Последовательность активации миокарда желудочка атлантической трески (Gadus morhua marisalbi)
")
Автор: Вайкшнорайте М.А., Витязев В.А., Азаров Я.Э.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 4 (36), 2018 года.
Бесплатный доступ
Проведены электрофизиологические исследования с целью установления последовательности деполяризации интрамуральных слоев миокарда желудочка сердца атлантической трески (Gadus morhua marisalbi, n=5) при температуре 12°С. Частота сердечных сокращений сердца трески - 34±5 уд./мин. Первые очаги активации миокарда появляются в субэндокардиальных слоях возле атриовентрикулярного отверстия, а поздние - в субэпикардиальных слоях верхушки. Волна возбуждения в миокарде желудочка трески движется от зоны атриовентрикулярного отверстия к области верхушки (p
Активация, апикобазальный градиент, трансмуральный градиент, сердце, атлантическая треска
Короткий адрес: https://sciup.org/149128807
IDR: 149128807 | УДК: 612.171.1 | DOI: 10.19110/1994-5655-2018-4-31-35
The sequence of activation of ventricular myocardium in Atlantic cod (Gadus morhua marisalbi)
Electrophysiological studies were carried out to establish the sequence of depolari- zation of intramural layers of ventricular myocardium of the Atlantic cod ( Gadus morhua marisalbi, n = 5) at 12°C. The heart rate of cod is 34 ± 5 bpm. The first foci of myocardial activation appear in the subendocardial layers near the atrioven- tricular orifice, and the later - in the subepicardial layers of the apex. The excita- tion wave in the myocardium of cod ventricle moves from the zone of atrioventricu- lar orifice to the region of the apex (p
Текст научной статьи Последовательность активации миокарда желудочка атлантической трески (Gadus morhua marisalbi)
Для рыб вследствие слабого развития проводящей системы [1–3] характерен «последовательный» тип активации миокарда – c медленным распространением волны возбуждения в желудочке сердца [4]. В исследованиях на рыбах-хищниках – щука (Esox lucius), хариус (Thymallus thymallus) – установлено, что ранние очаги активации находятся в субэндокардиальных слоях задней стенки основания желудочка, а поздние – в субэпикардиальных слоях верхушки и передней стенки основания сердца; движение волны активации в большинстве зон миокарда – вдоль стенок желудочка от основания к верхушке [4]. Известно, что на активацию миокарда влияет не только организация желудочковой проводящей системы, но и строение миокардиального слоя стенки желудочка. Показано, что рыбы в связи с адаптацией к разному образу жизни (активные и малоактивные) имеют: 1) разную форму желудочка сердца (цилиндрическую, мешкооб- разную и пирамидальную) [5]; 2) разную толщину компактного слоя миокарда и тип коронарного кровоснабжения [6–8].Типы макроскопической организации желудочка коррелируют с двигательной активностью рыб. Пирамидальные желудочки характерны для рыб с активным образом жизни, относительно высоким сердечным выбросом и стенкой желудочка смешанного строения, состоящей из губчатого и компактного миокарда. Стенка желудочка с мешкообразной и цилиндрической формой, как правило, имеет только губчатый миокард и встречается у костистых малоподвижных рыб [9, 10]. Связь формы желудочка и строения его стенки с последовательностью его электрической активации мало изучена. Однако к настоящему времени исследованы некоторые закономерности активации желудочка разных рыб. Установлено, что эпикардиально-эндокардиальный (трансмуральный) градиент во времени активации миокарда у щук, как типичного представителя «активных» рыб, присутствует только в области, вблизи атриовентрикуляр- ного (АВ) соединения желудочка [11, 12]. У карпа, ведущего «малоактивный образ жизни», имеющего пирамидальный желудочек, но с крайне слаборазвитым слоем компактного миокарда [5], трансмуральный градиент времени активации существует и в области верхушки, и в области основания желудочка [13]. Показано, что длительность потенциала действия в желудочке малоподвижного карпа больше, чем у щуки или форели [14]. Таким образом, при сходных типах макроскопической организации желудочка рыб возможны различные варианты паттерна активации.
Известно, что треска ведет донный образ жизни, в то же время предпринимает длительные сезонные миграции, заставляющие стаи рыб преодолевать расстояния до 1,5 тыс. км от нерестилищ до мест откорма. Известно, что желудочек сердца «активной» трески, как у щуки и карпа, имеет пирамидальную форму. Однако не изучены особенности активации желудочка у этого вида рыб.
Целью исследования явилось установление последовательности деполяризации в интрамуральных слоях желудочка сердца атлантической трески.
Материал и методы
Работа проводилась в сентябре на Беломорской биостанции им. Н.А.Перцова биофака МГУ им. М.В.Ломоносова. Электрофизиологические исследования проведены на сердце атлантической трески шт situ ( Gadus morhua marisalbi, n=5, масса 310±145 гр.) при температуре 12°С, что является физиологической нормой для животных в данное время года, так как соответствует температуре акклиматизации [15]. Для проведения экспериментов животных обездвиживали, жизнедеятельность поддерживали, осуществляя искусственную перфузию жабр морской водой.
Регистрация электрофизиологических показателей производилась с помощью 128-канальной компьютерной картографической установки (полоса пропускания 0,05–1000 Гц, частота дискретизации 4000 Гц, динамический диапазон входных сигналов от ±10 мВ до ±100 мВ, уровень шума не более ±10 мкВ, разрешающая способность от 10 до 100 мкВ на один разряд аналого-цифрового преобразования). Регистрацию интрамуральных электрограмм (ЭГ) желудочка сердца трески осуществляли при помощи трех интрамуральных игольчатых электродов (далее «игла») [16]. В качестве индифферентного электрода использовали объединенный электрод по Вильсону от трех удаленных от сердца точек на поверхности тела. Иглы вводили перпендикулярно стенке желудочка (рис.1, А, Б): в область «основания» желудочка, которой считали зону соединения желудочка с луковицей аорты (игла №1); в область вблизи атриовентрикулярного канала (игла №2); в апикальную область («верхушка») желудочка (игла №3). Каждая интрамуральная игла состояла из металлического основания, на который были намотаны изолированные (ПЭВ-2) медные провода (0.07мм). На каждой игле располагались восемь регистрирующих макроэлектродов, рав-
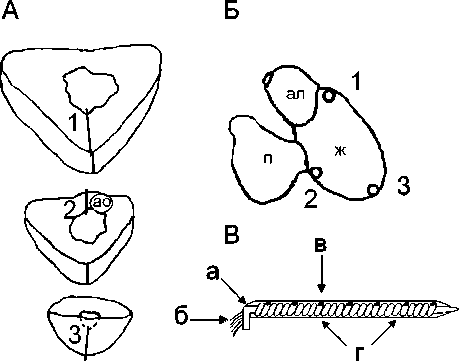
Рис.1. Схема расположения интрамуральных игл в толще миокарда (А) и на эпикарде желудочка (Б); схема строения интрамурального игольчатого электрода (В) [3].
-
1, 2, 3 – номера иголок, ао – атриовентрикулярное отверстие, п – предсердие, ж – желудочек, ал – артериальная луковица, а – металлическая основа (стальная проволка, 0,17–0,20мм), б – микропровода (изолированные (ПЭВ-2) медные провода), в – электродная бляшка, г – эпоксидная смола.
Fig.1. Scheme of intramural needles location in the myocardium thickness (A) and on the ventricular epicardium (Б). Scheme of intramural needle electrode (B) [3].
-
1, 2, 3 – numbers of needles, ao – atrioventricular orifice, п – atrium, ж – ventricle, aл – arterial bulb, a – metal base (steel wire, 0.17-0.20 mm); б – microwires (insulated (PEV-2) copper wires), в – electrode plaque, г – epoxy resin.
номерно распределенных по ее длине (рис. 1, В). Длина интрамуральной иглы зависела от толщины стенки желудочка трески (от 3,5 до 4 мм). Субэпикардиальные электрограммы регистрировали от двух первых электродов на игле, а субэндокардиальные – от двух последних электродов, остальные четыре служили для отведения интрамиокардиаль-ных ЭГ (рис. 1, В). Одновременно с желудочковыми электрограммами регистрировали ЭКГ от поверхности тела в отведениях, соответствующих отведениям от конечностей человека.
Время деполяризации (момент прихода волны возбуждения в точку регистрации) определяли по времени минимума первой производной потенциала по времени в период комплекса QRS униполярной ЭГ [11]. Глобальную дисперсию активации рассчитывали как разность между максимальным и минимальным значением времени деполяризации из всех зарегистрированных желудочковых ЭГ в желудочке. Трансмуральный градиент активации определяли как интервал времени между моментами активации субэндокарда и субэпикарда, апико-базальный градиент – как интервал времени между моментами активации А-В соединения и моментом активации верхушки желудочка.
Статистическая обработка данных проводилась с помощью программ BIOSTAT 4.03, SPSS. Для сравнения значений использовали непарамет- рический критерий Уилкоксона, критерий Фридмана с поправкой Ньюмена-Кейсла, различия считали значимыми при p<0,05.
Результаты и обсуждение
В наших экспериментах частота сердечных сокращений у трески составляла 34±5 уд./мин (n=5) при температуре воды 12°С. Длительности электрокардиографических интервалов (медиана и интерквартильный интервал, IQR): QRS комплекс - 52 (IQR 49-53) мс, QT- 574 (IQR 536-602) мс Продолжительность комплексов QRS у разных видов рыб неодинакова, что отражает различное время охвата возбуждением желудочка вследствие различий в размерах сердца: у хариусов Thymallus thymallus – 71 мс, у щук Esox lucius – 94 мс [4], у рыб Danio rerio – 14 [3], у карпа Сyprinus carpio – 79 (72; 85) мс [13].
Начальный желудочковый комплекс электрокардиограммы (ЭКГ) у трески в отведении по продольной оси сердца представлен положительным зубцом R (рис. 2, А), также как и у щук [12] и окуней Perca fluviatilis [17], но не у карпа. Начальная желудочковая активность у карпа в отведении голова-хвост представлена отрицательным зубцом QS [13].
1 2 3
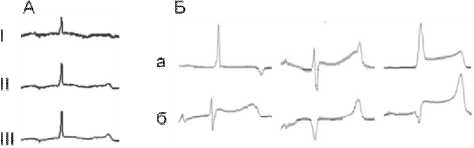
Рис. 2. Репрезентативная электрокардиограмма трески. A – (I, II, III – отведения), Б – репрезентативные электрограммы миокарда желудочка сердца трески, субэпикард (а), субэндокард (б); 1, 2, 3 – номера иголок (см. рис. 1).
Fig. 2. Representative cod electrocardiogram. A – (I, II, III – leads), Б – representative electrograms of ventricular myocardium in cod, subepicardium (a), subendocardium (б); 1,2, 3 – numbers of needles (see Fig 1.).
Вблизи АВ-соединения (рис. 2, Б, позиция 2) внеклеточные потенциалы представлены отрицательными комплексами, а в зонах позднего возбуждения – положительными (рис. 2, Б, позиции 1 и 3). Глобальная дисперсия активации составляет 39 (34; 42) мс. Первые очаги активации желудочка сердца трески, как и у ранее исследованных рыб [4, 11, 12, 13, 18] появляются в субэндокардиальных слоях возле атриовентрикулярного канала, а поздние – в субэпикардиальных слоях верхушки желудочка. Волна активации в миокарде желудочка трески движется от зоны АВ-соединения к верхушке и основанию (аортальному отверстию) желудочка, и от эндокарда к эпикарду. На эпикарде желудочка как между зоной вокруг атриовентрикулярного канала и верхушкой желудочка, так и между зоной основания и верхушкой существует значимая разница во времени возбуждения (см. таблицу). Во всех областях миокарда желудочка присутствует зна-
Длительность активации миокарда желудочка трески при температуре 12°С, Me (25%; 75%), мс
The duration of activation of the ventricular myocardium of cod at a temperature of 12oC (Me (25%; 75%), ms)
Примечание: сравнение между значениями в области соединения желудочка с луковицей аорты и значениями в области атриовентрикулярного отверстия (*) и со значениями в области верхушки (#) – р<0.05, нз – незначимые различия.
Note: The comparison between the values in the junction of the ventricle and the bulb of aorta and the values in the area of atrioventricular orifice (*) and with the values in the apex area (#), p <0.05, нз – nonsignificant differences.
чимый трансмуральный градиент активации, медиана которого составляет 13 (IQR 12-15) мс. Следовательно, у трески, как и у карпа [13], трансмуральный градиент времени активации миокарда существует во всех исследованных областях миокарда, в отличие от щук, у которых он обнаружен только в области АВ-соединения [11, 12]. Показано, что наличие трансмурального градиента у карпа может быть связано с особенностями строения стенки желудочка [19]. В совокупности данные, полученные в настоящем исследовании и предыдущих работах, показывают, что наличие трансмурального градиента активации в большей части желудочка ассоциировано с малоподвижным образом жизни рыбы. Исследование строения стенки желудочка сердца трески могло бы прояснить механизмы и роль градиентов активации в желудочке рыб.
Заключение
Таким образом, показано, что последовательность распространения волны возбуждения в миокарде желудочка трески характеризуется общим направлением от атриовентрикулярного отверстия к верхушке и к зоне соединения желудочка с аортальной луковицей (основанием желудочка), а также от эндокарда к эпикарду.
Работа выполнена в рамках темы «Сравнительно-физиологическое исследование пространственно-временной организации электрофизиологических процессов и сократимости миокарда позвоночных животных (№ ГР АААА-А17-1170123
10154-6) по Программе ФНИ на 2013–2020 гг. и в рамках Комплексной программы УрО РАН №ГР АААА-А18-118012290365-2 (2018–2020).
Список литературы Последовательность активации миокарда желудочка атлантической трески (Gadus morhua marisalbi)
- Dillon S., Morad M. A new Laser scanning system for measuring action potential propagation in the heart. Science. 1981. Vol. 214. No. 4519. P. 453-456
- Hu N., Sedmera D., Yost H.J., Clarc E.B. Structure and function of the developing zebrafish heart. Anat. Rec. 2000. Vol. 260. P. 148-157
- Sedmera D., Reckova M., DeAlmeida A., Sed- merova M. et al. Functional and morphological evidence for a ventricular conduction system in zebrafish and Xenopus hearts // Am. J. Physiol. Heart Circ. Physiol. 2003. Vol. 284. P. 1152-1160
- Шмаков Д.Н., Рощевский М.П. Активация миокарда / Институт физиологии Коми научного центра УрО РАН. Сыктывкар, 1997. 165 c
- Santer R.M. Morphology and innervation of the fish heart//Adv.Anat. Embryol. Cell.Biol. 1985. Vol. 89. Р. 100-102
- Ostadal B., Ostadalova I., Dhalla N.S. Development of cardiac sensitivity to oxygen deficiency: comparative and ontogenetic aspects// Physiol. Rev. 1999. Vol. 79. Р. 635-659
- Ostadal B., Rychter Z., Poupa O. Comparative aspects of the development of the terminal vascular bed in the myocardium // Physiol. Bohemoslov. 1970. Vol. 19. Р. 1-7
- Tota B. Vascular and metabolic zonantion in the ventricular myocardium of mammals and fishes // Comp.Biochem.Physiol. 1983. Vol. 76A. Р. 423-438
- Santer R.M., GreerWalker M., Emerson L., Witthames P.R. On the morphology of the heart ventricle in marine teleost fish (Teleostei) // Comp. Biochem. Physiol. 1983. Vol. 76. Р. 453-459
- Simхes K., Vicentini C.A., Orsi A.M., Cruz C. Myoarchitecture and vasculature of the heart ventricle in some fresh water teleosts // J. Anat. 2002. Vol. 200. Р. 467-475
- Последовательность реполяризации миокарда желудочка щук / М.А.Вайкшнорайте, А.С. Цветкова, В.А. Витязев, Я.Э. Азаров, Д.Н.Шмаков // Рос. физиол. журн. им. И.М. Сеченова. 2009. Т.95. № 2. С. 116-122
- The contribution of ventricular apicobasal and transmural repolarization patterns to the development of the T wave body surface potentials in frogs (Rana temporaria) and pike (Esox lucius)/M.A.Vaykshnorayte, J.E.Aza- rov, A.S.Tsvetkova, V.A.Vityazev, A.O.Ovech- kin, D.N.Shmakov // Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2011. Vol. 159. № 1. Р. 39-45
- Вайкшнорайте М.А. Последовательность активации миокарда желудочка карпа (Сyprinus carpio) // Рос. физиол. журн. им. И.М. Сеченова. 2018. Т. 104. № 2. С. 238-244
- Кузьмин В.С., Абрамочкин Д.В., Тарасова О.С., Ловать М.Л. Практические задачи по физиологии морских животных: учебное пособие. М.: Университетская книга, 2014. 188 с
- Лукьянов А.Н., Сухова Г.С., Удельнов М.Г. Локализация и структурно-функциональная организация пейсмекера сердца трески // Журн. эволюционной биохимии и физиологии. 1983. Т. 3. С. 231-236
- Патент. Российская Федерация. №2167599/ В.А.Витязев, Д.Н.Шмаков. Способ изготовления игольчатого электрода. Бюл. №15. 2001
- Вайкшнорайте М.А., Цветкова А.С., Витязев В.А., Азаров Я.Э. Зависимость последовательности реполяризации от последовательности активации на фрагменте эпикарда желудочка окуня (Perca fluviatilis) // Основные проблемы естественных и математических наук: Сборник научных трудов по итогам Международной научно-практической конференции. Волгоград, 2017. №4. 28 с
- Кардиоэлектрическое поле на эпикарде и поверхности тела щук в период деполяризации и реполяризации миокарда желудочка/ М.А.Вайкшнорайте, А.С.Белоголова, В.А.Витязев, Я.Э.Азаров, Д.Н. Шмаков // Рос. физиол. журн. им. И.М. Сеченова. 2007. Т.93. № 8. Р. 870-877
- Kochová P., Cimrman R., Štengl M., Ošťádal B., Tonar Z. A mathematical model of the carp heart ventricle during the cardiac cycle // J. Theor. Biol. 2015. Vol. 373. Р. 12-25
