV-VI). Растения популяций, вошедших в первый кластер, отличаются от растений популяций второго кластера большей длиной и шириной листочков сложного листа, длиной плодоножек и числом орешков в соплодии, а также пропорционально большими междоузлиями соцветия (отношение числа паракладиев к длине соцветия) (табл. 1). Таким образом, на различия локальных популяций по морфологическим параметрам растений влияет комплекс экологических факторов, обусловливающих особенности горно-лесных, лесостепных и степных местообитаний, а не географическая удаленность популяций друг от друга.
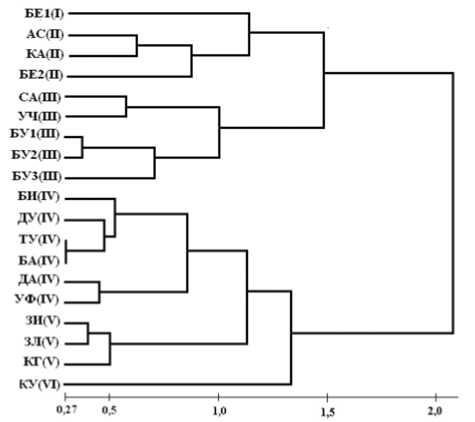
Рис. 1. Дендрограмма различий локальных популяций Thalictrum minus L. по средневыборочным значениям морфологических параметров растений
Таблица 1. Различие групп локальных популяций Thalictrum minus L., выделенных при проведении кластерного анализа
|
Морфологические параметры растений |
Кластеры |
|||||
|
I |
II |
III |
IV |
V |
VI |
|
|
длина побега, см |
85±4 |
122±3 |
97±2 |
78±2 |
73±2 |
67±2 |
|
доля растений с прямыми стеблями, % |
22,7 |
61,7 |
44,3 |
21,7 |
2,7 |
0,0 |
|
доля растений с изогнутыми стеблями, % |
77,3 |
38,3 |
55,7 |
78,3 |
97,3 |
100,0 |
|
длина листочка среднего листа, мм |
25,1±1,1 |
23,1±0,8 |
20,4±0,5 |
13,7±0,2 |
14,8±0,4 |
11,7±0,3 |
|
ширина листочка среднего листа, мм |
18,3±0,8 |
16,8±0,7 |
15,4±0,4 |
10,1±0,2 |
10,8±0,3 |
11,2±0,4 |
|
отношение ширины листочка среднего листа к его длине |
0,74±0,03 |
0,73±0,02 |
0,76±0,01 |
0,74±0,01 |
0,73±0,02 |
0,96±0,03 |
|
доля растений с кожистыми листочками, % |
0,0 |
6,7 |
78,3 |
99,3 |
97,3 |
100,0 |
|
доля растений с тонкими некожистыми листочками, %, |
100,0 |
93,3 |
21,7 |
0,7 |
2,7 |
0,0 |
|
доля растений с остротреугольными лопастями, % |
100,0 |
48,3 |
76,4 |
95,1 |
86,3 |
100,0 |
|
доля растений с короткояйцевид-ными тупыми лопастями, % |
0,0 |
51,7 |
23,6 |
4,9 |
13,7 |
0,0 |
|
доля растений с отклоненными листьями, % |
81,8 |
93,3 |
91,5 |
78,3 |
2,7 |
100,0 |
|
доля растений с прижатыми листьями, % |
18,2 |
6,7 |
8,5 |
21,7 |
97,3 |
0,0 |
|
длина соцветия, см |
20±3 |
31±2 |
29±2 |
21±1 |
17±1 |
11±1 |
|
число паракладиев соцветия, шт. |
5,3±0,5 |
8,9±0,4 |
8,7±0,4 |
8,6±0,3 |
8,2±0,5 |
6,6±0,3 |
|
отношение числа паракладиев к длине соцветия |
0,32±0,03 |
0,32±0,02 |
0,36±0,02 |
0,48±0,02 |
0,51±0,02 |
0,65±0,03 |
|
средняя длина плодоножек, мм |
10,3±1,1 |
4,9±0,2 |
4,8±0,2 |
3,4±0,1 |
3,8±0,2 |
3,6±0,2 |
|
максимальная длина плодоножек, мм |
26,1±2,7 |
12,7±0,6 |
12,5±0,6 |
6,2±0,2 |
6,7±0,2 |
4,7±0,2 |
|
среднее число орешков, шт. |
4,6±0,3 |
4,4±0,1 |
4,5±0,1 |
3,4±0,1 |
4,2±0,1 |
3,8±0,1 |
|
максимальное число орешков, шт. |
10,7±0,7 |
7,2±0,2 |
7,9±0,2 |
5,4±0,1 |
5,9±0,2 |
4,8±0,1 |
Первая группа первого кластера представлена одной популяцией (БЕ1), приуроченной к сосново-березовым лесам центральной части Южного Урала. Растения этой группы отличаются от растений всех остальных групп (в том числе групп II и III) более крупными тонкими листочками, большей длиной плодоножек и большим числом орешков в соплодии, а также меньшим числом паракладиев в соцветии. По этим морфологическим особенностям растения популяции БЕ1, можно отнести к подвиду T. minus subsp. macrophyllum (V. Boczantzeva) Krupkina [5, 6]. Вторая группа первого кластера объединяет локальные популяции, приуроченные к зоне распространения темнохвойно-широколиственных лесов на севере Республики Башкортостан (АС и КА) и к елово-пихтово-сосновым горным лесам Южного Урала (БЕ2). Растения этой группы популяций морфологически близки к растениям первой группы – у них тоже крупные, тонкие некожистые листочки у листьев, что позволяет их также отнести к подвиду subsp. Macro-phyllum. От первой группы эти популяции отличаются более крупными размерами побегов у растений, что, возможно, связано с большей влажностью почвы в характерных для них местообитаниях. Кроме того, они отличаются меньшей длиной плодоножек (более характерной для других подвидов T. minus (Krup-kina) [5]). В третью группу входят 5 локальных популяций: из южной части горно-лесной зоны, расположенные на границе с горной лесостепью (БУ1, БУ2, БУ3), из лесостепного района восточного макросклона Южного Урала (УЧ) и из западной части северо-восточной лесостепи Башкирского Предуралья (СА). Эта группа отличается от второй, прежде всего, преобладанием растений с кожистыми листочками, что не характерно для подвида subsp. macrophyllum и может быть следствием гибридизации с другими подвидами T. minus.
Во втором кластере наиболее сильно отличается локальная популяция КУ (группа VI), приуроченная к остепненным лугам Пред-бельского лесостепного района. Растения этой популяции отличаются небольшими высотой и соцветием и имеют отклоненные от стебля листья с мелкими, округлыми листочками с большим числом лопастей. По этим признакам они соответствуют описанию T. minus s. str. (T. minus subsp. minus). Растения локальных популяций, объединившиеся на дендрограмме в пятую группу, соответствуют форме T. minus subsp. flexuosum Krupkina. Для растений этого подвида характерны прижатые к стеблю листья с мелкими листочками и сближенные орешки [5], однако, по мнению других авторов [7], орешки могут быть как сближенными, так и растопыренными. Популяции 5 группы приурочены к остепненным лугам и степным сообществам степных районов Предуралья (КГ и ЗИ) и Зилаирского плато (ЗЛ). В «Определителе сосудистых растений Оренбургской области» [8] и в последней редакции «Флоры Средней полосы Европейской части России» [6] этот подвид описан как самостоятельный вид T. flexuosum Bernh. ex Reichenb. В последнюю, четвертую группу второго кластера объединились 6 локальных популяций, описанных в лесных и лесостепных районах средней части Башкирского Предуралья (БИ, ДА, ТУ, УФ), на северо-востоке Башкирского Пред-уралья в Месягутовской лесостепи (ДУ) и степных районах Зауралья (БА). В этих локальных популяциях преобладают растения с отклоненными листьями, но также встречаются и с прижатыми.
Выводы: проведенный кластерный аннализ различия южно-уральских локальных популяций T. minus по средневыборочным значениям морфологических признаков позволяет сделать вывод о том, что T. minus представлен на Южном Урале 3 различающимися по экологии подвидами: subsp. minus, subsp. flexuosum и subsp. macrophyllum. Подвид subsp. macrophyllum (группы локальных популяций I, II) распространен в возвышенной части Южного Урала и в северной части Башкирского Предуралья в подзоне южной тайги. Во «Флоре Восточной Европы» [5] для этого подвида указывается распространение в лесах, и пойменных лугах и в зарослях кустарников по берегам водоемов на Кавказе, в Западной и Восточной Сибири, северо-восточной части Средней Азии, Скандинавии (Финляндия) и Арктике, но не указывается на Южном Урале. Однако во «Флоре СССР» [9] была описана лесная уральская и восточно-сибирская раса T. minus – Т. globiflorum Ldb., которая является синонимом подвида subsp. macrophyllum [10]. На границе с лесостепными подзонами subsp. macrophyllum гибридизируется с другими подвидами, вследствии чего образуется лесостепная форма этого вида с высокой представ-леностью растений с кожистыми листочками (группа III). Подвид subsp. flexuosum (группа V) распространен на остепненных, часто каменистых склонах с выходами коренных пород южных cтепных районов Предуралья и Зи-лаирского плато. Этот подвид также ДОСтаточно широко распространен южнее, в Оренбургской области [8]. Подвид subsp. minus (группа VI) выявлен только в центральной части Башкирского Предуралья. В подавляющем большинстве предуральских и зауральских локальных популяций этого подвида встречаются также растения с прижатыми листьями, что свидетельствует о возможной гибридизации подвида subsp. minus с подвидом subsp. flexu-osum, видимо, имевшего большее распространение в плейстоцене, когда на территории Предуралья и Южного Урала имели большое распространение лесостепные сообщества.
Список литературы Популяционная структура Thalictrum minus L. на Южном Урале
- Юнусов, М.С. Химия изохинолиновых алкалоидов: Учеб. пособие. -Уфа: Изд-во УГНТУ, 2007. 53 с.
- Жигунова, С.Н. Продуктивность надземной части Thalictrum minus L. в лесных и послелесных луговых сообществах/С.Н. Жигунова, О.И. Михайленко, Л.Ю. Самойлова//Аграрная Россия. 2009. Специальный выпуск. С. 178-179.
- Федоров, Н.И. Род Delphinium L. на Южном Урале: экология, популяционная структура и биохимические особенности. Монография. -Уфа: Гилем, 2003. 149 с.
- Podani, J. SYN-TAX IV: Computer programs for data analysis in ecology and systematics on IBM-PC and Macintosh Computers. -Trieste, 1990. 145 p.
- Цвелев, Н.Н. Василисник -Thalictrum L./Флора Восточной Европы. Т. Х. -СПб.: Мир и семья, 2001. С. 186-195.
- Маевский, П.Ф. Флора Средней полосы Европейской части России. -М.: Товарищество научных изданий КМК. 2006. С. 249-250.
- Бочанцева, В.В. Thalictrum minus L. в Европейской части СССР//Ботанический журнал. 1973. Т. 58, № 11. С. 1641-1644.
- Рябинина, З.Н. Флора сосудистых растений Оренбургской области/З.Н. Рябинина, М.С. Князев. -М.: Товарищество научных изданий КМК, 2009. С. 304-305.
- Невский, С.А. Василистник -Thalictrum L./Флора СССР. Т. 7. -М.: Изд-во АН СССР, 1937. C. 510-524.
- Фризен, Н.В. Василистник -Thalictrum L.//Флора Сибири. Portulacaceae -Ranunculaceae. -Ново-сибирск, 1993. Т. 6. С. 198-206.
