Популяционные циклы рыжей полевки Myodes (Clethrionomys) glareolus: связь с репродуктивным процессом
 glareolus: связь с репродуктивным процессом")
Автор: Жигальский О.А.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 5-1 т.13, 2011 года.
Бесплатный доступ
На основании многолетних комплексных наблюдений за численностью и демографической структурой населения рыжей полевки, исследованы основные факторы, вызывающие зимнее, ранневесеннее и весеннее размножение. Показано, что зимнее размножение в условиях Удмуртии явление достаточно регулярное и происходит примерно раз в три года. Оценено влияние сроков начала размножения на сезонную динамику и процессы формирования популяционных циклов. Разным срокам начала сезонного цикла размножения мелких млекопитающих свойственна характерная для них сезонная динамика численности и структуры населения. Разнообразное сочетание сроков начала сезонного размножения и экзогенных факторов формирует многолетнюю динамику и все многообразие фазовых переходов численности рыжей полевки в оптимуме ее ареала. Исследование связей между сроками начала сезонного размножения и фазами популяционного цикла показало, что фазе «пик» практически всегда предшествует зимнее размножение, что позволяет популяции достигать высоких численностей уже в середине сезонного репродуктивного периода. Из фазы «пик» популяция во всех случаях переходит в фазу «депрессия». Размножение во все годы наблюдений, относящиеся к фазе «депрессия», начинается в конце апреля - начале мая (весеннее размножение). Между фазами популяционного цикла и сроками начала репродуктивного периода не существует жесткой связи.
Популяционные циклы, демографическая структура, зимнее размножение, репродуктивный процесс, рыжая полевка myodes (clethrionomys) glareolus
Короткий адрес: https://sciup.org/148200317
IDR: 148200317 | УДК: 574:574.3:599.323.4
Population cycles bank vole Myodes (Clethrionomys) glareolus: communication with reproductive process
On the basis of about long-term if complex observations upon numbers and demographic structure of the bank vole population. The most important factors determining launching of reproduction in winter and early spring were studied. It was shown that the phenomenon of reproduction under the snow cover is quite a common event in Udmurtia. This phenomenon seems to be more or less regular, occurring usually once in three years. Since winter reproduction is energy consuming process, then additional energy flow is needed for its success. The value of energy spending in the case of winter reproduction is equal approximately to the half of total energy expenditures for reproduction during the whole year. In the most part of cases, reproduction under the snow cover preceded the peak phase of population cycles. In the phase of depression reproduction usually started in April-May (spring reproduction). However, no rigorous links between phases of population cycles and dates of start of reproduction were discovered phases population the cycle and terms of the beginning of the reproductive period.
Текст научной статьи Популяционные циклы рыжей полевки Myodes (Clethrionomys) glareolus: связь с репродуктивным процессом
Д ля популяций многих видов мелких млекопитающих характерны значительные колебания численности. Некоторые изменения носят кратковременный характер (например, сезонные, протекающие в течение весенне-осеннего периода репродукции), тогда как другие могут продолжаться от 2 до 5 лет [5, 6, 19, 25, 26, 31].
Выживание, воспроизводство и миграции – характеристики, ответственные за пространственнодемографическую динамику популяций мелких млекопитающих, в том числе и в начале сезонного репродуктивного цикла [3, 16, 20, 21, 22, 27, 32].
Высокие весенние численности могут быть вызваны, с одной стороны, благоприятными погодными явлениями, а с другой, низкой смертностью зверьков в осенне-зимний период, а также высокой интенсивностью воспроизводства населения осенью предыдущего года. Низкие численности весной, как правило, связывают с неблагоприятными погодными условиями и высокой смертностью в осенне-зимний период, а также запаздывающим действием высоких численностей прошлого года на качество естественной среды и состояние животных. Чередование низких и высоких весенних численностей и связанные с ними изменения демографической структуры населения определяют цикличность популяционных процессов [3, 4, 9, 11, 26, 28, 33].
Гипотезы, описывающие изменения интервалов между очередными пиками численности, объясняют цикличность популяционных процессов отсроченным плотностно-зависимым эффектом высоких численностей, возникающих во время фазы «пик», на скорость восстановления пространственно-
демографической структуры населения, адекватной условиям среды обитания [1, 3, 10, 12, 13-15, 17, 18, 30]; погодными и пищевыми факторами, а также задержанным плотностно-зависимым эффектом численности хищников на смертность полевок [8, 23, 24, 29].
Цель исследования – анализ особенностей динамики численности и структуры населения рыжей полевки при различных сроках начала сезонного репродуктивного цикла и процессов, определяющих вклад в формирование популяционных циклов разных сроков начала размножения. Работа основана на материалах 29-летних стационарных наблюдений за состоянием популяции рыжей полевки и комплексом внешних условий.
МАТЕРИАЛ И МЕТОДИКА
Материал, используемый в данной работе, собран сотрудниками ЦГСЭН в Удмуртской республике РФ и Института полиомиелита и вирусных энцефалитов. Учеты проводились на стационарных участках вблизи г. Ижевска (56052'СШ, 53010'ВД) в период с 1973 по 2001гг. Стационары расположены на границе двух зон: хвойно-широколиственных и широколиственных лесов (оптимум ареала рыжей полевки). В древостое преобладает липа и ель с примесью пихты и березы. В подлеске преобладают широколиственные породы: клен платановидный, липа, вяз, лещина, бересклет. Травяной покров развит хорошо. Материал собирался стандартным методом ловушко-линий [7]. Отловы проводили регулярно четыре раза в год (апрель, июнь, август и октябрь). За весь период работы отловлено 26893 мелких млекопитающих, в отловах преобладала европейская рыжая полевка (Myodes (Clethrionomys) glareolus Shreber, 1780) - 70,4% от общего числа отловленных животных (18936). За весь период отработано более 103000 ловушко-суток.
Морфологический анализ добытых зверьков проводили по общепринятой схеме. Возраст рыжих полевок определяли по стадии формирования шейки второго верхнего коренного зуба и по доле, которую составляет корень от высоты всего зуба. В работе анализируются три возрастные класса: 7-16; 3-6 и 1-2 месяца [7]. Состояние популяции определяли по оценкам численности и структуры (доле возрастных классов и активных самок и самцов классе).
Для обработки данных доле репродуктивно-в каждой возрастном
использовали методы
одномерной и многомерной статистики: спектральный, дискриминантный анализ. Различия считались достоверными на уровне значимости р <0,05.
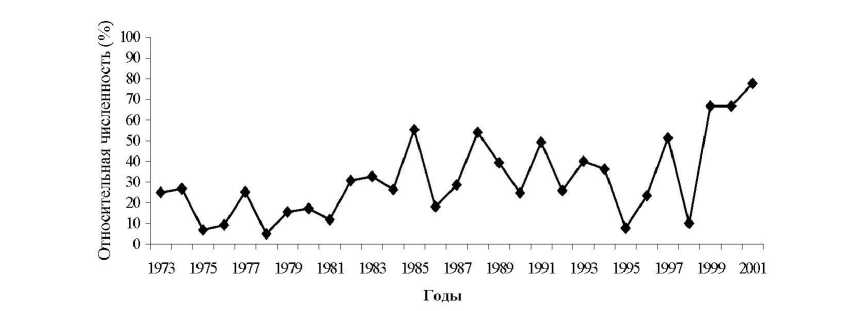
Рис. 1. Многолетняя динамика сезонного пика численности рыжей полевки в елово-липовых лесах Удмуртии
(1973-2001 гг.).
При рассмотрении динамики сроков начала сезона размножения рыжей полевки возникает вопрос, носят ли наблюдаемые колебания случайный характер или в них скрыты некоторые закономерности. Для решения этого вопроса многолетние ряды показателей, характеризующих репродуктивный процесс, в апреле были подвергнуты спектральному анализу. Проведенный анализ показал, что численность беременных второй раз самок и доля сеголеток 3-6 мес. (рожденных в декабре предыдущего года - в январе настоящего) в апреле имеют один пик спектральной плотности, близкий к трехлетнему периоду. Наличие периодичности популяционных процессов позволяет говорить о том, что зимнее размножение в условиях Удмуртии явление достаточно регулярное и происходит примерно раз в три года. Изменение доли 1-2 мес. зверьков (рожденных в феврале - марте) в апреле имеет два пика спектральной плотности - 2 и 3 года, т.е. ранневесеннее размножение явление более редкое, чем зимнее размножение. Для статистической проверки полученных результатов использован дисперсионный анализ, который показал (табл. 1), что для рассматриваемых признаков межгодовая дисперсия статистически значима. Наличие явно выраженных гармонических составляющих в динамике сроков начала репродуктивного цикла, весенней численности, числа и доли размножающихся
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Для исследуемой популяции рыжей полевки характерна высокая и относительно стабильная численность при значительных колебаниях сроков начала и окончания репродуктивного периода, в результате чего возрастной состав населения в разные годы претерпевает значительные изменения (рис.1). В отдельные годы размножение в популяции начиналось в декабре - январе, свидетельство чего высокая доля неполовозрелых сеголеток и репродуктивно-активных самок в апрельских отловах, хотя в некоторые годы в апреле присутствуют беременные второй раз самки. Встречаются годы, когда размножение начинается значительно позже, а репродуктивная активность самок очень низка.
самок и числа прибылых зверьков в апреле, позволяют утверждать, что эти изменения представляют собой закономерный циклический процесс.
Таблица 1. Компоненты межгодовой дисперсии для наиболее информативных показателей, характеризующих начало репродуктивного сезона
|
Показатели (для апреля) |
df |
F |
р |
Объясняемая доля дисперсии,% |
|
Численность беременных самок 2 раз, 7-16 мес. |
29 |
6, 68 |
5E- 10 |
65,9 |
|
Доля 3-6 мес. сеголеток |
29 |
5, 16 |
5,6E -08 |
58,7 |
|
Доля 1-2 мес. сеголеток |
29 |
2, 62 |
0,00 09 |
35,6 |
Наиболее четкой цикличности подвержены сроки начала размножения полевок в декабре-январе (доля объясняемой дисперсии 58,7%) и появление в популяции в апреле 3-4 месячных зверьков. Значительно меньшая доля объясняемой дисперсии, а значит, и степень выраженности цикличности наблюдается в случае начала репродуктивного периода в феврале-марте (табл. 1). Вероятно, если начало размножения приходится на февраль- март, то появление в популяции в апреле 1-2 месячных сеголеток определяется более широким набором факторов.
Межгодовая компонента изменчивости сроков начала размножения и пик ее спектральной плотности статистически значимы, поэтому для разных сроков начала размножения, вероятно, должны существовать годы со сходными сезонными динамиками численности.
Таблица 2. Распределение лет наблюдений по срокам начала репродуктивного сезона рыжей полевки. Результаты пошагового дискриминантного анализа
|
КДФ |
Собственное число |
Коэффициент канонической корреляции, R |
λ -Уилкса |
χ2 |
df |
p |
||
|
0 |
18,77 |
0,97 |
0,01 |
91,27 |
16 |
1,5E-12 |
||
|
1 |
2,53 |
0,85 |
0,28 |
27,11 |
7 |
0,00032 |
||
|
Начало репродуктивного сезона |
Годы |
Начало репродуктивного сезона |
||||||
|
Зимнее |
Ранневесеннее |
Весеннее |
||||||
|
Зимнее n=11 |
1973, 1977, 1980, 1982, 1985, 1988, 1991, 1994, 1997, 1999, 2001 |
F= 25,2 p ≤ 2,97E-08 |
F= 38,6 p ≤ 8,8E-10 |
|||||
|
Ранневесеннее n=4 |
1979, 1984, 1993, 1996 |
66,0 |
F= 7,3 p ≤ 0,00024 |
|||||
|
Весеннее n=14 |
1974, 1975, 1976, 1978, 1981, 1983, 1986, 1987, 1989, 1990, 1992, 1995, 1988, 2000 |
101,2 |
20,3 |
|||||
Примечание: Под диагональные элементы - квадрат расстояния Махаланобиса между центроидами групп, над диагональные - F - критерий (df 8,18) и p - уровень значимости.
Таблица 3 . Демографические характеристики рыжей полевки (М±m) при зимнем, ранневесенним и весенним начале размножения, (непараметрический анализ Kruskal-Wallis ANOVA)
|
Начало размножения |
Значимые контрасты ( р <0,05), ( р <0,1*) |
|||||
|
Весеннее I |
Ранневесеннее II |
Зимнее III |
||||
|
5 s 2 H
|
Й < |
общая численность |
37,0 ± 2,72 |
26,8 ± 4,41 |
25,7 ± 3,33 |
I – от всех |
|
общая доля беременных ♀♀ |
1,6 ± 1,10 |
8,7 ± 3,65 |
17,3 ± 3,23 |
III – от всех, *Все |
||
|
доля группы 7-16 мес. |
5,5 ± 0,63 |
3,8 ± 0,65 |
3,3 ± 0,82 |
I – III, I – (II+III) |
||
|
доля группы 3-6 мес. |
7,7 ± 1,04 |
15,3 ± 2,99 |
17,2 ± 1,55 |
I – от всех, *Все |
||
|
доля группы 1-2 мес. |
86,8 ± 1,13 |
80,9 ± 3,03 |
79,5 ± 1,69 |
I – III, I – (II+III), *Все |
||
|
среднее число эмбрионов |
1,4 ± 0,46 |
2,5 ± 0,54 |
4,6 ± 0,36 |
III – от всех, *Все |
||
|
-Q Ю ch и О |
общая численность |
21,9 ± 2,54 |
16,2 ± 2,93 |
17,5 ± 2,49 |
*I – от всех |
|
|
общая доля беременных ♀♀ |
0,0 ± 0,00 |
0,0 ± 0,00 |
0,4 ± 0,39 |
Нет |
||
|
доля группы 7-16 мес. |
2,6 ± 0,73 |
2,4 ± 0,65 |
2,2 ± 0,65 |
Нет |
||
|
доля группы 3-6 мес. |
59,1 ± 6,05 |
44,9 ± 6,79 |
38,5 ± 5,43 |
I – III, I – (II+III) |
||
|
доля группы 1-2 мес. |
38,3 ± 6,17 |
52,7 ± 7,13 |
56,0 ± 5,59 |
I – III, I – (II+III) |
||
|
-Q
< |
общая численность |
5,4 ± 0,89 |
8,0 ± 1,43 |
13,3 ± 1,64 |
III – от всех |
|
|
доля беременных ♀♀ 1 раз (7-16 мес.) |
48,8 ± 8,00 |
69,7 ± 7,27 |
43,9 ± 5,18 |
II – III, II – (I+III), *I - II |
||
|
доля беременных ♀♀ 2 раз (7-16 мес.) |
0,0 ± 0,00 |
0,0 ± 0,00 |
35,5 ± 5,32 |
III – от всех |
||
|
доля беременных ♀♀ (7-16 мес.) |
48,8 ± 8,00 |
69,7 ± 7,27 |
79,5 ± 3,14 |
I – III, I – (II+III), *I - II |
||
|
доля беременных ♀♀ (3-6 мес.) |
0,0 ± 0,00 |
13,0 ± 6,17 |
35,0 ± 8,35 |
Все |
||
|
доля беременных ♀♀ (1-2 мес.) |
0,0 ± 0,00 |
0,7 ± 0,74 |
35,2 ± 6,91 |
III – от всех |
||
|
доля группы 7-16 мес. |
96,3 ± 3,70 |
95,6 ± 2,36 |
76,7 ± 3,72 |
III – от всех |
||
|
доля группы 3-6 мес. |
0,0 ± 0,00 |
1,1 ± 0,58 |
5,3 ± 1,26 |
Все |
||
|
доля группы 1-2 мес. |
0,0 ± 0,00 |
3,3 ± 2,33 |
18,0 ± 3,35 |
Все |
||
|
зимняя выживаемость |
15,1 ± 2,20 |
34,1 ± 4,70 |
44,0 ± 3,44 |
Все |
||
|
среднее число эмбрионов |
3,0 ± 0,54 |
5,6 ± 0,42 |
6,4 ± 0,11 |
I – от всех |
||
Все годы наблюдений были разделены по срокам начала размножения на зимнее, ранневесеннее и весеннее. Проверку и уточнение данной классификации провели с использованием пошагового дискриминантного анализа и получили три значимо различающиеся по срокам начала размножения группы лет наблюдений (табл. 2). Разделение лет по срокам начала репродуктивной активности было проведено по 12 демографическим характеристикам, из которых дискриминантный анализ позволил выделить наиболее информативные (табл. 1). Как видно из таблицы, что характеристики лет, относящиеся к зимнему, ранневесеннему и весеннему размножению, статистически значимо отличаются друг от друга. По совокупности характеристик популяции более значительно различаются зимнее и весеннее начало размножения, а наиболее близки ранневесеннее и весеннее (табл. 3).
Зимнее размножение, которое начинается в декабре–январе, наблюдалось 11 раз (табл. 2). Для этих лет в апреле характерна наибольшая численность и доля беременных самок (у перезимовавших и сеголеток). Кроме того, только в эти годы присутствуют беременные второй раз самки (табл. 2). Зимнее начало размножения также характеризуется высокой зимней выживаемостью и самым большим числом эмбрионов на одну самку.
Ранневесеннее размножение наблюдалось 9 лет наблюдений, в эти годы размножение начинается в конце февраля в начале марта. Животные этой группы лет характеризуются более высокой смертностью перезимовавших и поздними сроками вступления в размножение перезимовавших самок, в результате чего и низкой общей численностью в апреле (табл. 3). Кроме того, в этой группе лет небольшое число прибылых зверьков и доля половозрелых среди них, но в классе 3-6 месячных полевок вообще отсутствуют половозрелые животные (табл. 3).
Весеннее размножение начинается в конце апреля - начале мая включает 9 лет наблюдений. В этой группе лет в апреле самая низкая общая численность животных, численность и доля беременных самок (в некоторые годы в это время они отсутствуют вообще), нет половозрелых сеголеток, значимо ниже других сроков начала размножения число эмбрионов (табл. 3). Смертность зимующих зверьков этой группы лет почти в 3 раза выше, чем при зимнем размножении, и в 2 раза выше, чем при ранневесеннем начале репродуктивного периода (табл. 3).
Различным срокам начала размножения сопутствуют различные весенние численности, которые определяют последующую сезонную динамику (Friedman ANOVA Chi Sqr. (N = 44, df = 1) = 12,3 p < 0, 00045). На рисунке 2 ранневесеннее размножение не рассматривалось из-за малого числа лет входящих в эту группу. Для лет с весенним размножением характерна низкая численность в апреле и на протяжении всего репродуктивного периода. Годы с зимним размножением имеют более высокую сезонную амплитуду (рис. 2А). При этом следует отметить, что в октябре предыдущего года численности полевок в обоих случаях были практически равны, но из-за разной зимней выживаемости животных (62,0 и 26,9% соответственно для зимнего и весеннего начала сезонного размножения) их численности весной различаются почти в два раза (рис. 2А). Различия в выживаемости зверьков в осенне-зимний период главным образом связаны с различиями в возрастной структуре населения осенью предыдущего года. Зимнему размножению предшествует высокая доля в популяции 1-2 месячных зверьков (рис. 2С), которые, как правило, неполовозрелы и поэтому их смертность в зимний период значительно ниже, чем у полевок раннелетних пометов (возраста 3-6 месяцев) в большинстве своем половозрелых.
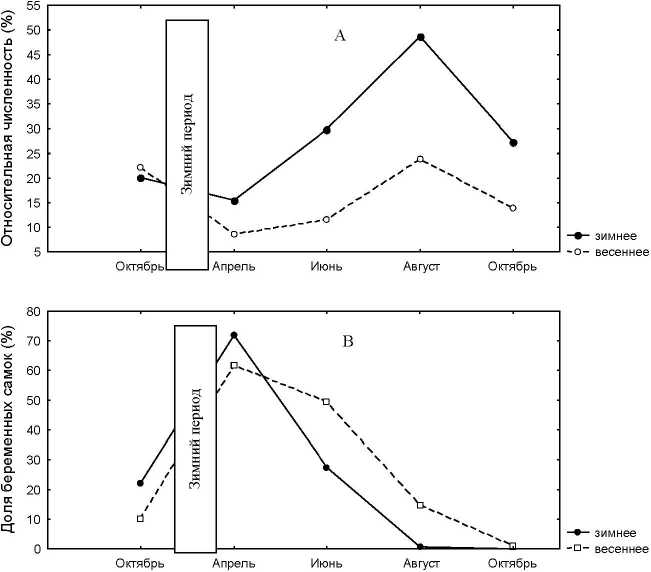
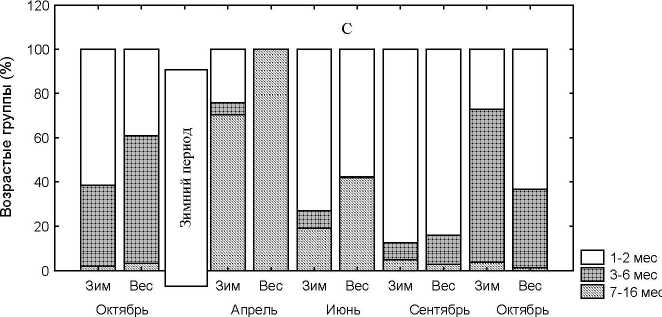
Рис. 2 . Сезонная динамика демографических характеристик рыжей полевки при разных сроках начала сезонного размножения. А – численность. В – доля беременных самок. С – возрастная структура, надписи «зим» и «вес» соответственно зимнее и весеннее начало размножения
Доля беременных самок в апреле при зимнем размножении достигает 71,1%, а при весеннем 61,5%, но на протяжении последующего репродуктивного периода доля репродуктивно активных самок при весеннем размножении выше и даже в октябре встречаются беременные самки (рис. 2В). Различия в долях беременных самок при зимнем и весеннем начале размножения статистически достоверны (Friedman ANOVA Chi Sqr. (N =44, df = 1) = 12,37 p<0,00045). Результат этих различий состоят в том, что при зимнем размножении наибольшее число беременных самок, а значит и прирост популяции приходится на первую половину лета, а при весеннем на вторую. В результате чего формируется разная функциональная и возрастная структура популяции. В первом случае основу населения уходящего в зиму составляют зверьки раннелетних пометов, а во втором животные, родившиеся во вторую половину лета.
Зимнему размножению предшествует высокая доля в популяции 1-2 месячных зверьков, а весеннему 3-6 месяцев (рис. 2С). Соотношение возрастных групп в популяции в апреле, июне и сентябре при различных сроках начала размножения статистически неотличимы. В апреле во всех типах начала сезонного размножения популяция в основном состоит из перезимовавших зверьков. В июне и в августе во всех случаях основу населения составляют 1-2 месячные животные. В октябре при зимнем размножении в популяции преобладают 3-6 месячные зверьки, а при весеннем начале сезонного размножения основную часть населения составляют 1-2 месячные (рис. 2 С). Высокая доля 3-6 месяцев в октябре при зимнем начале размножения обусловлена тем, что уже в начале июля общая численность полевок достигает высоких значений, в результате чего практически полностью прекращается размножение перезимовавших и прибылых самок и поэтому основу населения (69%) осенью составляют зверьки раннелетних пометов. Для малых численностей, характерных для весеннего начала размножения, плотностно-зависмых ограничений, как активности участия в размножении животных всех возрастных групп, так и в половом созревании прибылых зверьков нет в течение всего сезона размножения, и поэтому основу осенней популяции составляют неполовозрелые зверьки 1-2 месяцев.
В оптимуме ареала рыжей полевки было выделено три типа сезонных динамик - «пик», «депрессия» и «рост», чередование которых имеет близкий к 3 летнему популяционный цикл [5, 6]. Подобного можно было ожидать и при чередовании типов сезонных динамик, соответствующих различным срокам начала размножения. Как мы видели ранее, демографические характеристики популяции в апреле изменяются с периодом 2 - 3 года. Однако многолетняя динамика переходов из разных сроков начала сезонной репродуктивной активности в другие не имеет таких строгих закономерностей, как в фазовых переходах популяционного цикла. Так, например, после зимнего начала репродуктивного периода на следующий год с той или иной вероятностью размножение может начаться в любой другой период (зимой, весной ранней весной). Зимнее размножение в предыдущем году определяет начало репродуктивного периода на следующий год зимой, которое повторялось через 2 - 4 года. Зимнее размножение в 6 случаях наступало после ранневесеннего размножения и в 4 - после весеннего.
Фаза «пик» практически всегда (кроме 1984 и 1989 годов) сопровождается зимним размножением, что позволяет популяции достигать высоких численностей уже в средине сезонного репродуктивного периода [6]. Из фазы «пик» популяция во всех случаях переходит в фазу «депрессия». Годы «пик» характеризуются высокой численностью животных, низкой скоростью полового созревания, снижением жизнеспособности потомства, малой долей репродуктивно-активных зверьков и низкой выживаемостью в осенне-зимний период. Размножение во все годы относящихся к фазе «депрессия» начинается в конце апреля - начале мая (весеннее). Для фазы «депрессия» характерна низкая общая численность, небольшое число репродуктивноактивных животных, низкая скорость полового созревания молодых зверьков, что не позволяют популяции за один сезон размножения достигать высоких численностей [6].
ЗАКЛЮЧЕНИЕ
Приуроченность репродуктивных циклов животных к весенне-летнему сезону определяет дискретность популяционной динамики, а поэтому может рассматриваться как один из основных факторов, вызывающих запаздывание реакции популяции на изменения внешних и внутренних условий. Наличие подобного рода запаздывания, высокая смертность и высокий репродуктивный потенциал вида, плотностно-зависимая регуляция, влияние хищников и целый ряд других факторов могут стать причинами возникновения, как популяционных автоколебаний, так и хаотического режима изменения демографических параметров.
Демографическая структура населения осенью предыдущего года, зимняя выживаемость, обилие зимних кормов, метеорологические условия зимовки, время наступления сезонного цикла размножения, весенняя численность рассматриваемого года определяют сезонную динамику популяции. Зимнее и ранневесеннее размножение рыжей полевки в Удмуртии явление достаточно регулярное и происходит примерно раз в два-три года. Наличие многолетнего ряда позволило все годы наблюдений разделить на три группы лет по срокам начала размножения: зимнее, ранневесеннее и весеннее. Каждой группе лет соответствует характерная только для него сезонная динамика изменения демографических характеристик населения (рис.2 A,B,C). Различным срокам начала размножения сопутствуют различные весенние численности, которые определяют последующую сезонную динамику. При высокой весенней численности 68% от общего числа прибылых - потомки перезимовавших, 32% -от зверьков первых пометов [2]. Так, если размножение начинается при значительном числе перезимовавших, то уже во второй половине июня плотность популяции начинает превышать критический уровень, что снижает интенсивность размножения. Для низких численностей характерно позднее (на полтора месяца) «включение» плотностнозависимых обратных связей, что обусловливает появление дополнительных пометов. Однако, при высоких весенних численностях, общее число прибылых животных больше, чем при низких. При низкий весенних численностях число прибылых увеличивается пропорционально числу самок; превышение критического уровня нарушает пропорциональность прироста, что связано с повышенной смертностью молодых и лишь частичным участием в размножении половозрелых зверьков [2]. Кроме того, высокая численность приводит к увеличению сезонного пика и подъему уровня осенней численности. Но для каждых конкретных условий существует такая численность, превышение которой вызывает снижение, как сезонного пика, так и осенней численности. Основные причины раннего ограничения размножения - высокие численности и различия в динамике прибылых в годы малых и больших численностей [3]. Наиболее четко этот эффект проявляется при значительных перепадах весенних численностей. Так, прирост популяции за май - июнь в год низкой численности составляет 42%, а в год высокой — 68%. Во втором случае основную часть сеголеток составляют зверьки первых трех пометов, которые погибают к осени, что является причиной снижения осенней численности. Когда размножение начинается с малого числа зверьков, популяция медленно наращивает репродуктивный потенциал, в результате чего большая часть прибылых (58%) приходится на вторую половину лета. Зверьки родившиеся во вторую половину лета не размножаются в год своего рождения, поэтому большинство из них успешно переживают зиму и формируют высокую весеннюю численность следующего года. Поэтому становится понятным резкое падение численности в годы, следующие за годами с высокой весенней численностью. Главная причина, описанных процессов - малое число зверьков, уходящих в зиму (32%), в то время как в годы с малой весенней численностью процент таких зверьков повышается до 58. Тогда в первом случае на следующий год наблюдается закономерный спад численности, а во втором - ее подъем. Если популяция за один сезон размножения сможет повысить свою численность до высокого уровня, возникнут двухлетние колебания, если это произойдет за два сезона, то возникнет трехлетний цикл, и так далее [2].
Кроме описанных эффектов, раннее размножение и следующая за этим высокая весенняя численность почти на месяц сокращает длительность периода размножения. Последние прибылые в этом случае выходят из гнезд 20 - 22 августа, при низкой – 10 - 12 сентября. Изменение интенсивности размножения и его длительности приводит к перестройке структуры популяции (рис. 2 A,B,C). При высокой численности в начале июля резко падает процент прибылых половозрелых зверьков в популяции с полным исчезновением этой группы к первой декаде сентября, но параллельно этому возрастает число неполовозрелых. При низкой численности группа молодых (1 –2 месячных) зверьков на протяжении всего периода размножения остается наиболее многочисленной, а последние молодые исчезают из популяции только в конце сентября (табл.3). Вследствие высокой плотности удлиняется срок полового созревания настолько, что основная часть сеголеток остается неполовозрелой [3, 11]. Динамика численности и структуры населения определяется не только процессами, протекающими в популяции в текущем году, но и ее состоянием в предыдущем, в частности она зависит от того, особи каких пометов принимают участие в формировании весенней численности следующего года.
Группа животных уходящих зиму после фазы «пик» и служащих основой населения следующей за ней фазы «депрессия», состоит главным образом из зверьков раннелетних пометов, развитие которых проходили при высоких плотностях, и, вероятно, как следствие этого - высокая зимняя смертность и низкий репродуктивный потенциал популяции на следующий год. Популяция в фазу «депрессия», начинает свой репродуктивный сезон значительно позднее. Этой фазе, как правило, предшествует плохая перезимовка (низкая урожайность липы и ели и ранний сход снега).
Исследование связей между сроками начала сезонного размножения и фазами популяционного цикла показало, что фазе «пик» практически всегда предшествует зимнее размножение, что позволяет популяции достигать высоких численностей уже в средине сезонного репродуктивного периода. Из фазы «пик» популяция во всех случаях переходит в фазу «депрессия». Размножение во все годы наблюдений, относящиеся к фазе «депрессия» начинается в конце апреля – начале мая.
Автор выражает глубокую признательность А.Д. Бернштейн, А.В. Хворенкову и С.П. Трушину за предоставленные материалы и обсуждение полученных результатов.
Работа выполнена при поддержке гранта РФФИ 10-04-01657
Список литературы Популяционные циклы рыжей полевки Myodes (Clethrionomys) glareolus: связь с репродуктивным процессом
- Большаков В.Н., Кубанцев Б.С. Половая структура популяций млекопитающих и ее динамика. М. Наука, 1984.233 с.
- Жигальский О.А. Исследование плотностно-зависимых механизмов регуляции численности красной полевки. Имитационное моделирование.//Журн. общей биол. 1982. Т. XLIII, №1. С. 121-128.
- Жигальский О.А. Анализ популяционной динамики мелких млекопитающих//Зоол. журн. 2002. Т. 81, № 9. С. 1078-1106.
- Жигальский О.А., Белан О.Р. Пространственно -временная динамика полевок в гетерогенных местообитаниях Иремельского горного массива.//Известия РАН. Серия биол., 2004. № 2, С. 238-246
- Жигальский О.А., Кшнясев И.А. Популяционные циклы европейской рыжей полевки в оптимуме ареала//Экология. 2000. №5. С. 383-390
- Жигальский О.А., Хворенков А.В., Бернштейн А.Д. Циклы численности и демографическая структура в популяциях мелких млекопитающих//Многолетняя динамика численности птиц и млекопитающих в связи с глобальными изменениями климата: Материалы международного симпозиума (11-16 ноября 2002) -Казань. 2002. С. 30-38.
- Карасева Е. В., Телицына А. Ю., Жигальский О. А. Методы изучения грызунов в полевых условиях. М. из-во. ЛКИ. 2008. 416 с.
- Кузнецов Г.В, Михайлин А. П. Особенности питания и динамики численности рыжей полевки в условиях широколиственного леса.//Млекопитающие в наземных экосистемах. М. из-во. Наука. 1985.С. 127-156
- Мамина В.П., Жигальский О.А., Оценка оплодотворяющей способности сперматозоидов у рыжей полевки на разных фазах динамики численности.//Успехи современной биологии. 2006. Т. 126, №4. С. 414-421.
- Маркина Т.А. Влияние половодья на сезонную динамику численности и структуру населения мелких млекопитающих юго-востока Мещеры.//Изв. Самар. НЦ РАН. 2010. Т. 12, № 1. С. 147-152.
- Роговин К.А., Мошкин М.П. Авторегуляция численности в популяциях млекопитающих и стресс (штрихи к давно написанной картине).//ЖОБ. Т. 68, № 4. 2007. С. 244-267
- Чернявский Ф.Б, Лазуткин А.Н. Циклы леммингов и полевок на Севере. Магадан. 2004. 105 с.
- Шварц С.С. О возрастной структуре популяций млекопитающих.//Труды Уральского отделения МОИП, вып.2. Свердловск, 1959. С.3-22.
- Шварц С.С. Возрастная структура популяций животных и проблемы микроэволюции (теоретический анализ проблемы).//Зоологический журнал. 1965. Т.44, вып.10. С.1443-1453.
- Шварц С.С., Оленев В.Г., Кряжимский Ф.В., Жигальский О.А. Исследование динамики численности и возрастной структуры популяции мышевидных грызунов на имитационной модели//Докл. АН СССР. 1976. Т.228, N6. С.1482-1485.
- Batzli, G. O. Population cycles revisited.//Trends in Ecology and Evolution. 1996. 11, Р. 448-449.
- Christian, J. J. Population density and reproductive efficiency.//Biology of Reproduction. 1971. 4. P. 248-294
- Christian, J. J. Endocrine factors in population regulation. in Biosocial mechanisms of population regulation (M. N. Cohen, R. S. Malpass, and H. G. Klein, eds.).//Yale University Press, New Haven, Connecticut. 1980. P. 367-380
- Bjoernstad, O. N., Stenseth, N. C., Saitoh, T. & Lingjaerde, O. C. Mapping the regional transition to cyclicity in Clethrionomys rufocanus: special densities and functional data analysis.//Research in Population Ecology. 1998. 40. Р. 77-84.
- Dueser, R. D., Wilson, M. L. & Rose, R. K. Attributes of dispersing meadow voles in open-grid populations.//Acta Theriologica. 1981. 26. Р. 139-162.
- Gaines, M. S. & McClenaghan, L. R. Dispersal in small mammals.//Annual Review of Ecology and Systematics. 1980. 11. Р. 163-196.
- Getz, L. L. Vole population fluctuations: why and when.//Acta Theriologica Sinica. 2005. 25. Р. 209-218.
- Huitu O, Norrdahl K, Korpimäki E. Landscape effects on temporal and spatial propertiesof vole population fluctuations.//Oecologia 2003. 135. Р.209-220
- Klemola, T., Koivula, M., Korpimaeki, E. & Norrdahl, K. Experimental tests of predation and food hypotheses for population cycles of voles.//Proceedings of the Royal Society of London, B. Biological Sciences. 2000. 267. P. 351-356.
- Krebs, C. J. Population cycles revisited.//Journal of Mammalogy. 1996. 77. Р. 8-24
- Krebs C. J., Myers J. H. Population cycles in small mammals//Adv. Ecol. Res. 1974. Vol. 8, -P. 267-399.
- Lin, Y. K. Batzli, G. O. The influence of habitat quality on dispersal, demography and population dynamics of voles.//Ecological Monographs. 2001. 71. Р. 245-275.
- Millon A & Bretagnolle V. Nonlinear and population-specific offspring sex ratios in relation to high variation in prey abundance//OIKOS. 2005. 108. P. 535-/543,
- Norrdahl, K. & Korpimaeki, E. Do nomadic avian predators synchronize population fluctuations of small mammals a field experiment. Oecologia. 1996. 107. P. 478-483
- Norrdahl, K. & Korpimaeki, E Changes in individual quality during a 3-year population cycle of voles.//Oecologia..2002.130. P. 239-249.
- Taitt, M. J. & Krebs, C. J. Population dynamics and cycles. in Biology of New World Microtus (R. H. Tamarin, ed.)//Special Publication 8, The American Society of Mammalogists. 1985. P. 567-620.
- Verner, L. & Getz, L. L. Significance of dispersal in fluctuating populations of Microtus ochrogaster and M. pennsylvanicus.//Journal of Mammalogy, 1985. 66. Р. 338-347.
- Zhigalsky O.A. Factorial analysis of population dynamics in rodents.//Polish ecological studies, 1993. V.18. No.1-2. Quarterly. P. 3-158.
