Позднеантропогеновые зайцы Сибири. К вопросу о филетической эволюции внутри рода Lepus

Автор: Оводов Н.Д.
Журнал: Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий @paeas
Рубрика: Археология каменного века палеоэкология
Статья в выпуске: XII-1, 2006 года.
Бесплатный доступ
Естественнонаучные методы в археологии, палеозоология, антропогеновый период, антропогеновая фауна, сибирь, филетическая эволюция, зайцы, род lepus
Короткий адрес: https://sciup.org/14521183
IDR: 14521183
Текст статьи Позднеантропогеновые зайцы Сибири. К вопросу о филетической эволюции внутри рода Lepus
А.А.Борисяк (1947)
Проблема процесса видообразования на основе изучения изменчивости признаков, вероятно, самая захватывающая по интересу для биологов, и, в частности, для палеотериологов. Общие принципы и конкретные примеры её решения приведены во многих работах: Д.Г.Симпсон (1948), В.О.Ковалевский (1956), А.В.Яблоков (1966), И.И.Шмальгаузен (1968), Э.Майр (1974), Н.В.Тимофеев-Ресовский, Н.Н.Воронцов, А.В.Яблоков (1969), С.С.Шварц (1980), Н.Н.Воронцов (2005). По словам Эрнста Майра (1968, с. 35): «…в природе можно найти много популяций, которые прошли лишь часть пути, ведущего к статусу вида. Если бы от всех популяций сохранялись ископаемые остатки, то наличие промежуточных популяций между сменяющими друг друга видами было бы нормальной ситуацией в палеонтологии. На самом же деле пробелы в палеонтологической летописи так многочисленны, что лишь в очень немногих случаях удается восстановить все звенья цепи и получить непрерывную линию, связывающую хорошие виды».
Что можно сказать относительно зайцев в свете выраженной Э.Майром мысли? В свое время известный палеотериолог, мой бывший учитель проф. Н.К.Верещагин, касаясь проблемы происхождения современного зайца-беляка, безответно задал вопрос: «Спрашивается, однако, где же был в это время (финал плейстоцена,- Н.Ов.) предок зайца трусливого – ведь со времени существования донского зайца прошло всего 10-12 тыс. лет» (Верещагин, 1977, с. 12-13). Теперь, кажется, можно дать ответ.
Донской заяц (Lepus tanaiticus Gureev) для науки «молодой» вид, - его остатки описал 40 лет назад А.А.Гуреев (1964) по материалам из слоёв Костёнковских палеолитических стоянок с берегов р. Дон. Ко времени позднего плейстоцена ареал донского зайца занимал, по-видимому, значительную часть северной половины бывшего СССР, биотопически соседствуя в западных районах с зайцем-русаком. Существенный вклад в изучение этого вида, обитавшего на территории Сибири, внёс А.О.Аверьянов (1995).
Касаясь вопроса появления современных арктических зайцев (Lepus timidus) Евразии, этот автор категорично пишет, что они «не могли происходить от L. tanaiticus» (Аверьянов, 1995а, с.595). Для решения этой проблемы желательно и необходимо иметь достаточно многочисленный хронологически выстроенный материал от позднеплейстоценового времени до современности и по-возможности с ограниченной территории.
Материалом для настоящего сообщения послужили остатки донских зайцев из палеолитических стоянок Лиственка (раскопки Е.В.Акимовой), Караульный Бык (раскопки Н.П.Макарова) и Афонтова Гора III (раскопки Н.К.Ауэрбаха с коллегами), расположенных в окрестностях г. Красноярска (коллекция Красноярского краеведческого музея и Гос. пед. университета). Голоценовые кости зайца-беляка собраны мной за 30-летний период в пещерах Алтая, по югу Средней и Восточной Сибири.
Сравнение скелетных элементов плейстоценового донского зайца и голоценового беляка (таблица 1, измерения сделаны автором) указывают, что за редчайшим исключением последний в размерах проигрывает донскому зайцу в массивности туловища (конечностей) и головы, что отмечал в своих работах и А.О.Аверьянов. Находясь в меньшей «весовой категории», голоценовый беляк легче передвигался по снегу и соответственно был более проворен, спасаясь от хищников. Особо заметны отличия в размерах и форме черепных элементов обоих зайцев, в частности, это относится к внешнему радиусу кривизны резцов, на что раньше зоологи не обращали внимание.
По мнению первоописателя донского зайца А.А.Гуреева (1964, с. 188): «Крупные размеры угловой лопасти нижней челюсти указывают на значительное развитие жевательных мышц, а высокие коронки коренных зубов и короткая, круто изогнутая вверх резцовая часть нижней челюсти позволяют предположить, что пищей этому виду служили преимущественно грубые корма, особенно в зимнее время » (выделено мной,- Н.Ов). Однако, учитывая пологий радиус резцов легче и правильней представить донского зайца, питающегося не только летом, но и зимой в значительной мере травянистой растительностью. Именно эти анатомическая и этологическая особенности донских зайцев, по-видимому, явились на фоне перестройки состава фауны основной причиной вымирания рассматриваемого вида.
Многим, очевидно, не только по литературе известно, что даже в малоснежных районах скотоводы выгоняют на пастбища овец вместе с табуном лошадей, тебенюющих снег. Аналогичную картину не сложно перенести в эпоху позднего плейстоцена Северной Азии, когда популяции растительноядных млекопитающих: мамонтов, диких лошадей, носорогов, бизонов, разгребали снег в поисках жухлой травы и низких кустарничков, давая зайцем вслед за тем возможность кормиться травой. С гибелью «мамонтовой фауны», совпавшей по времени в течение нескольких тысячелетий с полным вымиранием донского зайца, выжили те «наследники», будущие твердые Lepus timidus, которым в силу полезных мутаций удалось укоро-
Таблица 1. Различия в размерах костей скелета донского зайца и современного беляка (мм)
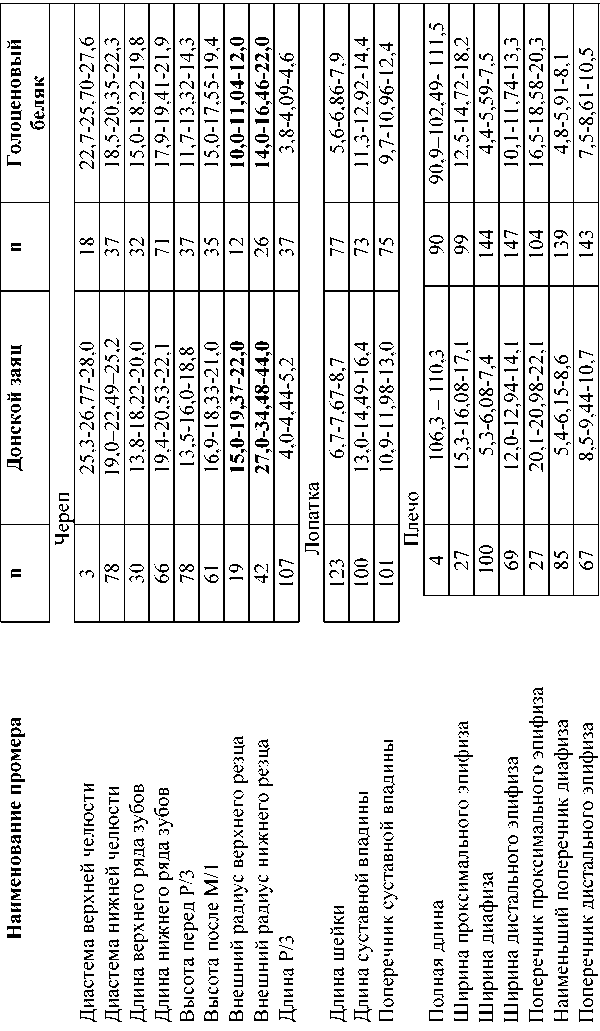
Продолжение табл.
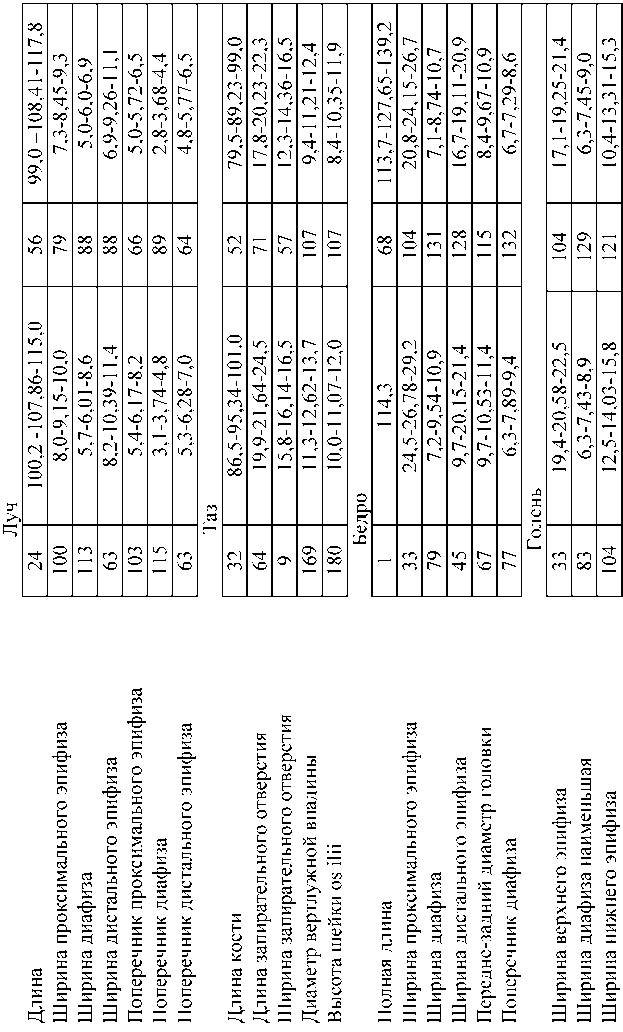
Продолжение табл.
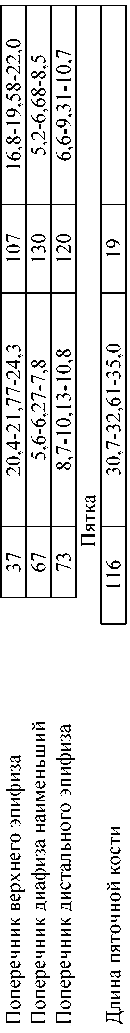
тить радиусы резцов, превратив их в «кусачки» для эффективного питания ветками и корой. О том же свидетельствует уменьшенная площадь жевательной поверхности щёчных зубов у голоценовых беляков. Процесс совершенствования резцового аппарата позволил белякам, обитающим нынче на территории Байкало-Ленского заповедника, подгрызать и валить деревца с диаметром ствола до 4,5 см (Мельников, 2003). К сожалению, зоологи отловили этих “инакодействующих” прогрессивных животных, оборвав зарождающуюся ниточку эволюционного процесса.
Постепенность реального процесса укорочения радиуса резцов удалось проследить на материалах раскопок нижних слоёв рыхлых отложений пещеры Еленева (окрестности Красноярска), обеспеченной 49 радиоуглеродными датами в пределах от 1050 до 13665 лет (Оводов, Мартынович, Орлова, 2003). Интересно проверить насколько чётко размер радиуса резцов донских зайцев, коррелирующий по времени с вымиранием плейстоценовой мегафауны на Среднем Енисее, будет проявляться в других регионах. Думаю, отнюдь не всегда картина будет столь же стройная, какая проявилась на памятниках окрестностей Красноярска. В частности, остатки донского зайца обнаружены мной в слоях культурного мезолитического слоя стоянки Усть-Шилка-2 (раскопки П.В.Мандрыки), расположенной на берегу Енисея в 200 км ниже Красноярска. Фаунистический материал этой стоянки имеет почти целиком яркий голоценовый облик (Мандрыка, Оводов, 2005).
Вместо резюме : Э . Каляева, отдавая дань памяти своемуучителюталантливомубиологуН.Н.Воронцову (1934-2000 гг.), рано оценившему направление и результаты работ Т.Д.Лысенко, захватившего власть на генетической ниве СССР, пишет (Каляева, 2005) по поводу лекций, читанных 20-летним Николаем Николаевичем московским юным биологам: «Всё это коренным образом отличалось от школьной программы по биологии, где в те годы господствовали взгляды академика Т.Д.Лысенко, который допускал превращение одного вида в другой в результате изменения состава пищи. Типичный пример – утверж- дение Т.Д.Лысенко, что если пеночку кормить мохнатыми гусеницами, из её яиц выведутся кукушата». Можно ли в таком случае считать, что в «истории» с зайцами мы имеем чистую лысенковщину, хотя она касается не межродовых бредовых идей, а эволюционно последовательных изменений внутри рода Lepus, приведших к появлению нового вида?
