Популяционные реакции Galium odoratum (L.) Scop. (Rubiaceae) на смену условий местообитания
 Scop. (Rubiaceae) на смену условий местообитания")
Автор: Фдорова Светлана Владиславовна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Флористика
Статья в выпуске: 1-7 т.14, 2012 года.
Бесплатный доступ
В статье представлены результаты сравнительного морфоструктурного анализа пяти ценопопуляций Galium odoratum (L.) Scop. в подзоне хвойно-широколиственных лесов. Описаны изменения показателей по градиентам трёх экологических факторов среды. Это - ценотическое затенение, влажность почвы и богатство почвы доступными для растений формами азота. Проведена ординация местообитаний в системе экологических координат.
Популяция, факторы, экологические координаты, ординация, длиннокорневищное травянистое растение
Короткий адрес: https://sciup.org/148200949
IDR: 148200949 | УДК: 582.4/9-15,
Galium odoratum (L.) Scop. (Rubiaceae) population reactions on change of habitat conditions
In article results comparative morph-structure the analysis of five Galium odoratum (L.) Scop. cenotic population are submitted In a sub-zone of coniferous and widely-deciduous woods. Are described changes of parameters on gradients of three factors environment ecological. It – cenotic shadow, humidity of soil and riches of soil forms of nitrogen accessible to plants. It is carried out ordynation habitats in system of ecological coordinates.
Текст научной статьи Популяционные реакции Galium odoratum (L.) Scop. (Rubiaceae) на смену условий местообитания
Проблемы сохранения биоразнообразия и рационального использования растительных ресурсов решаются ботаниками на популяционном и фито-ценотическом уровне организации жизни. Растительная популяция в моем понимании представляет собой самоорганизованную и саморазвивающуюся систему особей одного вида. Реагируя на изменение условий местообитания, популяции изменяют свою организацию для наиболее рационального использования пищевых ресурсов. Каковы эти реакции у Galium odoratum (L.) Scop . (Rubiaceae) -растения, которое, начиная с третичного периода, пережило множество сменяющих друг друга местообитаний? Получить ответ на этот вопрос - цель настоящего исследования. G.odoratum – гипогео-генно-длиннокор-невищное растение с симподи-альным нарастанием. Имеет моно- и дицикличе-ские ортотропные ассимилирующие побеги высотой до 35 см с мутовками по 6-8 шт. ланцетных сидячих листьев. Цветки формируются в верхушечных полузонтиках, плоды односемянные ореховидные. Корневище представляет собой столон -одно-двухлетний побег с чешуевидными листьями, в каждом узле которого может сформироваться ассимилирующий побег. Столоны имеют плагио-тропный рост и располагаются в слое лесной подстилки или в поверхностном слое почвы. В подзоне хвойно-широколиcтвенных лесов, куда входит район исследования (Республика Татарстан, Васильевское лесничество) для самоподдержания популяции одинаково важно семенное размножение и вегетативное, которое осуществляется по принципу неглубоко омоложенной партикуляции. Вегетативное размножение приводит к формированию диффузных клонов. Нарастание столонов, формирование придаточных корней и ассимилирующих побегов происходит на протяжении всего вегетационного сезона. Цветение наблюдается в мае, плодоношение в июне-июле [1].
Регулярный отбор контрольных особей (по 50 экз.) для морфоструктурного анализа проведён ле-
том 2001 г. в 5-ти ценопопуляциях (ЦП) на серой лесной почве. Особь - полицентрическая система со всей системой столонов и ассимилирующих побегов. Условия местообитаний оценены с помощью экологических координат [2], представленных в табл. 1. Характеристика местообитаний дана в табл. 2. Обработка данных проведена с помощью пакета анализа, заключённого в редакторе Microsoft Excel. Парный двух выборочный F-тест для дисперсий использован для выявления различий между вариационными рядами показателей в разных ЦП (таб. 3). Приём построения точечных диаграмм с подбора наиболее достоверных аппроксимаций с помощью функции «Мастер диаграмм» использован для выявления зависимости морфоструктурного показателя (у) от изменения фактора (х) (рис. 1). Прием построения лепестковой диаграммы использован для проведения ординацию местообитаний G.odoratum в трёхмерной системе экологических координат (рис. 2).
ЦП 1 в березово-пролесниково-снытевом (Betula pendula L. - Aegopodium podagraria L.+ Mercurialis perennis L.) фитоценозе с Acer platanoides L. и Sor-bus aucuparia L. в подлеске имела большое покрытие (36 %). Единичные особи G.odoratum (6 % выборки) плодоносили с образованием 4-8 шт. плодов. Они были малоурожайными. Дефицит органов генеративного размножения компенсировался наиболее интенсивным ростом органов, обеспечивающих вегетативное размножение. ЦП II в липново-чистецово-разнотравном (Tilia cordata Mill. - mix herbs + Stachys sylvatica L.) фитоценозе с Acer pla-tanoides и Corylus avelana L. в подлеске имела покрытие 16 %. Редкие особи (2 % выборки) G. odoratum плодоносили и были малоурожайными. Дефицит генеративных органов компенсировался более интенсивным развитием органов, обеспечивающих вегетативное размножение. Среднестатистические особи отличались большим количеством узлов на столонах. ЦП III в липово-осоково- pillosa, Picea fennica (Regel) Kom.) фитоценозе разнотравном с елью (T.cordata - mix herbs + Carex
Таблица 1 . Экологические координаты и формулы для их расчёта
|
Название и символ |
Формула |
Примечание |
|
Ценотический коэффициент затенения (англ.: coefficient of shadow from plants) СSP, % Ценотический индекс богатства почвы азотом (англ.: cenotic Index of the Nitrogen-rich soil) NtRS, % Влажность почвы (англ.: humidity of soil) HS, % |
= V--L V4-L V-W:! CSP=( а - сомкнутость крон деревьев; b - сомкнутость кустарников; с - покрытие растений из травяно-кустарничкоаого яруса, которые отбрасывают густую тень на растение-объект. Это широколистные формы растений и/или мелколистные, но с сильно разветвлёнными побегами и густо расположенными листьями. Всё в процентах. NtRS = 100 (^a-^b)/Y(a + Ы-c) а - покрытие нитрофильного и/или субнитро-фильного вида; b - покрытие анитрофильного и/или субанитрофильного вида; с - покрытие вида с широкой экологической амплитудой в режиме богатства почвы азотом. Z100S(Pl- Р2Л HS = ---------- /Г1 \ P1 / Р1 - вес влажной почвы; Р2 - вес сухой почвы; n – число проб (n = 8). |
СSP = 100 % - абсолютная величина затенения. Она обуслов-леноа 100 % смыканием крон деревьев, 100 % смыканием кустарников и 100 % покрытием травянистых и кустарничковых растений над местом произрастания вида-объекта. Отношение растений к азоту определено по шкале «Богатства почв азотом - Nt» [3]. Виды с диапазоном не шире «jm» считались анитрофильными и субанитро-фильными, не шире «+lo» - суб-нитрофильными и нитрофиль-ными. Покрытие видов оценено в баллах по шкале КТШ-5 [4], где интервалы покрытия 0-4-1636-64-100 (%) соответствуют баллам 1, 2, 3, 4, 5. Влажность почвы определена на глубине корнеобитаемого слоя растения-объекта (5-10 см). |
Таблица. 2 . Характеристика местообитаний Galium odoratum. Данные 28.06.2001
|
К ^ |
Деревья-эдификаторы |
H |
& о ° О cd К & |
Координаты |
||||
|
возраст, лет |
высота, м |
диаметр стволов на уровне груди, см |
СSP, % |
HS, % |
NtRS, % |
|||
|
I |
60-70 |
25 |
40-45 |
80 |
90 |
54 |
25 |
67 |
|
II |
40-45 |
25 |
25-30 |
70 |
75 |
43 |
23 |
75 |
|
III |
40-45 |
25 |
25-30 |
60 |
65 |
30 |
18 |
39 |
|
IV |
40-45 |
25 |
35-40 |
80 |
70 |
53 |
20 |
85 |
|
V |
30-40 |
18 |
20-30 |
50 |
65 |
28 |
12 |
37 |
Таблица 3 . F-критерий для сравнения морфоструктурных показателей в ценопопуляциях Galium odoratum . Данные 28.06.2001-2.07.2001
Из проведённого исследования были сделаны выводы: 1. Проведение ординации местообитаний G. odoratum в трёхмерной системе экологических координат позволяет учёсть комплекс основных факторов среды и предсказать наиболее благопри-
ятные условия для развития растения; 2. Расчёт ценотического коэффициента затенения, ценотиче-ского индекса богатства почвы азотом и влажности почвы способствовал выявлению точек, в которых особи в популяционной системе Galium odoratum развиваются наиболее активно: CSP = 28-43 %, NtRS = 77- 75 %, HS = 18-25 %; 3. Изменения в популяционной организации G. odoratum по градиентам 3-х эколого-ценотических факторов вполне предсказуемы. Изменения морфоструктурных показателей роста и размножения имеют полиномиальный характер.
Примечание: *, **, *** - означает то, что F статистики больше F критическое на уровне значимости 90, 95, 99 %
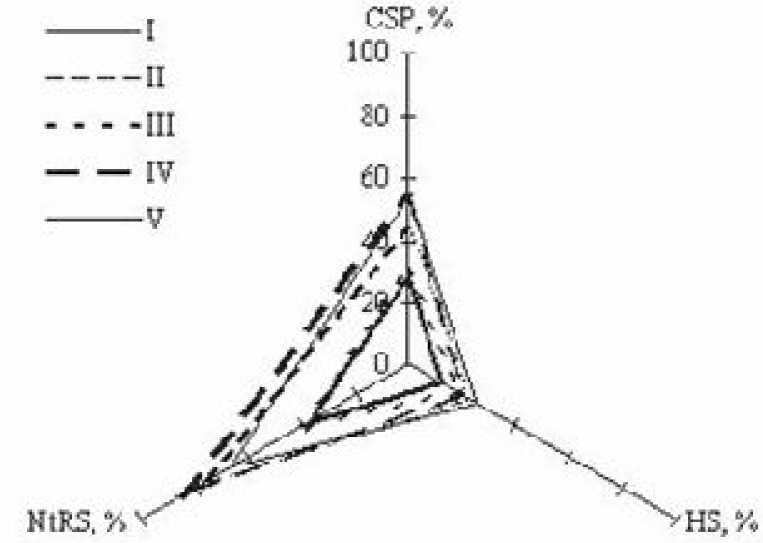
Рис. 2 . Ординация местообитаний Galium odoratum в трёхмерной системе экологических координат. I-V - №
ЦП
Список литературы Популяционные реакции Galium odoratum (L.) Scop. (Rubiaceae) на смену условий местообитания
- Смирнова О.В. Структура травяного покрова широколиственных лесов. М.:Наука, 1987. 139 с.
- Фёдорова С.В. Популяционные отклики Fragaria vesca L. (Rosaceae) на смену эколого-фитоценотических факторов//Труды Тигирекского заповедника. 2010. Вып. 3. С. 160-165.
- Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-широколиственных лесов. М.: Наука, 1983. 196 с.
- Любарский Е.Л. Об оценке проективного покрытия компонентов травостоя//Экология. 1974. № 1. С. 155-158.
