Популяционный мониторинг сукцессий степной растительности на территории ботанического памятника природы местного значения «Балка Сухая»

Автор: Остапко Владимир Михайлович, Ибатулина Юлия Валериевна, Зыбенко Ольга Валериевна
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 3-7 т.15, 2013 года.
Бесплатный доступ
В статье рассмотрено состояние ценопопуляций видов-эдификаторов на территории ботанического памятника природы местного значения «Балка Сухая» (Украина, Донецкая обл.). Выявлены индикаторные особенности эколого-демографической структуры ценопопуляций эдификаторов степных фитоценозов, которые могут отражать состояние растительных сообществ на разных стадиях сукцессий разнотравно-типчаково-ковыльной степи.
Сукцессия, растительность, фитоценоз, ценопопуляции, эдификатор
Короткий адрес: https://sciup.org/148202181
IDR: 148202181 | УДК: 581.55:502.7
Population monitoring of steppe vegetation successions in the «Balka Sukhaya» nature sanctuary of local significance
The paper studies condition of edificatory species’ cenopopulations in the «Balka Sukhaya» nature sanctuary of local significance (Ukraine, Donetsk region). The study revealed some indicator features in ecologic and demographic structure of cenopopulations of steppe phytocenoses’ edificators. These features are indicative of the condition of plant communities that are at different stages of mixed grasses-fescue-feather grass succession.
Текст научной статьи Популяционный мониторинг сукцессий степной растительности на территории ботанического памятника природы местного значения «Балка Сухая»
А нтропогенное влияние на растительный покров приводит к сокращению участков с естественной растительностью и ставит под угрозу существование многих видов растений. Всё более актуальными становятся исследования современного состояния растительного покрова с целью нормирования и лимитирования использования природных ресурсов [3, 4]. Изменения в структурно-функциональной организации ценопопуля-ций являются показателем сукцессионных преобразований фитоценозов. Популяционный мониторинг позволяет не только оценить состояние фитоценозов, но и определить необходимость вмешательства человека в их развитие. Данные, необходимые для оценки состояния экосистемы, можно получить в результате наблюдения за изменениями популяционных параметров доминирующих видов, в частности эдификаторов.
Цель работы – оценить индикаторные возможности ценопопуляций эдификаторов и доминан-тов степных фитоценозов для организации мониторинга сукцессий растительности на территориях природно-заповедного фонда.
Объекты исследования – ценопопуляции Fes-tuca valesiaca Gaudin, Stipa capillata L., S. lessingi-ana Trin. et Rupr., S. joannis L. в степных фитоценозах на разных стадиях сукцессий растительности.
Исследования проводили с 2003 г. на территории ботанического памятника природы местного значения «Балка Сухая» (далее БПП) в Ясиноват-ском р-не Донецкой обл. (Украина) в экотопах, охватывающих разнообразные условия местообитаний, отличающихся растительным и почвенным покровами, режимом использования.
Выделение возрастных групп, изучение возрастной структуры осуществляли в соответствии с общепринятой методикой [1, 6, 8]. Основные типы пространственного распределения особей определяли согласно А.М. Гилярову: использовали отношение дисперсии к среднему: 5— , где о2 m
– дисперсия, m – средняя. Если показатель около единицы, то исследуемое распределение – случайное, если больше – контагиозное, если меньше – регулярное. Виталитетную структуру изучали согласно Ю.А. Злобину [8]. По качеству ценопо- пуляции подразделяли на три основных типа:
процветающие - q = ( a + b ) > с , равновесные
( a + b )
- Q =
с , депрессивные –
q = ( a + b ) < с , где q — индекс качества цено-2
популяции; " a ", " b ", " c ", – соответствующие частоты особей высшего, среднего и низшего классов. Для выявления ключевых признаков был применён факторный анализ. Виталитетное состояние определяли по следующим морфометрическим параметрам: высота растений, диаметр дерновин, количество генеративных побегов. Для определения степени антропогенной трансформации растительности использовали работы А.З. Глухова, О.М. Шевчук [2], В.М. Остапко [10], В.В. Осычнюка [11], В.В. Ткаченко [12]. Стадии сукцессий определяли по состоянию ценопопуля-ций степных эдификаторов, флористическому составу фитоценозов, соотношению видов в них.
БПП относится к малым биоцентрам с характерной степной растительностью [7], его площадь 150 га. Наиболее распространёнными являются настоящие степи, типичные для юго-востока Украины, менее распространены сообщества петро-фитного и кустарникового вариантов степи. На- стоящие разнотравно-типчаково-ковыльные степи представлены на черноземах плакорных участков на смытых черноземах склонов балки. Также встречаются участки с остепнёнными лугами и суходольной луговой растительностью. Есть небольшой участок байрачного леса, участки с болотной, водной и синантропной растительностью. Все исследованные фитоценозы являются в той или иной степени антропогенно трансформированными. Наиболее глубокое деструктивное влияние на растительный покров БПП оказало нерационально организованное пастбищное использование территории.
На территории БПП выявлено 53 ассоциации доминантной классификации степной растительности. Из них 14 ассоциаций подлежат охране – занесены в Зелёную книгу Украины [7]: Stipetum (capillatae) bromopsiosum (ripariae) , S. caraganosum (fruticis) ., S. elytrigiosum (repentis) , S. festucosum (valesiacae) , S. galatellosum (villosae), S. phleosum (tuberosi) , Stipetum (grafianae) festucosum (valesiacae) , Stipetum (joannis) purum., S. festucosum (valesiacae) ., Stipetum (lessingianae) bromopsiosum (ripariae) , S. galatellosum (villosae) , S. festucosum (valesiacae) , S. purum , S. caraganosum (fruticis) , а 3 ассоциации являются регионально редкими: Caraganetum (fruticis) sti-posum (capillatae) , Elytrigietum (repenis) stiposum (capillatae ), Festucetum (valesiacae) stiposum (capillatae) [10]. Все они являются слабо нарушенными и занимают значительную часть территории БПП.
Главным эдификатором сохранившихся фрагментов степей выступает Stipa capillata L. – наиболее стойкий к различным видам антропогенного воздействия (стравливание скотом, вытаптывание, выжигание, скашивание) вид ковыля. Распространёнными являются также фитоценозы, в которых роль этого ковыля является менее выраженной – кустарниковые степи, дигрессивные типчаковые сообщества. Они встречаются на более или менее пологих склонах и, изредка, на узких полосках плакора на чернозёмных, часто эродированных, щебнистых почвах. На территории БПП наиболее распространённой является формация Festuceta valesiacae. Усиление антропогенной нагрузки на растительный покров здесь вызвало значительное сокращение общей площади степных сообществ и увеличение количества участков, непригодных к освоению типчаковыми травостоями (молочайные, груднициевые фитоценозы сбойных участков с участием Poa bulbosa L.). Кустарниковые степные сообщества занимают участки в верхней и средней частях пологих склонов. В качестве доминанта, как правило, выступает Caragana frutex L. Снижение пастбищной нагрузки в последнее время способствовало расширению площади сообществ длиннокорневищных растений на некоторых участках степи, в том числе Elytrigia repens (L.) Nevski, Poa angustifolia L., Bromopsis inermis (Leys.) Holub., то есть плот-нодерновинные степные ценозообразователи на таких участках практически вытеснены из сообществ.
Многие из участков с остатками степной растительности в настоящее время используются как пастбища для крупного рогатого скота. Чрезмерные пастбищные нагрузки привели к формированию довольно однообразных по составу и физиономическому облику дигрессивных сбоев. Кроме того, в БПП отмечено наличие залежей, где формируются растительные сообщества с доминированием сорных видов растений, находящиеся на разных стадиях демутационной сукцессии.
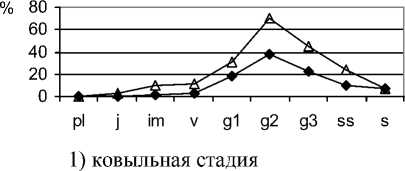
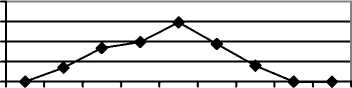
На стадии умеренного выпаса (ковыльной) це-нопопуляции эдификаторов степных фитоценозов в большинстве своём полночленные, нормальные, зрелые (рис.1).
A Stipa capillata L.
-♦ Festuca valesiaca Gaudin.
2) типчаковая стадия
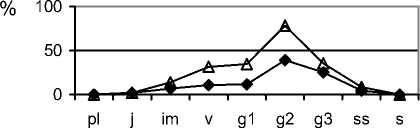
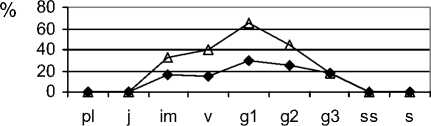
Рис. 1. Возрастные спектры ценопопуляций эдификаторов степных фитоценозов на разных стадиях пастбищной дигрессии в памятнике природы «Балка Сухая»
3) луковичномятликовая стадия
Онтогенетические группы субсенильных и сенильных растений малочисленные. Ценопопуля-ции характеризуются разнообразием возрастного состава, что обеспечивает не только их устойчивое положение в фитоценозах, но и длительность существования самих растительных сообществ. Молодых вегетативных растений немного, что связано с их быстрым онтогенетическим развитием в благоприятных условиях произрастания. Большая часть ценопопуляций относится к процветающему виталитетному типу, характеризуется высокой плотностью, контагиозным размещением особей в пространстве (табл. 1).
Таблица 1. Виталитетная структура, плотность и тип пространственного размещения особей ценопопуля-ций эдификаторов степных фитоценозов на разных стадиях пастибщной дигрессии в памятнике природы «Балка Сухая»
|
Стадия |
Вид |
Виталитет ный спектр |
Плотность, особей/м2 |
2 σ m |
|||
|
c |
b |
a |
Q |
||||
|
ковыльная |
Festuca valesiaca |
0,340 |
0,340 |
0,320 |
0,330 |
6,1 ± 1,5 |
0,93 |
|
Stipa capillata |
0,200 |
0,600 |
0,200 |
0,300 |
3,5 ± 0,6 |
0,8 |
|
|
типчаковая |
Festuca valesiaca |
0,320 |
0,320 |
0,360 |
0,340 |
14,6 ± 2,4 |
0,97 |
|
Stipa capillata |
0,360 |
0,320 |
0,320 |
0,320 |
4,3 ± 1,1 |
1,8 |
|
|
луковичномятликовая |
Festuca valesiaca |
0,440 |
0,320 |
0,240 |
0,280 |
2,0 ± 0,8 |
2,1 |
|
Stipa capillata |
0,600 |
0,280 |
0,120 |
0,200 |
1,3 ± 0,6 |
2,5 |
|
|
сбой |
Festuca valesiaca |
угнетенные особи |
единичные особи |
||||
|
Stipa capillata |
угнетенные особи |
единичные особи |
|||||
При усилении пастбищной нагрузки из состава фитоценозов в первую очередь исчезают виды рода Stipa. На стадии сильного выпаса (типчаковой) ценопопуляции S. capillata неполночленные, нормальные, зрелые (рис. 1). Общей чертой для них является не только уменьшение доли особей постгенеративных групп, но и увеличение количества молодых и средневозрастных генеративных особей, что связано с замедлением онтогенетического развития [5]. Уменьшается количество молодых вегетативных растений из-за нерегулярности семенного возобновления. Особи в пространстве размещены контагиозно. Ценопопуля-ции депрессивные, их плотность постепенно снижается. Ценопопуляции F. valesiaca – полночленные, нормальные, зрелые (рис. 1). Для них характерным является некоторое омоложение в результате активизации возобновления, что может быть следствием уменьшения конкурентного влияния со стороны видов рода Stipa и появления свободного пространства для поселения новых растений. Ценопопуляции, в большинтве случаев, являются процветающими. Отмечено заметное увеличение плотности ценопопуляций этого вида . Ценопопу-ляции эдификаторов степных фитоценозов на стадии чрезмерного выпаса (луковичномятликовая) неполночленные, «ложномолодые» (рис. 1), относятся к депрессивному виталитетному типу. Их плотность низкая, особи по площади размещены контагиозно (табл. 1). В возрастном составе ценопопуляций эдификаторов степных фитоценозов отсутствуют субсенильные, сенильные, часто старые генеративные растения, существенно увеличивается доля молодых вегетативных растений. Подобные изменения отмечены и в возрастной структуре ценопопуляций S. lessingiana, S.
joannis. Во всех исследованных ценопопуляциях видов рода Stipa сильно сокращается плотность особей, в меньшей степени она снижена у S. capil-lata. Ценопопуляции F. valesiaca более устойчивы и подобные изменения в возрастной структуре происходят с меньшей скоростью. Плотность ценопопуляций выше, чем у ценопопуляций S. capillata. Исследованные ценопопуляции депрессивные. Фитоценозы, основу которых составляют ценопопуляции с таким неполным возрастным составом, являются уже сильно нарушенными и их быстрое восстановление без вмешательства человека уже невозможно. Необходимо не только существенно снизить антропогенную нагрузку или, если фитоценозы являются сильно трансформированными, вообще её прекратить на несколько лет с последующим установлением на данной территории режима умеренной нагрузки, но и использовать активные методы восстановления растительного покрова, в том числе, и реинтродукцию. В сообществах на четвёртой стадии пастбищной дигрессии (сбой) отмечены сильно угнетённые единичные, в основном, зрелые генеративные особи F. valesiaca. Главным фактором, определяющим будущее этих ценопопуляций, является семенное размножение. Поэтому почти полное исчезновение из их возрастного состава особей генеративных групп (молодые, зрелые и старые генеративные растения), ставит под угрозу их жизнеспособность. На этой стадии пастбищной дигрессии они распадаются, изменения в ценопопуляциях и фитоценозах становятся необратимыми. В этом случае необходимо использовать методы натурного моделирования искусственных фитоценозов для быстрого восстановле- ния сильно нарушенного растительного покрова [9].
Таким образом, глубокие изменения, приводящие к нарушению структурно-функциональной организации ценопопуляций, при сильных антропогенных нагрузках происходят на последних стадиях дигрессии. Существенные изменения в эколого-демографической структуре могут привести к катастрофическим преобразованиям не только организации самих ценопопуляций отдельных эдификаторов степных фитоценозов, но и растительных сообществ в целом. Подобные изменения сопровождаются сдвигами в соотношении видов, что приводит к смене типов расти-
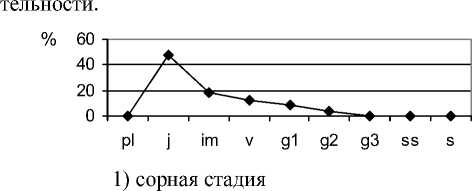
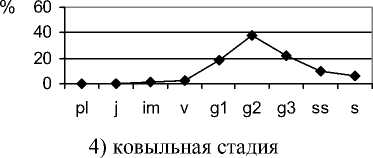
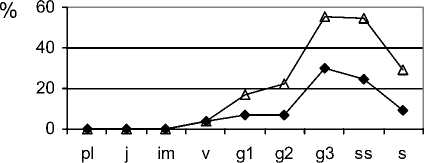
Снижение антропогенной нагрузки приводит к восстановлению структурно-функциональной организации ценопопуляций плотнодерновинных ценозообразователей и стабилизации фитоценозов. В связи с этим целесообразно использовать для определения состояния фитоценозов маркирование стадий демутационной сукцессии степной растительности особенностями экологодемографической структуры ценопопуляций F. valesiaca , так как этот вид присутствует в составе фитоценозов почти на всех стадиях сукцессии. В растительных сообществах на сорной стадии це-нопопуляции F. valesiaca «ложномолодые», подобны неполночленным нормальным молодым (рис. 2).
%

pl j im v g1 g2 g3 ss s
-
2) стадия длиннокорневищных злаков
%
pl j im v g1 g2 g3 ss s
-
3) типчаковая стадия
Рис. 2. Возрастные спектры ценопопуляций Festuca valesiaca Gaudin в фитоценозах на разных стадиях демутационной сукцессии
В их возрастном составе отсутствуют проростки, ювенильные, имматурные, старые генеративные, субсенильные и сенильные растения. Молодые и средневозрастные генеративные особи немногочисленны. К концу этой стадии в возрастном спектре отмечены молодые особи всех онтогенетических групп левой части возрастного спектра ценопопуляций, максимум приходится на группу виргинильных растений. В виталитетном составе ценопопуляций этого вида большинство растений являются угнетёнными. Но к завершению сорной стадии в ценопопуляциях преобладают особи с высоким жизненным уровнем. Це-нопопуляции, как правило, становятся процветающими. Плотность F. valesiaca на этой стадии очень низкая, растения в пространстве размещены контагиозно (табл. 2).
Таблица 2. Виталитетная структура, плотность и тип пространственного размещения особей ценопопуляций Festuca valesiaca Gaudin в фитоценозах на различных стадиях демутации в памятнике природы «Балка Сухая»
|
Стадия демутационной сукцесии |
Виталитетный спектр |
2 σ m |
Плотность, особей/м2 |
|||
|
c |
b |
a |
Q |
|||
|
сорная |
угнетённые особи |
2,7 |
1,4 ± 0,8 |
|||
|
длиннокорневищных злаков |
0,240 |
0,440 |
0,320 |
0,380 |
3,1 |
2,9 ± 1,0 |
|
дерновинных злаков |
0,280 |
0,320 |
0,360 |
0,340 |
1,9 |
7,8 ± 1,5 |
|
вторичная целина |
0,320 |
0,240 |
0,440 |
0,340 |
1,2 |
5,7 ± 0,9 |
В растительных сообществах на стадии корневищных злаков демутационной сукцессии цено-популяции F. valesiaca являются молодыми нормальными неполночленными (нет старой генера- тивной, субсенильной и сенильной онтогенетических групп растений). Максимум в возрастном спектре ценопопуляций приходится на возрастные группы имматурных, реже виргинильных особей. К концу этой стадии в возрастном составе ценопопуляций возрастает доля молодых генеративных растений и, в меньшей степени, зрелых, значительно увеличивается плотность ценопопуляций в целом. Ценопопуляции процветающие. Формируется пространственная структура контагиозного типа. Переходу на стадию дерновинных злаков способствует регулярный умеренный выпас, который стимулирует ксерофитизацию растительных сообществ. Это, в свою очередь, способствует ускорению вытеснения из состава фитоценозов мезофитных корневищных видов и появлению свободного пространства, которое быстро занимают особи степных эдификаторов. Формируются эколого-фитоценотические условия для внедрения видов степного разнотравья и развития их ценопопуляций. Ценопопуляции F. valesiaca в фитоценозах на стадии плотнодерно-винных злаков (типчаковой) молодые, нормальные, неполночленные (отсутствуют субсенильные и сенильные растения) (рис. 2). В возрастном составе преобладают молодые вегетативные растения, увеличивается доля молодых и зрелых генеративных особей. Принадлежность большинства растений к первому виталитетному классу также способствует упрочнению положения F. valesiaca, так как именно эти растения ответственны за функцию семенного возобновления и самоподдержание популяционных систем. Отмечено существенное увеличение плотности цено-популяций данного вида (табл. 2). В конце этой стадии ценопопуляции являются нормальными, зрелыми, полночленными. Особи в пространстве расположены контагиозно. За счёт семенного возобновления не только увеличилась плотность растений, но и сократилось расстояние между их скоплениями, что в будущем может привести к смыканию популяционного поля.
При условии отсутствия регулируемого выпаса, формирование вторичной целины может затягиваться на неопределённо долгий период времени (а возможно, и вообще не осуществиться) [11]. Это может послужить причиной для длительного пребывания некоторых ценозов на стадии длиннокорневищных злаков. Полное исключение антропогенной нагрузки на растительный покров может привести не к восстановлению степных сообществ с доминированием ксерофитных плот-нодерновинных злаков, а к образованию фитоценозов, подобных тем, что формируются на последних стадиях резерватогенной сукцессии с доминированием длиннокорневищных злаков. Осуществляется «оборот», «возвращение» от стадии дерновинных злаков или от конечного этапа стадии корневищных злаков к периоду восстановления растительных сообществ, который соответствует середине стадии длиннокорневищных злаков. Возможен вариант восстановления растительного покрова, минуя стадию не только вто- ричной целины, но и дерновинных злаков. Впоследствии в такие сообщества внедряются кустарники, что может привести к формированию кустарниковой степи. Ценопопуляции плотно-дерновинных злаков начинают стареть, нарушается цикличность возобновления. Поэтому с целью более быстрого восстановления степных экосистем, возможно, следует устанавливать режим умеренного выпаса уже на начальных этапах пырейной стадии демутационной сукцессии. В противном случае возможно формирование фитоценозов лугово-степного и лугового типов. Прекращение антропогенного воздействия, введение абсолютно заповедного режима в течение длительного времени вызывают изменения растительного покрова резерватогенного типа . Это приводит к формированию «нетипичных» корневищных лугово-степных и луговых фитоценозов [11]. Это является подтверждением того, что для сохранения ксерофитных степных растительных сообществ, в которых доминируют плотнодерно-винные злаки, необходима разработка эффективного комплекса антропогенных нагрузок на растительный покров (умеренный выпас, палы в чётко установленное время и т.д.).
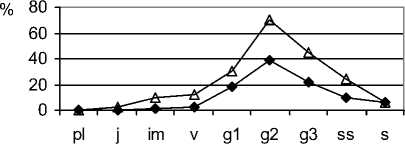
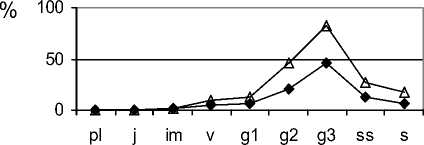
Для определения состояния фитоценозов, прогнозирования дальнейшего их развития, маркирования конкретных стадий резерватогенной сукцессии степной растительности эффективно использовать особенности экологодемографической структуры ценопопуляций F. valesiaca , S. capillata , которые присутствуют в составе фитоценозов почти на всех стадиях. Такие ковыли, как S. lessingiana и S. joannis часто исчезают из фитоценозов на корневищнозлаковой стадии, особенно чувствителен к изменениям среды S. lessingiana. Ценопопуляции F. valesiaca в фитоценозах на первой стадии резерватогенной сукцессии относятся, в основном, к нормальным полночленным зрелым или стареющим (рис. 3). Большинство их являются процветающими. Их плотность достаточно высокая. Пространственная структура случайного типа, иногда (при ухудшении условий) - контагиозного (табл. 3).
Подобные характеристики наблюдаются и у ценопопуляций не только Stipa capillata (рис. 3) , но и других эдификаторов - S. lessingiana, S. joannis . В растительных сообществах на этой стадии резерватогенной сукцессии «ведущие це-нопопуляции», к которым относятся виды родов Festuca и Stipa , сбалансированы по структуре и темпам обновления.
Таким образом, растительные сообщества на ковыльной стадии резерватогенной сукцессии при умеренной регулируемой антропогенной нагрузке являются устойчивыми, способными к саморегуляции и сохранению состава и структуры. Этому способствует то, что составляющие их ценопопу- ляции видов-эдификаторов, образуя среду обитания для сопутствующих видов и определяя состав и структуру их ценопопуляций, формируют устойчивую основу степных ценозов.
A Stipa capillata L.
-
-♦ Festuca valesiaca Gaudin.
-
1) ковыльная стадия
2) стадия
% 100
% 100
3) корневищно-злаковая
-
4) корневищно-злаковоразнотравная стадия
-
5) кустарниковая стадия
Рис. 3. Ценопопуляции эдификаторов степных фитоценозов на разных стадиях резерватогенной сукцессии в памятнике природы «Балка Сухая»
Таблица 3. Плотность и тип пространственного размещения особей ценопопуляций эдификаторов степных фитоценозов на различных стадиях резерватогенной сукцессии в памятнике природы «Балка Сухая»
|
Стадия |
Вид |
Виталитетный спектр |
Плотность, особей / м2 |
2 σ m |
|||
|
c |
b |
a |
Q |
4,4 ± 0,8 |
0,98 |
||
|
ковыльная |
Festuca valesiaca |
0,320 |
0,280 |
0,400 |
0,340 |
||
|
Stipa capillata |
0,280 |
0,320 |
0,360 |
0,340 |
5,8 ± 0,6 |
0,97 |
|
|
типчаковая |
Festuca valesiaca |
0,280 |
0,360 |
0,360 |
0,360 |
6,1 ± 1,3 |
1,1 |
|
Stipa capillata |
0,300 |
0,350 |
0,350 |
0,350 |
3,5 ± 1,1 |
1,3 |
|
|
корневищнозлаковая |
Festuca valesiaca |
0,440 |
0,360 |
0,200 |
0,280 |
3,0 ± 1,2 |
1,4 |
|
Stipa capillata |
0,440 |
0,200 |
0,360 |
0,280 |
2,8 ± 1,1 |
2,5 |
|
|
злаковоразнотравная |
Festuca valesiaca |
0,500 |
0,300 |
0,200 |
0,250 |
2,2 ± 1,4 |
2,1 |
|
Stipa capillata |
0,480 |
0,360 |
0,160 |
0,260 |
2,0 ± 1,0 |
2,9 |
|
|
кустарниковая |
Festuca valesiaca |
угнетённые особи |
единичные особи |
||||
|
Stipa capillata |
угнетённые особи |
единичные особи |
|||||
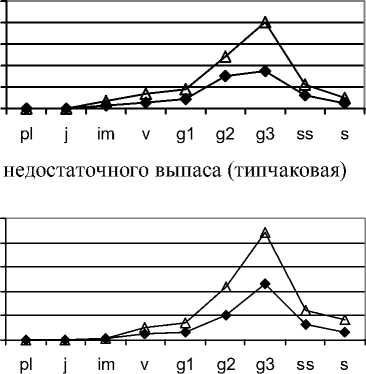
В растительных сообществах на стадии недостаточного выпаса ценопопуляции эдификаторов являются, неполночленными нормальными стареющими. Отмечено снижение плотности их це-нопопуляций. При этом плотность F. valesiaca выше, чем видов рода Stipa, ценопопуляции которых относятся, как правило, к депрессивному типу. При недостаточной антропогенной нагрузке в этих сообществах происходит постепенная за- мена доминантов, возрастает фитоценотическая роль мезофитных видов. Это можно считать начальным звеном резерватогенных преобразований степного растительного покрова: дерновинные злаки замещаются корневищными видами. Особенно к таким изменениям чувствительными являются виды рода Stipa, которые одни из первых значительно сокращают плотность своих ценопо- пуляций. Постепенно доминирующее положение занимает Festuca valesiaca.
Подобные характеристики наблюдаются и у ценопопуляций не только Stipa capillata (рис. 3) , но и других эдификаторов - S. lessingiana, S. joannis. В растительных сообществах на этой стадии резерватогенной сукцессии «ведущие ценопопуляции», к которым относятся виды родов Festuca и Stipa , сбалансированы по структуре и темпам обновления.
Таким образом, растительные сообщества на ковыльной стадии резерватогенной сукцессии при умеренной регулируемой антропогенной нагрузке являются устойчивыми, способными к саморегуляции и сохранению состава и структуры. Этому способствует то, что составляющие их ценопопу-ляции видов-эдификаторов, образуя среду обитания для сопутствующих видов и определяя состав и структуру их ценопопуляций, формируют устойчивую основу степных ценозов .
В растительных сообществах на стадии недостаточного выпаса ценопопуляции эдификаторов являются неполночленными, нормальними, стареющими. Отмечено снижение плотности их це-нопопуляций. При этом плотность F. valesiaca выше, чем видов рода Stipa , ценопопуляции которых относятся, как правило, к депрессивному типу. При недостаточной антропогенной нагрузке в этих сообществах происходит постепенная замена доминантов, возрастает фитоценотическая роль мезофитных видов. Это можно считать начальным звеном резерватогенных преобразований степного растительного покрова: дерновинные злаки замещаются корневищными видами, Особенно к таким изменениям чувствительными являются виды рода Stipa , которые одни из первых значительно сокращают плотность своих ценопо-пуляций. Постепенно доминирующее положение занимает Festuca valesiaca .
Ценопопуляции эдификаторов степных фитоценозов на корневищнозлаковой стадии резерватогенной сукцессии уже не играют значительной роли; в лучшем случае, у них может сохраняться разнообразие возрастного состава, но плотность будет низкой. Возрастной спектр большинства исследованных ценопопуляций неполный, отсутствуют молодые вегетативные растения, что говорит о длительном перерыве в семенном возобновлении ценопопуляций и эффективном закреплении проростков в фитоценозах. Это свидетельствует о неблагоприятных условиях для эффективной инспермации из-за избыточного накопления мортмассы, которая мешает прорастанию и достижению семенами почвы, а также из-за развития ценопопуляций длиннокорневищных видов. Преобладают старые генеративные растения, существенно увеличивается доля субсенильных и сенильных особей. Особи размещены контагиозно, что свидетельствует об ухудшении условий существования. Именно такой тип размещения способствует выживанию особей, увеличивает способность к сопротивлению стрессу. Ценопо-пуляции эдификаторов степных фитоценозов депрессивные. Смещение максимума в возрастных спектрах ценопопуляций данных видов на более старые группы растений является неоспоримым свидетельством отрицательных тенденций в изменении жизнеспособности ценопопуляций плотнодерновинных злаков, не способных к поддержанию стабильности базовых параметров структуры популяции за счёт активизации вегетативного размножения. В растительных сообществах на данной стадии развития существенную роль начинают играть «дополняющие ценопопуляции» (ассектаторы, вегетативноподвижные виды, которые в типичных степных фитоценозах не обладают высоким обилием) [5].
В растительных сообществах на корневищно-злаково-разнотравной стадии ценопопуляции эдификаторов, как правило, старые неполночленные, депрессивные. В их возрастном составе присутствует небольшое количество зрелых и старых генеративных особей, изредка есть молодые растения. Плотность особей очень низкая, малочисленные группы особей расположены на значительном расстоянии друг от друга, вследствии чего ценопопуляции степных видов теряют способность к самоподдержанию, что ставит их на грань гибели. Они постепенно уступают место другим видам, ценопопуляции которых формируют «нехарактерные» для степи сообщества. В дальнейшем возможно полное исключение данных эди-фикаторов и полное доминирование длиннокорневищных злаков, а среди разнотравья - увеличение роли ксеромезофитных, мезоксерофитных и мезофитных видов. Из-за существенных деструктивных изменений в структуре ценопопуляций видов, которые составляют основу фитоценозов , и тем самым определяют их организацию и внешний вид, происходит нарушение структурнофункциональной организации экосистем, их замена нетипичными для степной зоны растительными сообществами.
На кустарниковой стадии резерватогенной сукцессии степной растительности ценопопуля-ции F. valesiaca и S. capillata относятся к старым нормальным неполночленным (отсутствуют многие или все ранние группы молодых вегетативных растений), они находятся на стадии перехода в регрессивные или уже являются регрессивными . Численность очень низкая (единичные особи размещены контагиозно). S. lessingiana и S. joannis на этой стадии резерватогенной сукцессии отсутствуют.
При мезофитизации растительного покрова усиливается ценотическая роль степных кустарников, что приводит к деструктивным резерватогенным преобразованиям в степных сообществах:
сокращаются площади дерновиннозлаковых сообществ, увеличиваются площади корневищнозлаковых и разнотравных ценозов, происходит замена доминирующих ксерофитных видов, в том числе эдификаторов, на степные кустарники. Помимо Caragana frutex (L.) K. Koch и Amygdalus nana L., которые формируют основу кустарниковой степи, заметно усиливается участие Prunus stepposa Kotov и видов рода Rosa L.
Таким образом, мониторинговые исследования в БПП «Балка Сухая» позволили выявить изменения популяционных параметров, отражающие процессы трансформации степных растительных сообществ. Полученные данные могут иметь индикаторное значение для определения стадий резерватогенной сукцессии разнотравно-типчаково-ковыльных степей на смытых чернозёмах. Для каждого типа сукцессии степной растительности установлены виды-эдификаторы, особенности эколого-демографической структуры которых целесообразно использовать: состояние структуры ценопопуляций Festuca valesiaca, Stipa capillata , как биомаркер стадий пастбищной дигрессии; F. valesiaca – демутации, F. valesiaca , S. capillata – резерватогенной сукцессии степного растительного покрова.
Список литературы Популяционный мониторинг сукцессий степной растительности на территории ботанического памятника природы местного значения «Балка Сухая»
- Гиляров А.М. Популяционная экология. М.: Изд-во Москов. гос. ун-та, 1990. 191 с.
- Глухов О.З., Шевчук О.М., Кохан Т.П. Науковi основи вiдновлення трав’яних фiтоценозiв в степовiй зонi України. Донецьк: Вид-во «Вебер», 2008. 198 с.
- Гнатюк Н.Ю. К изучению флоры бассейна реки Крынки (бассейн реки Миус)//Проблеми екологiї та охорони природи техногенного регiону. 2010. № 1. С. 47 -56.
- Емельянов И.Г., Емельянова Л.В., Песков В.Н. Популяция как объект экологического мониторинга//Матер. мiжнарод. наук. конф.: Заповiднi степи України. Стан та перспективи їх збереження. 2007. С. 49 -51.
- Жиляев Г.Г. Жизнеспособность популяций растений. Львов: Б.и., 2005. 304 с.
- Заугольнова Л.Б., Денисова Л.Б., Никитина С.В. Подходы к оценке состояния ценопопуляций растений//Бюл. МОИП. Отд. биол. 1993. 98, вып. 5. С. 100 -109.
- Зеленая книга Украинской ССР: Редкие, исчезающие и типичные, нуждающиеся в охране растительные сообщества/[под общ. ред. Ю.Р. Шеляга-Сосонко]. Киев: Наук. думка, 1987. 216 с.
- Злобин Ю.А. Принципы и методы изучения ценотических популяций растений. Казань: Изд-во Казан. ун-та, 1989. 146 с.
- Кондратюк Е.Н., Чуприна Т.Т. Ковыльные степи Донбасса. Киев: Наук. думка, 1992. 172 с.
- Остапко В.М. Эйдологические, популяционные и ценотические основы фитосозологии на юго-востоке Украины. Донецк: ООО «Лебедь», 2005. 408 с.
- Осичнюк В.В. Змiни рослинного покриву степу//Рослиннiсть УРСР. Степи, кам’янiстi вiдслонення, пiски. К.: Наук. думка, 1973. С. 249 -315.
- Ткаченко В.С. Фiтоценотичний монiторинг резерватних сукцесiй в Українському степовому природному заповiднику. К.: Фiтосоцiоцентр, 2004. 184 с.
