Поведение и распределение молоди теплолюбивых видов рыб в термоградиентных условиях в летний и зимний сезоны года

Автор: Капшай Дмитрий Сергеевич, Голованов Владимир Константинович
Журнал: Инженерные технологии и системы @vestnik-mrsu
Рубрика: Экология
Статья в выпуске: 3-4, 2013 года.
Бесплатный доступ
В статье определяется значение окончательно избираемой температуры у молоди карпа (сазана), серебряного карася, леща и головешки-ротана в летний и зимний сезоны.
Молодь рыб, термоизбирание, избираемая температура, окончательно избираемая температура, термоградиентные условия, карповые, головешка-ротан
Короткий адрес: https://sciup.org/14720043
IDR: 14720043 | УДК: 597:
Behaviour and distribution of warm-requiring species juvenile fish in thermogradient conditions in summer and winter seasons of year
In summer and winter seasons of a year the value of final preferred temperature in the juvenile carp, silver crucian, bream and Amur sleeper were defined. The most thermophilic species - carp, silver crucian and Amur sleeper - in both seasons firmly selected the same level of final preferred temperature. On the contrary, it was revealed that bream fingerlings in different seasons of year prefer two vastly differing levels of temperature.
Текст научной статьи Поведение и распределение молоди теплолюбивых видов рыб в термоградиентных условиях в летний и зимний сезоны года
Терморегуляционное поведение (термо-избирание) – особая форма температурной адаптации, в процессе которой рыбы самопроизвольно избирают зону эколого-физиологического оптимума в экспериментальных или естественных условиях. Окончательно избираемая температура (ОИТ) рыб зависит от ряда биотических и абиотических факторов (возраста рыб, времени суток, физиолого-биохимического и иммунологического статуса рыб, а также сезона года и др.) [1; 2; 6–8; 11; 13; 14].
Несмотря на большое количество публикаций, освещающих роль разных факторов в термоизбирании рыб, влияние сезона на поведение и распределение молоди рыб в термоградиентных условиях изучено недостаточно полно. Известно два варианта выбора ОИТ в зависимости от сезона. Так, для некоторых теплолюбивых, а также холодолюбивых видов рыб показано, что значение ОИТ относительно постоянно как зимой, так и летом [1; 2]. В то же время для некоторых видов карповых более характерным является выбор двух уровней ОИТ, в зимний и летний сезоны. Выбор не одного, а двух существенно различающихся значений ОИТ в условиях экспериментального термо- градиента, связывают с наличием двух типов белкового и углеводного обмена веществ – летнего и зимнего [5; 8]. У рыб, которые постоянно выбирают только один устойчивый уровень ОИТ в разные сезоны, предполагается отсутствие положительной корреляции между температурой акклимации и ОИТ [2; 8]. Исследование данного вопроса позволяет глубже понять процессы и механизмы термоизбирания как одной из форм температурной адаптации.
Существенный интерес представляет экспериментальная проверка наличия одного и двух уровней ОИТ у молоди четырех видов особенно теплолюбивых и теплолюбивых видов рыб в два сезона – летний и осенне-зимний.
Исследование поведения и распределения в экспериментальных термоградиентных условиях, а также определение избираемой температуры (ИТ) и ОИТ у молоди карпа Cyprinus carpio , серебряного карася Carassius auratus , леща Abramis brama и головешки-ротана Perccottus glenii проводилось в летний и осенне-зимний сезоны 2009–2012 гг.
Объекты исследования, серебряный карась и головешка-ротан, были отловлены в прудах на побережье Рыбинского водохранилища. Карп выращивался в прудах на стационаре полевых и экспериментальных работ «Сунога» ФГБУН Института биологии внутренних вод им. И. Д. Папанина РАН. Лещ отлавливался в прибрежье Рыбинского водохранилища. После отлова рыб поместили в лабораторные аквариумы объемом 200 л с аэрацией. Перед каждым опытом особей акклимировали в течение 10 суток. Температура акклимации у молоди карпа, серебряного карася, леща и голо-вешки-ротана в летний сезон составила 18, 22, 24 и 19 °С, в осенне-зимний сезон – 11, 12, 14 и 3 °С соответственно. Все опыты проводили в условиях естественного фотопериода. В период акклимации и во время экспериментов рыб кормили 1–2 раза в сутки личинками хирономид и рыбным фаршем из расчета 5 % от общей массы тела. Корм в термоградиентной установке размещали в один или несколько отсеков, в которых на момент наблюдения находились рыбы.
В летний сезон длина и масса тела у карпа составляли 6,1±0,4 см и 9,8±1,2 г, у серебряного карася – 6,9±0,2 см и 8,9±1,0 г, у леща – 5,3±1,0 см и 2,5±0,2 г, у головеш-ки-ротана – 3,2±0,1 см и 1,2±0,2 г; в зимний сезон – 7,1±0,2 см и 11,3±0,8 г у карпа, 4,3±0,1 см и 3,2 ±0,5 г – у серебряного карася, 7,9 ±0,1 см и 5,8±0,3 г – у леща, 4,2±0,1 см и 1,6±0,2 г – у головешки-ротана.
ИТ и ОИТ определяли методом термо-преферендума в горизонтальной термоградиентной установке, представляющей собой лоток из прозрачного стекла размером 320x23x17 см [2-4]. Значения ИТ отражают выбор температуры в начальный период опыта, значения ОИТ – на стабильном участке выбора. Горизонтальный градиент температуры создавали посредством нагрева и охлаждения воды на противоположных концах установки, в которых разница температуры составляла 20 °С. Лоток делили с помощью неполных перегородок на 12 камер, в каждой из которых устанавливали по 2 распылителя с подачей воздуха для предотвращения вертикального градиента температуры. Рыб помещали в отсек с температурой, равной температуре акклимации. ИТ и распределение рыб регистрировали визуально 10 раз в светлое время суток. Для определения зоны ОИТ был принят 3-суточный критерий стабильного выбора, когда достоверные различия среднесуточных значений ИТ в течение трех последовательных дней опыта отсутствовали [2]. Продолжительность каждого опыта составляла 10 суток. Использовано по 10 экземпляров каждого вида в опытах с одной повторностью в летний и зимний сезоны.
Данные по ОИТ представлялись в виде средних значений. Поскольку методические разработки А. М. Свирского и В. Г. Терещенко [10] и анализ данных [2] показали, что ошибка определения ОИТ у группы особей в горизонтальных термоградиентных установках с учетом всех методических погрешностей составляет ±1 °С, различия показателей, превышающие 1 °С, считались достоверными. Более детально методика изложена ранее [3; 4].
В первые сутки эксперимента были выявлены и сходство, и различия в поведении и распределении рыб разных видов в экспе- риментальных термоградиентных условиях. Так, молодь леща в первые часы проявляла индифферентное отношение к температурному градиенту. Значения ИТ сеголеток после посадки в градиент возрастали всего на 1,0 °С летом и 1,8 °С зимой. Сеголетки других видов продвигались вверх по градиенту температуры более активно. У серебряного карася значения ИТ возрастали на 8,8 °С летом и 6,5 °С зимой. Особи карпа и голо-вешки-ротана в первые сутки опыта продемонстрировали больший рост ИТ в зимний период (13,6 °С и 13,6 °С), чем в летний (10,9 °С и 4,6 °С). Это может быть связано с бульшей разницей значения ОИТ и температуры акклимации в зимний период. Таким образом, имея сходный уровень ОИТ в разные сезоны года, рыбы, помещенные в температурные условия, соответствующие сезонным значениям, быстрее уходили из энергетически невыгодных условий, стремясь к более оптимальным физиологическим температурам [2; 12]. Таких резких изменений ИТ, как в первые сутки, в дальнейшем уже не наблюдалось.
Следует отметить, что ИТ в первый день опыта являлась интегративным показателем, характеризующим поведение молоди рыб при попадании в новую термоградиентную среду и показывающим, насколько быстро происходит адаптация в сложных лабиринтных условиях термоградиентной установки. «Миграционная активность» сеголеток может быть как высокой, так и практически нулевой (у леща).
Молодь трех видов – карпа, серебряного карася и головешки-ротана, независимо от сезона года, достаточно быстро передвигалась в градиентных условиях температуры в область высокой температуры 25–31 °С (рисунок). Скорость выбора в разные сезоны года значений ИТ, близких к ОИТ, а также кривые среднесуточных значений ИТ, сходны у серебряного карася и головешки-ротана, однако несколько отличаются у карпа. В зимний сезон молодь карпа выбирает ОИТ несколько медленнее. Серебряный карась и головешка-ротан, несмотря на сезонные отличия температуры акклимации, уже со 2–3-х суток опыта передвигались по градиенту температуры почти идентично. В то же время выбор ИТ в процессе опыта у мо- лоди леща кардинально отличался от трех остальных видов. Значения ИТ к концу опыта различались почти на 10 °С, показывая, что предварительная сезонная температура акклимации влияет на уровень ОИТ и динамику процесса выбора.
Значение ОИТ молоди карпа в летний и зимний период отличались незначительно (28,7±0,3 °С и 29,2±0,2 °С соответственно). У молоди серебряного карася и головешки-ротана значения ОИТ в летний сезон несколько выше (30,2±0,3 °С и 30,0±0,1 °С), чем в осенне-зимний период (28,4±0,5 °С и 27,9±0,1 °С соответственно). Динамика ИТ у карпа в течение всего опыта карпа в летний и зимний периоды показала, что летом сеголетки достигали температуры, близкой к уровню ОИТ быстрее, чем зимой, но в обоих случаях в зоне ОИТ особи концентрировались только на 7–9-е сутки. Более быстрое повышение ИТ у карпа могло быть вызвано более высокой температурой акклимации, и, соответственно, меньшим временем необходимым для постепенной переакклимации и перестройки физиолого-биохимических процессов в организме рыб. Молоди серебряного карася и головешки-ротана требовалось меньше времени для достижения зоны ОИТ, чем сеголеткам карпа. У молоди леща было обнаружено два уровня значения ОИТ: «летний» – 22,8±0,2 °С и «зимний» – 13,5±0,1 °С.
Полученные результаты подтверждают предположение о существовании двух типов адаптации у бореальных пресноводных рыб в процессе сезонных изменений температуры рыб в естественных водоемах [1; 2; 6–8]. Два сезонных уровня ОИТ свойственны также отдельным особям молоди речного окуня Perca fluviatilis [9]. Отметим, что один, не зависимый от сезона года, уровень ОИТ, очевидно, свойственен самым теплолюбивым видам, обитающим в пресных водоемах России. Необходимы дальнейшие исследования по выяснению того, насколько характерно это явление для молоди более взрослых особей других теплолюбивых и холодолюбивых видов рыб.
Таким образом, в ходе проведенных исследований было выявлено, что у наиболее теплолюбивых видов рыб – карпа, серебряного карася и головешки-ротана существует
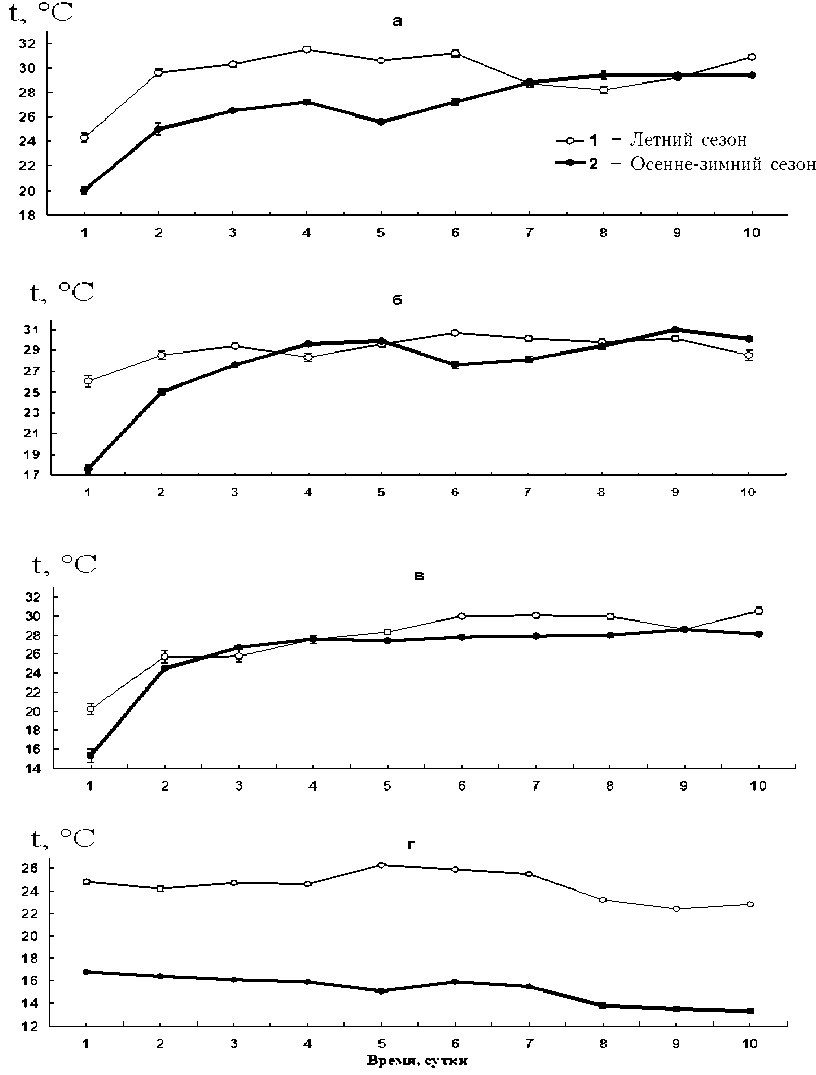
Рисунок. Динамика ИТ у молоди рыб: карпа (а), серебряного карася (б), головешки-ротана (в) и леща (г)
один стабильный уровень ОИТ, не зависящий от температуры предварительной сезонной акклимации. Для сеголеток леща было выявлено два уровня ОИТ в разные сезоны года, существенно различающихся по значению.
Результаты работы, имеющие предварительный характер, представляют значительный интерес для ихтиологов, специалистов высшей школы и рыбного хозяйст- ва. В первую очередь потому, что значение ОИТ характеризует эколого-физиологический оптимум молоди рыб. М. Джоблинг подтвердил эту зависимость на примере 49 видов рыб, показав высокую корреляцию между температурой оптимального роста и значением ОИТ [12]. Такое же соответствие было отмечено нами на примере молоди 11 видов пресноводных бореальных видов рыб [2].
Список литературы Поведение и распределение молоди теплолюбивых видов рыб в термоградиентных условиях в летний и зимний сезоны года
- Голованов В. К. Эколого-физиологические аспекты терморегуляционного поведения пресноводных рыб/В. К. Голованов//Поведение и распределение рыб: докл. 2-го Всерос. совещания «Поведение рыб». -Борок, 1996. -С. 16-40.
- Голованов В. К. Эколого-физиологические закономерности распределения и поведения пресноводных рыб в термоградиентных условиях: автореф. дисс.. д-ра биол. наук/В. К. Голованов. -М., 2012. -47 с.
- Голованов В. К. Экспериментальная оценка температурного оптимума у молоди пресноводных видов рыб методом термопреферендума/В. К. Голованов//Вестн. Морд. гос. ун-та. -(Сер. «Биолог. науки»). -2013. -№ 3-4. -С. 71-77.
- Голованов В. К. Сравнительный анализ окончательно избираемой и верхней летальной температуры у молоди некоторых видов пресноводных рыб/В. К. Голованов, А. К. Смирнов, Д. С. Капшай//Тр. Карел. НЦ РАН. -(Сер. Эксперимент. биология). -2012. -№ 2. -С. 70-75.
- Кирсипуу А. О сезонных изменениях белкового обмена у леща/А. Кирсипуу, К. Лаугасте//Экологическая физиология рыб. -М., 1973. -С. 211-212.
- Термоадаптационные характеристики леща Abramis brama (L.) Рыбинского водохранилища/В. В. Лапкин [и др.]//Структура локальной популяции у пресноводных рыб. -Рыбинск: Дом печати, 1990. -С. 37-85.
- Лапкин В. В. Возрастная динамика избираемых и летальных температур рыб/В. В. Лапкин, А. М. Свирский, В. К. Голованов//Зоол. журн. -1981, Т. 40, № 12. -С. 1792-1801.
- Поддубный А. Г. Сезонная динамика избираемых температур рыб/А. Г. Поддубный, В. К. Голованов, В. В. Лапкин//Теоретические аспекты рыбохозяйственных исследований водохранилищ. -Л., 1978. -С. 151-167 -Тр. ИБВВ АН СССР; вып. 32 (35).
- Свирский А. М. Сезонная и возрастная изменчивость избираемых температур у рыб Рыбинского водохранилища: Окунь (Perca fluviatilis L.)/А. М. Свирский, В. В. Лапкин//Биология внутренних вод: информац. бюл. -1987. -№ 76. -С. 45-49.
- Свирский А. М. Точность определения температуры, избираемой рыбами в установке с горизонтальным термоградиентом/А. М. Свирский, В. Г. Терещенко//Биология внутренних вод: информац. бюл. -1992.-№ 92. -С. 85-88.
- Beitinger Т. L. Physiological and ecological correlates of preferred temperature in fish/T. L. Be-itinger, L. C. Fitzpatrick//Amer. Zool. -1979. -Vol. 19, № 1. -P. 319-329.
- Jobling M. Temperature tolerance and the final preferendum -rapid methods for the assessment of optimum growth temperature/M. Jobling//J. Fish. Biol. -1981. -Vol. 19, № 4. -P. 439-455.
- Reutter J. M. Laboratory estimates of the seasonal final temperature preferenda of some Lake Erie fish/J. M. Reutter, C. E. Herdendorf//Proc. 17 th. Conf. Great Lakes Res. -Hamilton, 1974. -Part 1. Ann. Arbor., Mich. -1974. -P. 59-67.
- Reynolds W. W. Behavioral thermoregulation and the «final preferendum» paradigm/W. W. Reynolds, M. E. Casterlin//Amer. Zool. -1979. -Vol. 19, № 1. -P. 211-224.
