Популяционно-генетическая характеристика, оценка геномного инбридинга и гомозиготности крупного рогатого скота черно - пестрой и голштинской пород по STR и SNP маркерам в России

Автор: Недашковский И.С., Сермягин А.А., Костюнина О.В., Волкова В.В., Гладырь Е.А., Янчуков И.Н.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Генетика
Статья в выпуске: 4, 2021 года.
Бесплатный доступ
Объект исследования - быки-производители черно-пестрой и голштинской пород, имеющие STR профили и прошедшие процедуру генотипирования по SNP -маркерам. Предмет исследования - уровень геномного инбридинга и гомозиготности, а также популяционно-генетическая характеристика на их основе. С возрастанием Fx , основанного на данных родословного учета, отмечено увеличение уровня геномного инбридинга ( FROH ), рассчитанного по SNP -маркерам. Результаты по 9 STR -маркерам фиксируют волнообразное увеличение гомозиготности от I до IV группы с последующим возрастанием, начиная с V и до VII группы включительно. В ходе исследования средних значений FROH в соответствии с годами рождения быков было установлено достоверное отличие двух последних групп (2009-2011 и 2012-2014 гг.) от всех остальных при попарном сравнении. Результаты расчета FROH и Ca9 быков из разных стран происхождения указывают на статистически значимую дифференциацию. Отмечено достоверное отличие наибольшего по выборкам уровня FROH в голштинской черно-пестрой породе от красно-пестрой голштинской и черно-пестрой пород. Значения Fst между породной принадлежностью животных по SNP и STR имели незначительную дифференциацию (0.008-0.027). Различия в расчете по STR между черно-пестрой голштинской и красно-пестрой голштинской породами оказались незначительны (менее 20%), в то время как остальные величины разнились многократно. Fst между животными РФ и Германии, РФ и Нидерланд по данным STR равен 0.006 и 0.008, по данным SNP 0.005 и 0.006, соответственно. Отмечено взаимное увеличение значения Fst и года рождения быка-производителя, что подчеркивает наибольшую удаленность популяции 1983-1997 гг. от популяций последних лет. Схожесть значений Fst между генеалогическими линиями сменяется значительным разбросом в показателях при попарном их сравнении.
Бык-производитель, голштинская порода, черно-пестрая порода, геномный инбридинг, микросателлиты, однонуклеотидные полиморфизмы, паттерны гомозиготности
Короткий адрес: https://sciup.org/147236788
IDR: 147236788 | УДК: 636.22/28.082.12:575.113.2 | DOI: 10.17072/1994-9952-2021-4-295-306
Population and genetic features, genomic inbriding and homozygosity level for black -and-white and Holstein breeds by STR and SNP markers in Russia
The object of the study was sires of the Black-and-White and Holstein breeds, which had STR profiles and passed the genotyping procedure for SNP markers. The subject of the study was the level of genomic inbreeding and homozygosity, as well as population-genetic characteristics based on them. With an increase in Fx based on pedigree data, an increase in the level of genomic inbreeding (FROH) calculated from SNP markers was also noted. The results for 9 STR markers record a wavelike increase in homozygosity from group I to group IV with a subsequent increase from group V to group VII inclusive. During the study of the average FROH values in accordance with the years of birth of sires, a significant difference was found between the last two groups (2009-2011; 2012-2014) from all the others in a pairwise comparison. The results of calculating the FROH and Ca9 of bulls from different countries of origin indicate a statistically significant differentiation of the group of animals from Russia (RF) from animals, born in German and Canadian. A significant difference was noted for the highest FROH level in the samples in the Holstein Black-and-White breed from the Red-and-White Holstein and Black-and-White breeds. The Fst values between the SNP and STR species of the animals had insignificant differentiation (0.008-0.027). Differences in STR calculation between the Black-and-White Holstein and Red- and-White Holstein breeds were insignificant, less than 20%, while the rest of the values differed many times. Fst between RF and Germany, RF and the Netherlands according to STR data is 0.006 and 0.008, according to SNP data 0.005 and 0.006, respectively. A mutual increase in the value of Fst and the year of birth of the sires was noted, emphasizing the greatest remoteness of the population of 1983-1997 from the populations of recent years, when new genotypes of bulls were obtained. The similarity of Fst values between genealogical lines is replaced by a significant scatter in the indicators when they are compared in pairs.
Текст научной статьи Популяционно-генетическая характеристика, оценка геномного инбридинга и гомозиготности крупного рогатого скота черно - пестрой и голштинской пород по STR и SNP маркерам в России
Оценка генетической дифференциации популяций как на субпопуляционном, так и на индивидуальном уровнях остается актуальной в части подхода и методов расчета. Использование STR (short tandem repeats) и SNP (single nucleotide polymorphism) маркеров, а также их преимущества для этих целей по сравнению с данными по родословной животного, рассматривались нами ранее [Недашковский и др., 2019, 2021].
Заинтересованность в исследованиях по этому направлению просматривается в последних работах отечественных и зарубежных ученых, направленных на поиск наиболее точного, с научной точки зрения, а главное – оптимального с коммерческой, подхода к генетическому контролю популяции и анализу генома для ведения направленной селекционной работы и моделированию желаемых свойств пробанда [Kim, Cole, Huson, 2013; Marrаs et al., 2015; Сермягин и др., 2020].
Точность оценки уровня накопления гомозиготности является базисом для проведения селекционного процесса как в конкретном хозяйстве (стаде), так и на популяционном уровне в целом [Кузнецов, 2000; Кузнецов, Валохина, 2010]. Основной анализ методов дивергентности популяций сводится к сопоставлению результатов исследований в части единства понимания терминологии и интерпретации полученных результатов [Заид и др., 2008; Кузнецов, 2014]. Использование генотипирования по SNP -маркерам, безусловно, является наиболее точным методом контроля гомозиготности и отражает суть селекционных процессов внутри популяции, но уступает по ценовому критерию STR (ценообразованию). Так, сопоставление результатов микросателлитного анализа по дистанциям между аллелями, генотипами и по различиям в размерах аллелей с иными программами (Arlequin v.3.5, GenePop v.4.7.3, RST22), давали схожие AMOVA -оценки. Стандартизация оценок Fst (subpopulation fixation index) и Фрт по Хендрику [Hedrick, 2005] имела оценки 0.35‒0.37, сопоставимые с результатами, полученными методами Нея-Хедрика [Me-irmans, Hedrick, 2011] (0.364‒0.375), Джоста [Jost, 2008] (0.292) и Морисита-Хорна [Chao et. al., 2016] (0.308). Проекции генетических дистанций GD (genetic distance) в пространстве главных координат (PCoA) на 2D плоскости были схожи [Кузнецов, 2021].
Геномная селекция, эра которой наступила во всем мире, позволяет вести направленную работу по желаемому признаку и проводить генетическую оценку племенной ценности ( G EBV ) животного, не дожидаясь результатов классической оценки по потомству. В среднем оценка быка-производителя по потомству, включающая основные хозяйственно-полезные качества, возможна спустя 3‒4 года, когда половозрелая особь сможет произвести уже свое потомство и начать лактацию. Этот срок является недопустимой “роскошью” в период современного интенсивного ведения животноводства и поддержания потребностей растущего населения в мире с достаточным производством продуктов питания и продовольственной безопасностью, если мы говорим о Российской Федерации [Доклады …, 2021; Об утверждении …].
В России на сегодняшний день исследований подобного рода по сравнительной оценке STR- и SNP -маркеров для изучения популяционно-генетической структуры скота основных молочных пород, не проводилось.
Цель нашей работы – сравнительное изучение уровня накопления инбридинга и оценка гомозиготности, рассчитанной по STR- и SNP- маркерам, между собой, в популяции крупного рогатого скота России, отнесенного к черно-пестрому корню, на основе комплексной характеристики популяционногенетических особенностей животных.
Материал и методы исследования
Объектом исследований являлись быки-производители следующих пород: черно-пестрой (BWRUS, 7 гол.), голштинской красно-пестрой масти (RedHL, 13 гол.), голштинской черно-пестрой масти (BWHL, 333 гол.), линий: Монтвик Чифтейн 95679 (MCHF), Пабст Говернер 882933 (PG), Рефлекшн Соверинг 198998 (RFS), Силинг Трайджун Рокит 252803 (SLTR), Вис Бэк Айдиал 1013415 (VBA), страна происхождения: Канада (CAN), Дания (DEN), Германия (DEU), Нидерланды (NED), Россия (RUS), принадлежащие племенным организациям Московской обл. За контроль принималась информация по величине инбридинга, полученного на основе родословной животных, по не менее четырем рядам предков. Показатели гомозиготности и геномного инбридинга служили группами сравнения.
Выделение тотальной (геномной) ДНК проводили из спермы быков с помощью колонок Nexttec (Nexttec Biotechnologie GmbH, Германия) в соответствии с рекомендациями производителя. В качестве ДНК-маркеров использовалась мультиплексная панель из 12 микросателлитных локусов: TGLA227 , BM2113 , TGLA53 , ETH10 , SPS115 , TGLA122 , INRA23 , TGLA126 , BM1818 , ETH3 , ETH225 , BM1824 , рекомендованных FAO (Food and agriculture organization) и ISAG (International Society for Animal Genetics), для проведения популяционно-генетических исследований и определения чистопородности крупного рогатого скота, в частности. Продукты амплификации для их последующей детекции подвергались анализу на капиллярном генетическом анализаторе ABI 3130xl Genetic Analyzer («Applied Biosystems», «Life technologies», США). Информация о длине аллелей по исходным данным получена в программе Gene Mapper v.4 («Applied Biosystems», «Life technologies», США). Статистическую обработку данных осуществляли с помощью программы GenAlEx 6.50 [Peakall, Smouse, 2012]. Непосредственный расчет индивидуальной гомозиготности проводили как отношение количества гомозиготных локусов к общему количеству анализируемых локусов.
Для полногеномного генотипирования по SNP , концентрацию ДНК определяли с помощью флуориметра Qubit 3.0 (Life Technologies, США). Оценка чистоты нуклеиновых кислот проводилась на приборе NanoDrop 2000. SNP генотипирование осуществляли с помощью чипа средней плотности Illumina Bovine 50К Beadchip («Illumina Incorporated», США), включающего в себя 54 609 SNP- маркеров. Для получения надежных результатов был проведен контроль качества SNP с использованием программы Plink 1.9 [Chang et al., 2015]: отбирались только маркеры, локализованные на аутосомах, генотипированные не менее чем у 90% особей, и с частотой минорных аллелей не менее 3%. После проведения контроля качества было отобрано 42 797 SNP . Размер паттернов гомозиготности (ROH) у быков-производителей определяли с помощью пакета cgaTOH [Zhang et al., 2013].
ROH определяли при условии, если 15 или более последовательных гомозиготных SNP присутствовали на исследуемом участке генома при плотности не менее 1 SNP на каждые 100 кb с промежутками между ними не более 1000 кb. Дифференциацию ROH проводил согласно длине в Mb в связи с давностью возникновения демографического события в популяции молочного скота на группы: [1;2], (2;4], (4;8], (8;16] и >16 Mb [Ferenčakovic et al., 2013].
Статистически F ROH (inbreeding coefficient determined using ROH) определяется как длина аутосомного генома, присутствующего в ROH , деленная на общую длину аутосомного генома, охватываемого пулом SNP [Leutenegger et al., 2003]. Для каждого быка были рассчитаны F ROH > 1 Mb, F ROH > 2 Mb, F ROH > 4 Mb, F ROH > 8 Mb и F ROH > 16 Mb на основе ROH различной минимальной длины (>1, >2, >4, >8 или >16 Mb). F ROH определяли для разных минимальных длин ROH , потому что длина аутозиготных сегментов в геноме, по прогнозам, будет показывать экспоненциальное распределение, со средней длиной, равной 100/2 g Моргана, где g – число поколений со времени общего предка. Для корректной оценки ROH на 50 К панели учитывали ROH более 4 м.п.н. (миллион пар нуклеотид) [Purfield et. al., 2012]. К выбору такого подхода учета ROH , т.е. сканирования генома рамкой, учитывающей последовательности более 4 м.п.н., склоняются и отечественные учёные, проводившие исследования на популяции молочного скота Ленинградской обл. [Смарагдов, Кудинов, 2019].
Пространственную визуализацию осуществляли на основе метода многомерного шкалирования (Plink
-
1. 9) для групп быков-производителей в соответствии с: породой, годом рождения (генерационным периодом), генеалогической (линейной) принадлежностью и страной происхождения (выведения) животных. Регрессионный анализ и генетические корреляции проводили в Statistica v.10. Весь массив данных ран-
- жирован на группы, основным критерием формирования которых являлась возможность наиболее информативной визуализации изменения показателя геномного инбридинга и гомозиготности по мере увеличения коэффициента инбридинга. Начиная от аутбредных особей в I группе, последующий шаг в 1%-ном Fx продолжается до V группы, т.е. предельно-рекомендованных значений показателя инбридинга при линейном разведении. Последующие VI и VII группы обусловлены включением оставшихся животных в выборку особей, теоретически подверженных инбредной депрессии.
Результаты и их обсуждение
Анализ данных табл. 1 показывает, что с возрастанием коэффициента инбридинга, основанного на родословной, от 0% в первой до 7.03–12.50% в VII группе, параллельно увеличивается уровень геномного инбридинга, рассчитанного на информации по SNP -маркерам (0.061–0.12). Так, группы F ROH I, II и III имеют достоверные (р˂0.001) отличия от групп V, VI и VII. Коэффициент регрессии по F ROH (R=0.009) отражает не только основную положительную направленность (зависимость) тренда, но и является достоверно значимым. Информация по микросателлитным локусам демонстрирует схожие закономерности в случае с использованием 12 маркеров STR , где третья группа ( Ca =21.7%) имеет достоверные различия (р˂0.05; р˂0.01) от V, VI и VII групп с уровнем гомозиготности от 25.8 до 35.0%. Коэффициент регрессии в данном расчете также положителен (R=0.014) и достоверно значим. Результаты по 9 STR фиксируют волнообразное увеличение гомозиготности от I до IV группы ( Fx =0.00–1.97%) с 30.5 до 29.2% для Ca9 , с последующим возрастанием, начиная с V и до VII группы включительно ( Fx =7.03–12.5%) с 30.4 до 36.7% для Ca9 .
Таблица 1
Оценка уровня инбридинга по родословной и геномным данным, гомозиготности для популяции быков-производителей черно-пестрой и голштинской пород
[Assessment of the level of inbreeding by pedigree and genomic data, homozygosity for the population of bulls-producers of black-and-white and Holstein breeds]
|
Коэффициент инбридинга (Fx) |
n |
Fx |
F ROH |
Ca12 |
Ca9 |
|
I) 0.00 |
92 |
0.000±0.000 |
0.061±0.003 |
0.234±0.012 |
0.305±0.014 |
|
II) 0,00-0.98 |
212 |
0.306±0.022 |
0.066±0.002 |
0.224±0.008 |
0.300±0.010 |
|
III) 0.20-0.98 |
120 |
0.054±0.023* |
0.069±0.002 |
0.217±0.011 |
0.296±0.014 |
|
IV) 1.03-1.97 |
54 |
1.600±0.035***/*** |
0.072±0.004* |
0.227±0.017 |
0.292±0.020 |
|
V) 2.05-3.91 |
61 |
2.963±0.080***/***/*** |
0.100±0.003***/***/*** |
0.258±0.015н.д/* |
0.304±0.018 |
|
VI) 4.01-6.69 |
16 |
4.769±0.221***/***/***/*** |
0.109±0.008***/***/*** |
0.281±0.027н.д/* |
0.326±0.031 |
|
VII) 7.03-12.50 |
10 |
***/***/***/***/*** 7.918±0.526 |
0.120±0.013***/***/*** |
0.350±0.043*/**/* |
0.367±0.055 |
|
R |
353 |
- |
0.009 |
0.014 |
0.006 |
R – коэффициент регрессии (regression coefficient); *р˂0.05; **р˂0.01; ***р˂0.001; н.д. - недостоверно;
Fx – индивидуальный коэффициент инбридинга, основанный на оценке по родословной;
Froh – уровень геномного инбридинга, геномный коэффициент инбридинга;
Ca – уровень гомозиготности по STR для 12 ( Ca12 ) и 9 ( Ca9 ) STR маркеров;
/ * – достоверное различие между показателями II и III, II и IV, II и V, II и VI, II и VII групп;
/ */ * – достоверное различие между показателями III и IV, III и V, III и VI, III и VII групп;
/ */ */ * – достоверное различие между показателями IV и V, IV и VI, IV и VII групп;
/ */ */ */ * – достоверное различие между показателями V и VI, V и VII групп.
Была оценена взаимосвязь переменных по средствам непараметрического коэффициента корреляции Спирмена (r s ). В данном случае, коэффициент ранговой корреляции отражает генетические зависимости (является генетическим коэффициентом корреляции), поскольку рассчитан на основе геномной информации. Отмечена прямая умеренная связь r s =0.443 (р˂0.05) между Fx и F ROH . Положительная слабая связь r s =0.013 (р˂0.05) между Fx и Са12 отражает основные зависимости. Связь между Fx и Cа9 положительная, но не является достоверно значимой. Длина гомозиготного сегмента (паттерна гомозиготности ROH) в геноме пробанда, показала умеренную положительную связь r s =0.44 (р˂0.05) с Fx , что подтверждает закономерности демографических событий, чем теснее инбридинг, тем длиннее фрагмент нуклеотидной последовательности нуклеиновых кислот.
Рассмотрение вопроса оценки уровня инбридинга и накопления гомозиготности с использованием 12 STR-маркеров, на наш взгляд, является, скорее, альтернативным, менее информативным и статистически неверным с научной точки зрения, поскольку существует возможность неправильной интерпретации полученных результатов. Неоднозначные результаты приравнивались к нолю, что могло отразиться негативным образом, в конечном итоге, как индивидуально на пробанде, так и на групповом уровне в целом, поставив под вопрос точность достоверности разницы между исследуемыми группами. По этой причине, дальнейшее исследование с проведением PCoA (principal coordinates analysis) анализа, для визуализации значений индивидуального индекса фиксации Fis (individual fixation index), в части информации по STR-маркерам, осуществлялось по 9 микросателлитным локусам STR.
В ходе исследования средних значений F ROH по годам рождения быков-производителей было установлено достоверное (p<0.001; p<0.01; р˂0.05) отличие двух последних групп (2009‒2011 гг.; 2012‒2014 гг.) от всех остальных при попарном сравнении (табл. 2). Это объясняется уровнем ведения селекционной работы по принципу разведения «в себе» с ограниченным числом улучшателей по признакам молочной продуктивности и воспроизводства. Результат расчета F ROH по стране происхождения указывает на достоверно (p<0.001) значимую дифференциацию группы животных из Российской Федерации от животных, рожденных в Германии и Канаде (немецкого и канадского корня).
Таблица 2 Распределение быков-производителей в соответствии с годом и страной происхождения, породной и генеалогической принадлежностью
[Distribution of breeding bulls according to year and country of origin, breed and genealogical affiliation] Группа I n I Froh I Ca12 I Ca9 I Fx
Год рождения быка-производителя
|
I) 1983-1997 |
20 |
0.064±0.008 |
0.254±0.034 |
0.316±0.040 |
2.048±0.643 |
|
II) 1999-2003 |
71 |
0.069±0.004 |
0.214±0.015 |
0.302±0.017 |
1.462±0.191 |
|
III) 2004-2008 |
135 |
0.069±0.003 |
0.228±0.010 |
0.311±0.013 |
1.271±0.142 |
|
IV) 2009-2011 |
72 |
0.087±0.004*/**/*** |
0.251±0.012 |
0.299±0.016 |
1.409±0.220 |
|
V) 2012-2014 |
55 |
0.092±0.004**/***/*** |
0.259±0.018 |
0.283±0.018 |
1.264±0.214 |
Страна
|
I) РФ |
51 |
0.059±0.005 |
0.199±0.017 |
0.346±0.019 |
1.189±0.270 |
|
II) Нидерланды |
26 |
0.061±0.005 |
0.218±0.019 |
0.316±0.032 |
0.766±0.204 |
|
III) Дания |
21 |
0.068±0.007 |
0.190±0.019 |
0.291±0.020* |
1.215±0.291 |
|
IV) Германия |
150 |
0.076±0.002*/** |
0.251±0.010**/н.д/* |
0.296±0.012* |
1.515±0.145н.д/* |
|
V) Канада |
105 |
0.089±0.003***/***/**/*** |
0.248±0.011*/н.д/** |
0.290±0.014* |
1.468±0.181н.д/* |
Генеалогическая линия
|
I) Силинг Трайд-жун Рокит 252803 |
4 |
0.057±0.019 |
0.250±0.034 |
0.389±0.071 |
1.122±0.997 |
|
II) Пабст Говернер 882933 |
8 |
0.064±0.008 |
0.198±0.027 |
0.333±0.020 |
0.415±0.203 |
|
III) Монтвик Чифтейн 95679 |
40 |
0.068±0.005 |
0.248±0.020 |
0.339±0.023 |
1.389±0.256н.д/** |
|
IV) Вис Бэк Айдиал 1013415 |
183 |
0.076±0.002 |
0.234±0.009 |
0.299±0.010 |
1.477±0.142н.д/*** |
|
V) Рефлекшн Соверинг 198998 |
118 |
0.080±0.003н.д/н.д/* |
0.238±0.011 |
0.290±0.013 |
1.303±0.147н.д/*** |
Порода
|
I) Черно-пестрая |
7 |
0.041±0.005 |
0.155±0.042 |
0.365±0.047 |
0.391±0.166 |
|
II) Краснопестрая голштинская |
13 |
0.055±0.006 |
0.243±0.017 |
0.385±0.030 |
1.388±0.577 |
|
III) Чернопестрая голштинская |
333 |
0.078±0.002***/*** |
0.238±0.007 |
0.298±0.008н.д/** |
1.401±0.097*** |
*р˂0.05; **р˂0.01; ***р˂0.001; н.д. – недостоверно;
/ * – достоверное различие между показателями II и III, II и IV, II и V, II и VI, II и VII групп;
/ */ * – достоверное различие между показателями III и IV, III и V, III и VI, III и VII групп;
/ */ */ * – достоверное различие между показателями IV и V, IV и VI, IV и VII групп;
/ */ */ */ * – достоверное различие между показателями V и VI, V и VII групп.
На наш взгляд, это связано с особенностями системы генетической оценки животных и ведением племенной работы в стране-происхождения. Тем не менее, российская популяция скота черно-пестрого корня, на которой в настоящий момент основывается голштинская порода отечественной селекции, несет в своем генотипе ценные отличительные свойства относительно импортированных животных. Невысокий уровень геномного инбридинга и небольшое количество животных в выборках по генеалогическим линиям Силинг Трайджун Рокит 252803, Пабст Говернер 882933 и Монтвик Чифтейн 95679 связаны с их «непопулярностью», особенно в последние годы для использования в программах разведения молочного скота России. Достоверное (p<0.001) отличие наибольшего по выборкам уровня FROH в голштинской черно-пестрой породе от красно-пестрой голштинской и черно-пестрой обусловлено интенсивным повсеместным использованием и тотальной голштинизацией, что также характерно для исследуемой популя- ции. Масштаб применения данной системы разведения наглядно демонстрируется максимальным (n=333) по выборкам количеством генотипированных быков-производителей.
Результаты STR- анализа по группам породной принадлежности и стране происхождения быков-производителей, в целом, отражают основные зависимости, отмеченные по средствам SNP -маркеров. Наблюдается достоверное (р˂0.05) различие группы животных из Российской Федерации от животных, рожденных в Дании, Германии и Канаде. Также группа животных красно-пестрой голштинской породы была достоверно (p<0.01) дифференцируема от группы черно-пестрой голштинской породы. Дивергенция между группами быков-производителей по линейной принадлежности и годам рождения, отмеченная в результатах анализа по SNP маркерам, оказалась наглядной, но статистически незначимой в случае с STR .
На рисунке 1 представлена сравнительная популяционно-генетическая характеристика пространственного положения групп животных черно-пестрой и голштинской пород разной масти. Так, на рис. 1 (справа) наблюдается кластеризация черно-пестрой и красно-пестрой голштинской породы от чернопестрой голштинской, которая имеет более размытые границы в случае расчетов по STR , представленным на рис. 1 (слева). Четкая дифференциация быков-производителей по годам рождения, наглядно отражена на рис. 2 (справа). Заметная обособленная локализация последних групп по годам рождения, а именно 2009–2011 и 2012–2014 гг., имеет логическое объяснение и заключается в ведении направленной селекционной работы и отображает генетический прогресс популяции. Визуализация значений STR на рис. 2 (слева) имела менее различимые границы по искомым группам. Анализ результатов линейной принадлежности, представленный на рис. 3, демонстрирует отсутствие как таковой «линейной принадлежности» к тому или иному участку пространственного положения групп, за исключением линии Пабста Говернера 882933 и Силинг Трайджун Рокита 252803 на рис. 3 (справа).
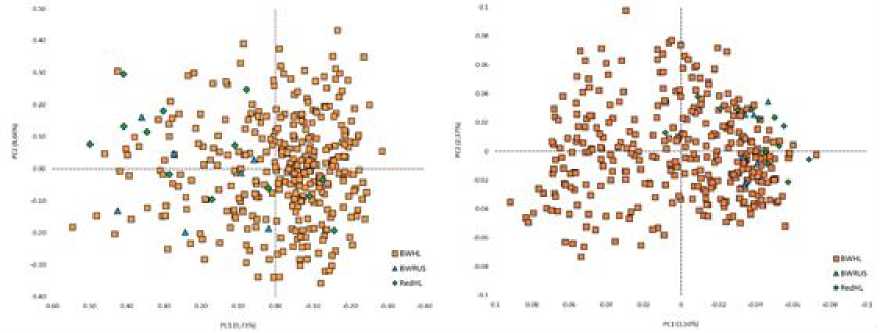
Рис. 1 . Сравнительная популяционно-генетическая характеристика пространственного положения групп животных черно-пестрой (BWRUS) и голштинской пород (BWHL, RedHL): на основе данных STR -маркеров (слева), на основе данных SNP- маркеров (справа)
[Comparative population-genetic characteristics of the spatial position of groups of animals of black-and-white (BWRUS) and Holstein breeds (BWHL, RedHL): based on STR marker data (left), based on SNP marker data (right)]
На рисунке 4, отображающем пространственное положение групп животных из разных стран происхождения, в обоих случаях расчетов, и по STR-, и по SNP- , наблюдается явное и определяемое различие в генетической структуре группы быков из Российской Федерации от зарубежных производителей, странами происхождения которых являются Канада и Германия.
Результаты анализа главных компонент, отражающие Fis по заданным критериям, представлены на рис. 1–4. Различия в структуре, наблюдаемые в пространстве двух главных координат, неслучайны, поскольку для визуализации пробанда по средствам STR , использовались данные по 9 микросателлитным локусам, содержащим по 2 аллеля (аллельных варианта) в каждом, т.е. 18 критериев для двух главных координат на плоскости. Для SNP , основанных на информации биочипов средней плотности (50 тыс. SNP ), количество значений составляет 81 в данном исследовании и может варьировать, превышая эти значения при использовании чипов большей плотности, таких как 150 тыс. SNP , 700 тыс. SNP и т.д.
Группа черно-пестрой породы характеризовалась наименьшим средним числом аллелей в расчете на 1 локус. Группа черно-пестрой голштинской породы занимала лидирующее положение по уровню генетического разнообразия (Ho = 0.727±0.029) и по среднему числу аллелей на локус (Na = 9.111±0.935), возможно потому, что представлена более многочисленной выборкой. Эффективное число аллелей (Ne) варьировало от 3.577±0.427 в черно-пестрой до 4.067±0.467 в черно-пестрой голштинской породах. При анализе индекса фиксации критических отклонений от равновесия по Харди-Вайнбергу не было отмече- но. Так, в группах красно-пестрой и черно-пестрой пород выявлено превышение числа гетерозигот над гомозиготами на 0.5 и 5.7% соответственно (Fis = –0.007…–0.084). Результат молекулярной дисперсии показал, что на долю внутригрупповой генетической изменчивости приходится 93% вариации, тогда как межгрупповые различия составляли 7% (p<0.001).
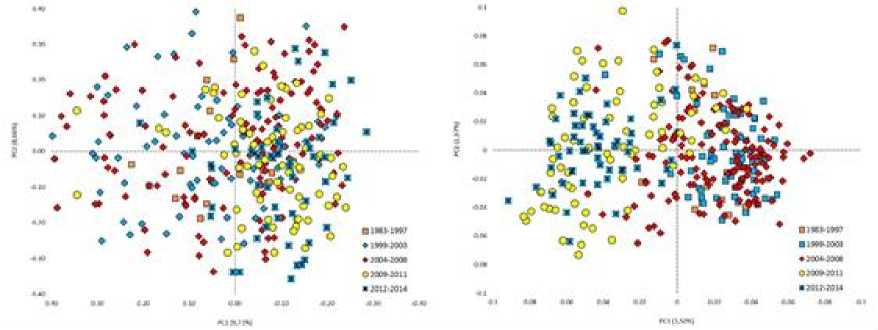
Рис. 2 . Сравнительная популяционно-генетическая характеристика пространственного положения групп животных в соответствии с годами рождения (генерационным интервалам): на основе данных STR- маркеров (слева), на основе данных SNP -маркеров (справа)
[Comparative population-genetic characteristics of the spatial position of animal groups in accordance with the years of birth (generation intervals): based on STR marker data (left), based on SNP marker data (right)]
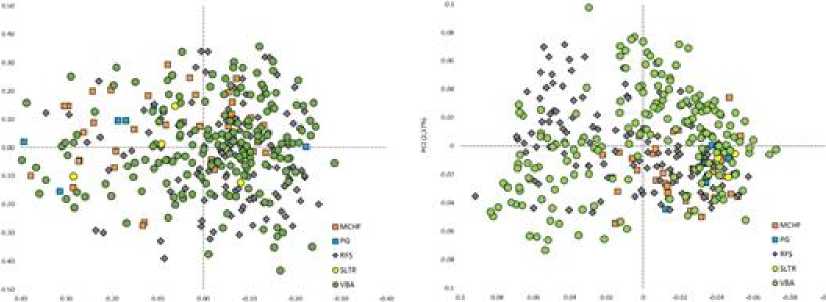
Рис. 3 . Сравнительная популяционно-генетическая характеристика пространственного положения групп животных разной генеалогической (линейной) принадлежности: на основе данных STR маркеров (слева), на основе данных SNP маркеров (справа): Монтвик Чифтейн 95679 (MCHF), Пабст Говернер 882933 (PG), Рефлекшн Соверинг 198998 (RFS), Силинг Трайджун Рокит 252803 (SLTR), Вис Бэк Айдиал 1013415 (VBA)
[Comparative population-genetic characteristics of the spatial position of groups of animals of different genealogical (linear) affiliation: based on STR markers (left), based on SNP markers (right): Montvik Chieftain 95679 (MCHF), Pabst Governor 882933 (PG), Reflection Sovering 198998 (RFS), Siling Trijun Rockit 252803 (SLTR), Vis Back Idial 1013415 (VBA)]
В таблицах 3–6 представлены попарные сравнения значений Fst , рассчитанные по STR- и SNP- маркерам, которые характеризуют степень дифференциации между исследуемыми субпопуляциями. D.L. Hart и A.G. Clark предложили классификацию, согласно которой генетическая дифференциация считается незначительной при Fst ≤ 0.05, умеренной при Fst = 0.05–0.15, и значительной при Fst = 0.15–0.25 [Hartl et al., 1997].
По этой классификации (табл. 3) все значения индекса фиксации имели незначительную дифференциацию (0.008–0.027), или слабую – по классификации Райта [Wright, 1978]. Различия в результатах расчета в части величины показателя Fst между черно-пестрой голштинской и красно-пестрой голштинской породами оказались незначительны, менее 20%, в то время как остальные величины разнились многократно.
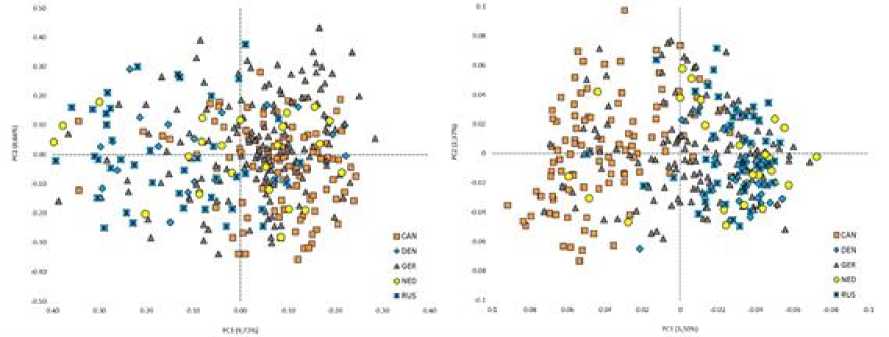
Рис. 4 . Сравнительная популяционно-генетическая характеристика пространственного положения групп животных из разных стран происхождения: на основе данных STR -маркеров (слева), на основе данных SNP -маркеров (справа): Канада (CAN), Дания (DEN), Германия (DEU), Нидерланды (NED), Россия (RUS)
[Comparative population-genetic characteristics of the spatial position of groups of animals from different countries of origin: based on STR markers (left), based on SNP markers (right): Canada (CAN), Denmark (DEN), Germany (DEU), the Netherlands (NED), Russia (RUS)]
Таблица 3
Субпопуляционный индекс фиксации Fst у изученных пород животных [Subpopulation index of Fst fixation in the studied animal breeds]
|
Красно-пестрая голштинская |
Черно-пестрая голштинская |
Черно-пестрая |
Порода |
|
- |
0.012 |
0.027 |
Красно-пестрая голштинская |
|
0.014 |
- |
0.025 |
Черно-пестрая голштинская |
|
0.008 |
0.012 |
- |
Черно-пестрая |
Над диагональю значения Fst по данным STR (9 STR ), под диагональю по данным SNP.
Таблица 4
Субпопуляционный индекс фиксации Fst у животных из разных стран происхождения [Subpopulation index of Fst fixation in animals from different countries of origin]
|
Германия |
Дания |
Канада |
Нидерланды |
РФ |
Страна происхождения |
|
- |
0.009 |
0.008 |
0.008 |
0.006 |
Германия |
|
0.006 |
- |
0.015 |
0.014 |
0.009 |
Дания |
|
0.010 |
0.024 |
- |
0.015 |
0.011 |
Канада |
|
0.003 |
0.007 |
0.017 |
- |
0.008 |
Нидерланды |
|
0.005 |
0.006 |
0.019 |
0.006 |
- |
РФ |
Над диагональю значения Fst по данным STR (9 STR ), под диагональю по данным SNP.
Таблица 5
Субпопуляционный индекс фиксации Fst в соответствии с годами рождения животных [Subpopulation index of fixation Fst in accordance with the years of birth of animals]
|
1983–1997 |
1999–2003 |
2004–2008 |
2009–2011 |
2012–2014 |
Год рождения |
|
- |
0.007 |
0.008 |
0.016 |
0.015 |
1983–1997 |
|
0.001 |
- |
0.005 |
0.012 |
0.014 |
1999–2003 |
|
0.004 |
0.002 |
- |
0.011 |
0.010 |
2004–2008 |
|
0.017 |
0.017 |
0.013 |
- |
0.006 |
2009–2011 |
|
0.022 |
0.022 |
0.017 |
0.004 |
- |
2012–2014 |
Над диагональю значения Fst по данным STR (9 STR ), под диагональю по данным SNP.
Субпопуляционный индекс между страной происхождения быков-производителей, представленный в табл. 4, демонстрирует схожие значения с соблюдением основных закономерностей, отмеченных при рассмотрении вопроса использования паттернов гомозиготности ROH , отраженных в показателе уровня геномного инбридинга F ROH . К примеру, Fst между РФ и Германией и РФ и Нидерландами по данным STR равен 0.006 и 0.008, по данным SNP 0.005 и 0.006 соответственно.
Показатели анализа Fst по группам года рождения животных, отображенные в табл. 5, проявляют близкий результат, особенно с группами 1983–1999 гг., подчеркивая её наибольшую удаленность от популяции последних лет, когда были получены новые генотипы быков. Значения Fst возрастали вместе с годом, так, по STR они составляли 0.007 в 1999–2003 гг., 0.008 в 2004–2008 гг., 0.016 в 2009–2011 гг., 0.015 в 2012–2014 гг., по SNP 0.001–0.004–0.017–0.022 соответственно.
Таблица 6
Субпопуляционный индекс фиксации Fst у животных разных линий [Subpopulation index of Fst fixation in animals of different lines]
|
Вис Бэк Айди-ал 1013415 |
Монтвик Чифтейн 95679 |
Пабст Говернер 882933 |
Рефлекшн Сове-ринг 198998 |
Силинг Трай-джун Рокит 252803 |
Генеалогическая линия |
|
- |
0.008 |
0.026 |
0.003 |
0.062 |
Вис Бэк Айдиал 1013415 |
|
0.008 |
- |
0.025 |
0.011 |
0.068 |
Монтвик Чифтейн 95679 |
|
0.015 |
0.013 |
- |
0.028 |
0.090 |
Пабст Говернер 882933 |
|
0.004 |
0.009 |
0.017 |
- |
0.065 |
Рефлекшн Сове-ринг 198998 |
|
0.009 |
0.008 |
0.014 |
0.015 |
- |
Силинг Трайджун Рокит 252803 |
Над диагональю значения Fst по данным STR (9 STR ), под диагональю по данным SNP.
Схожесть значений Fst между линиями Вис Бэк Айдиал 1013415 и Монтвик Чифтейн 95679 (0.008) и Рефлекшн Соверинг 198998 (0.003–0.004), представленная в табл. 6, сменяется значительным разбросом в показателях при попарном сравнении с другими линиями. Полагаем, это объясняется малым количеством выборки и меньшей востребованностью остальных линий в селекционном процессе, разведение по которым сократилось всего до трех на сегодняшний день. Сокращение использования генеалогических линий связано с интенсивным отбором быков-лидеров популяции по высоким значениям генетической ценности, а также кроссированием линий для минимизации накопления инбридинга.
Заключение
Проведенные исследования полиморфизма 9 панельных микросателлитных локусов у крупного рогатого скота черно-пестрого корня России показали, что в целом для данного типа маркеров характерен относительно высокий уровень генетического разнообразия ( Ne = 3.577–4.067; Ho = 0.699–0.727). При этом изучаемые популяции скота, представленные быками-производителями, не различались по своей генетической структуре и имели аллели, встречающиеся в других родственных популяциях. Микроса-теллитные профили и генетические дистанции между ними достаточно адекватно отражают их микроэволюцию и специфику селекционного процесса в России на данный момент времени.
В сложившихся условиях сокращение численности племенных быков-производителей, используемых для ведения селекционной работы, приобретает особую актуальность, учитывая возможность снижения гетерогенности популяции.
STR не потеряли своей актуальности и могут являться информативным инструментом в качестве подтверждения достоверности происхождения и составления характеристики аллелофонда исследуемой группы животных [Денискова и др., 2018; Khrabrova et. al., 2019; Бекетов и др., 2021]. Однако, по нашему мнению, SNP -маркеры являются более точным и надежным методом, позволяющим проводить оценку генофонда, вести контроль уровня гомозиготности как на индивидуальном, так и на популяционном уровнях, моделировать желаемые качества выдающихся особей в последующих поколениях, используя методы геномной селекции, вести мониторинг за распространением моногенных рецессивных генетических дефектов [Curik et. al., 2014; Dotsev et. al., 2020].
Формируемая в настоящий момент популяция крупного рогатого скота России по своим генетическим особенностям приближается к мировому генофонду высокоинтенсивных молочных пород. Использование информации по STR- и SNP- маркерам даст возможность более эффективно использовать племенные ресурсы животных посредством разработки программ генетического улучшения скота на региональном и национальном уровнях управления.
Список литературы Популяционно-генетическая характеристика, оценка геномного инбридинга и гомозиготности крупного рогатого скота черно - пестрой и голштинской пород по STR и SNP маркерам в России
- Бекетов С.В. и др. Генетическое разнообразие и филогения пуховых коз центральной и средней Азии // Генетика. 2021. № 7. С. 810-819. DOI: 10.31857/s0016675821070031
- Денискова Т.Е. и др. Изучение генетического разнообразия и дифференциации региональных популяций романовских овец по микросателлитным маркерам // Аграрная наука Евро-Северо-Востока. 2018. Т. 64, № 3. С. 75-80. DOI: 10.30766/2072-9081.2018.64.3.75-80
- Доклады 42 сессии FAO (Food and Agriculture Organization). Италия. Рим, 2021. С. 2021/2Rev1.
- Заид А. и др. Словарь терминов по биотехнологии для производства продовольствия и ведения сельского хозяйства // Продовольственная и сельскохозяйственная организация Объединенных Наций. Рим, 2008. 381 с.
- Кузнецов В.М. Инбридинг в животноводстве: методы оценки и прогноза / НИИСХ Северо-востока. Киров, 2000. 66 с.
- Кузнецов В.М. F-статистики Райта: оценка и интерпретация // Проблемы биологии продуктивных животных. 2014. № 4. С. 80-109.
- Кузнецов В.М. Оценка генетической дифференциации популяций молекулярным дисперсионным анализом (аналитический обзор) // Аграрная наука Евро-Северо-Востока. 2021. № 22(2). С. 167-187.
- Кузнецов В.М., Валохина Н.В. Об ограничении инбридинга в малочисленных популяциях молочного скота // Сельскохозяйственная биология. 2010. № 4. С. 55-58.
- Недашковский И.С. и др. Оценка племенной ценности быков-производителей голштинской породы по качеству потомства в связи с уровнем гомозиготности по STR маркерам // Вестник Рязанского государственного агротехнологического университета им. П.А. Костычева. 2019. № 3(43). С. 36-43.
- Недашковский И.С. и др. Влияние уровня геномного инбридинга, оцененного по ROH-паттернам, на воспроизводительные качества и молочную продуктивность дочерей, a также спермопродукцию голштинских быков-производителей // Достижения науки и техники АПК. 2021. Т. 35, № 3. С. 39-45. DOI: 10.24411/0235-2451-2021-10307.
- Об утверждении Доктрины продовольственной безопасности Российской Федерации: Указ Президента РФ № 20 от 21 янв. 2020 г.
- Сермягин А.А и др. Оценка геномной вариабельности продуктивных признаков у животных голштинизированной черно-пестрой породы на основе GWAS анализа и ROH паттернов // Сельскохозяйственная биология. 2020. Т. 55, № 2. С. 257-274. DOI: 10.15389/agrobiology.2020.2.257rus
- Смарагдов М.Г., Кудинов А.А. Полногеномная оценка инбридинга у молочного скота // Достижения науки и техники АПК. 2019. Т. 33, № 6. С. 51-53. DOI: 10.24411/0235-2451-2019-10612.
- Chang C.C. et al. Second-generation PLINK: rising to the challenge of larger and richer datasets // Giga-Science. 2015. Vol. 4. 7 p. URL: https://doi: 10.1186/s13742-015-0047-8.
- Chao A. et al. Online program SpadeR (Species-richness Prediction And Diversity Estimationin R) // 2016. 88 p. URL: https://doi:10.13140/RG.2.2.20744.62722
- Curik I. et al. Inbreeding and runs of homozygosity: a possible solution to an old problem // Livest Sci. 2014. № 166. P. 26-34. URL: https://doi: 10.1016/j.livsci.2014.05.034.
- Dotsev A.V. et al. PSXII-21 Genome-wide search for genomic regions under putative selection in two russian native cattle breeds using high-density SNP bead chip // J. of Animal Science. 2020. Vol. 98. № 4. P. 242-243. URL: https://doi: 10.1093/jas/skaa278.441
- Ferencakovic M. et al. Estimates of autozygosity derived from runs of homozygosity: empirical evidence from selected cattle populations // J. Anim. Breed Genet. 2013. Vol. 130. P. 286-293.
- Hartl D.L., Clark A.G. Principles of population. United Kingdom: Sunderland, 1997.
- Hedrick P.W. A standardized genetic differentiation measure // Evolution. 2005. Vol. 59, № 8. P. 16331638. URL: https://doi: stable/3449070
- Jost L. GST and its relatives do not measure differentiation // Mol. Ecol. 2008. Vol. 17, № 18. P. 40154026. URL: https://doi: 10.1111/j.1365-294X.2008.03887.x
- Khrabrova L.A. et al. Assessment of line differentiation in the Thoroughbred horse breed using DNA microsatellite loci // Vavilov Journal of Genetics and Breeding. 2019. Vol. 23, № 5. P. 569-574. URL: https://doi: 10.18699/VJ19.526
- Kim E.S., Cole J.B., Huson H. Effect of artificial selection on runs of homozygosity in U. S. Holstein // PLoS One. 2013. Vol. 8, № 11. 80813 p.
- Leutenegger A.L. et al. Estimation of the inbreeding coefficient through use of genomic data // Am. J. Hum. Genet. 2003. Vol. 73. P. 516-523.
- Marras G. et al. Analysis of runs of homozygosity and their relationship with inbreeding in five cattle breeds farmed in Italy // Animal Genetics. 2015. Vol. 46, № 2. P. 110-121.
- Meirmans P.G., Hedrick P.W. Assessing population structure: FST and related measures // Mol. Ecol. Res. 2011. Vol. 11, № 1. P. 5-18. URL: https://doi: 10.1111/j.1755-0998.2010.02927.x
- Peakall R., Smouse P.E. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and researchan update // Bioinformatics. 2012. Vol. 28. P. 2537-2539.
- Purfield D.C. et al. Runs of homozygosity and population history in cattle // BMC Genet. 2012. Vol. 13. 70 p.
- Wright S. Evolution and the genetics of populations. Vol.4 Variability within among natural populations. Univ. Chicago, 1978. 590 p.
- Zhang L. et al. cgaTOH: Extended approach for identifying tracts of // PLoS ONE. 2013. Vol. 8(3). 57772 p. URL: https://doi:10.1371/journal.pone.0057772.
