Позднеголоценовые птицы из археологической стоянки окрестностей г. Салехарда (север Западной Сибири)
")
Автор: Пантелеев А.В., Потапова О.Р.
Журнал: Русский орнитологический журнал @ornis
Статья в выпуске: 106 т.9, 2000 года.
Бесплатный доступ
Короткий адрес: https://sciup.org/140149052
IDR: 140149052
Late holocene birds from an archaeological site near Salekhard (north of Western Siberia)
Текст статьи Позднеголоценовые птицы из археологической стоянки окрестностей г. Салехарда (север Западной Сибири)
Сведения по ископаемым птицам северных районов России чрезвычайно малочисленны. До недавнего времени самым северным материковым местонахождением, содержащим остатки птиц, была Медвежья пещера на Северном Урале (Потапова 1990), находящаяся в подзоне средней тайги. Стоянка у Салехарда (Усть-Полуйское городище) — значительно севернее, в зоне лесотундры. Расположение в зоне контакта тундровой и таёжной фаун, на берегу крупной сибирской реки, вблизи от Уральских гор вызывает дополнительный интерес к этому местонахождению.
Городище находится на правом берегу р. Полуй, в 3 км ниже г. Салехарда (до 1933 — Обдорск) и в 4 км выше Ангальского мыса. С напольной стороны поселение было обнесено рвом и валом, выходящими с одной стороны к обрыву, с другой — в небольшой ложок, заболоченный в настоящее время. Население Усть-Полуя было оседлым. Хозяйство носило комплексный характер. Оно базировалось на охоте (главным образом на северного оленя), промысле морского зверя (белуха, морж) и рыболовстве, при наличии развитого упряжного собаководства (Мошинская 1953).
Материал и его возраст. Методика
Местонахождение было случайно открыто в 1932 г. во время строительных работ. Материал собран в 1935-1936. при раскопках, которыми руководил сотрудник Музея антропологии и этнографии АН СССР В.С.Адрианов. В 1935 раскопки проводились в рамках разведочной экспедиции Зоологического института АН СССР по обследованию условий залегания трупов мамонтов в пределах Ямальского национального округа. В 1935 было вскрыто 234.5 м2 отложений, в 1936 — примерно такая же площадь или несколько больше. Раскопки не были закончены, а документация 1936 года исчезла.
Мощность культурного горизонта составляет 20-30 см, в углублениях до 50 см. В.С.Адрианов обратил внимание, что слой, содержащий различные культурные остатки, не однороден, и по всем разрезам можно было проследить две резко выраженные различной цветности прослойки, в некоторых местах расслаивающиеся ещё на несколько слоев (Адрианов 19366). Позднее, во время работ Мангазейской экспедиции в 1946, было установлено единообразие и монолитность культурного слоя, что указывает на длительность существования, но не на многослойность городища (Мошинская 1953).
В.Н.Чернецов считает, что городище возникло в IV в. до н. э., а закончило существование внезапно, где-то в пределах II-I вв. до н. э. Но ещё некоторое время продолжало существовать жертвенное место, находившееся в пределах городища. “Но, видимо, и жертвенное место не просуществовало долгое время, поскольку не только на нем, но и на всей территории городища, несмотря на общее обилие находок, не обнаружено ни одного экземпляра мелких парных золоченых бус, характерных для памятников, а тем более для жертвенных мест первых веков нашей эры” (Чернецов 19536, с. 228). И.А.Талицкая (1953) для этого местонахождения приводит несколько иной интервал: IV в. до н. э.-II в. н. э. Но это — время существования всей усть-полуйской культуры, а не данного городища. Во вводной части работы она сама пишет, что датировки даются очень обобщенно.
Таким образом, материал мог накапливаться в IV-I вв. до н. э.
Птичьи кости сбора 1935 года происходят, по-видимому, из V раскопа, которым вскрыты два или три жилых комплекса и жертвенное место. Здесь находилось большое кострище, рядом с которым обнаружена груда собачьих черепов, осколки костей оленей, отдельные кости человека, громадное количество осколков керамики и многочисленные поделки из кости и бронзы (Мошинская 1953).
Мы не первыми заинтересовались сборами из Усть-Полуйского городища. В архиве Аркадия Яковлевича Тугаринова была обнаружена рукопись статьи А.Я.Тугаринова и В.М.Коровина “Кости птиц из древнего поселения в низовьях Оби”. Однако при просмотре костей оказалось, что часть материала определена неправильно (кости определял В.М.Коровин, АЯ.Тугаринов писал текст статьи). Например, голень серебристой чайки была отнесена к свиязи, 2 локтевые кости стерха — к орлану-белохвосту, 2 крестца разных видов уток — к белой куропатке и т.п. Всего переопределено около 600 костей из 1238 определявшихся. Причём, присутствие таких видов, как серощёкая поганка, серый гусь, серая утка, красноносый и красноголовый нырки и сизая чайка не подтвердились. Кроме того, в Зоологическом институте РАН были найдены неизвестные ранее кости птиц из этого же местонахождения (744 экз., сборы 19351936 гг.), а список видов дополнился морской чернетью, синьгой, лутком, большим крохалем, сапсаном, полевым лунем, серым журавлем и бородатой неясытью. В связи с этим, а также с учётом новых литературных данных по современному распространению птиц на севере Западной Сибири, палеогеографические построения статьи пришлось пересмотреть. Определение костей и написание статьи выполнены, фактически, заново. Поэтому мы сочли возможным опубликовать эту работу под своими именами.
Для измерения костей куриных использовались промеры, предложенные A.Driesch (1976). Промеры костей гагар и орлана-белохвоста изображены на рис. 1; промеры с локтевых костей (os ulna) серебристой чайки снимались так же, как у гагар, с бедренных костей (os femur) гусей — так же, как у орланов; общая длина тибиотарсуса (os tibiotarsus) гусей (промер 1) и куропаток измерялась от вершины самого высокого бугорка на проксимальном эпифизе, расположенном между crista patellaris и facies artic. medialis, до дистального края condylus medialis, ширина дистального эпифиза (промер 2) — между внешними краями condylus medialis и condylus lateralis.
Используемые сокращения промеров костей куриных Вр — наибольшая ширина проксимального эпифиза;
GB — наибольшая ширина черепа в области скуловых дуг;
GL — наибольшая длина кости;
Gif — фиксированная длина коракоида от проксимального эпифиза до нижних краев выступов суставной поверхности дистального эпифиза;
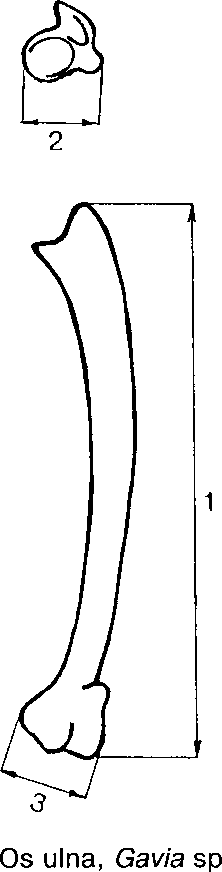
Рис. 1. Промеры костей,
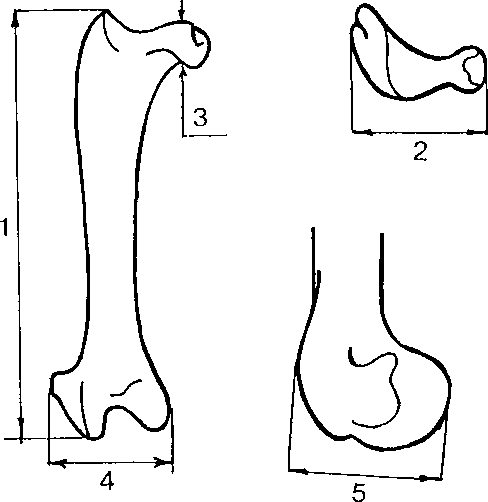
Os femur, Haliaeetus albicilla

Os carpometacarpus, Haliaeetus albicilla
используемые в работе. 1-7 — номера промеров.
Но — высота нижней челюсти у переднего края челюстного отверстия;
La — общая длина тибиотарсуса;
Lm — длина бедренной кости от верхнего края головки до медиального мыщелка:
Lp — длина черепной коробки от верхнего края foramen magnum до заднего края носовых костей.
И clav - clavicula quadr - quadratum cmc - carpometacarpus rad - radius cor - coracoideum scap - scapula
спользуемые сокращения
cran - cranium stern - sternum fem - femur syns - synsacrum hum - humerus tib - tibiotarsus
названий костей mand - mandibulae tmt - tarsometatarsus phal - phalanx vert - vertebra premax - premaxillare
Определение ископаемых костей проводили с помощью остеологических коллекций Зоологического и Палеонтологического институтов РАН. Статистические расчеты выполнялись в программе STATGRAPHICS, значимость отличий оценивалась по критерию Стьюдента (Ивантер, Коросов 1992).
Весь изученный материал из Усть-Полуйского городища хранится в отделениях орнитологии и истории фауны Зоологического института РАН.
Систематический список видов
2013 костных остатков из Усть-Полуйского городища принадлежали 39 видам птиц (табл. 1).
Таблица 1. Видовой состав птиц из Усть-Полуйского городища
|
Таксон |
Число костей |
Минимальное число особей |
|
Gavia stellata |
25 |
7 |
|
Gavia arctica |
48 |
10 |
|
Podiceps cristatus |
8 |
2 |
|
Cygnus cygnus |
28 |
5 |
|
Cygnus bewickii |
10 |
2 |
|
Anserct. albifrons |
132 |
41 |
|
Anserct. fabalis |
88 |
10 |
|
Branta sp. |
3 |
1 |
|
Anas platyrhynchos |
12 |
5 |
|
Anas crecca |
50 |
23 |
|
Anas penelope |
40 |
10 |
|
Anas acuta |
24 |
4 |
|
Anas querquedula |
6 |
2 |
|
Anas clypeata |
12 |
3 |
|
Anas sp. |
6 |
2 |
|
Aythya fuligula |
4 |
4 |
|
Aythya marila |
1 |
1 |
|
Melanitta fusca |
1 |
1 |
|
Melanitta nigra |
2 |
1 |
|
Clangula hyemalis |
14 |
5 |
|
Bucephala clangula |
7 |
2 |
|
Mergus albellus |
1 |
1 |
|
Mergus merganser |
1 |
1 |
|
Anatidae indet. |
46 |
18 |
|
Haliaeetus albicilla |
143 |
10 |
|
Accipiter gentilis |
4 |
1 |
|
Buteo lagopus |
1 |
1 |
|
Aquila chrysaetus |
28 |
7 |
|
Circus cyaneus |
2 |
1 |
|
Accipitridae indet. |
1 |
1 |
|
Falco peregrinus |
2 |
1 |
Продолжение таблицы 1
|
Таксон |
Число костей |
Минимальное число особей |
|
Lagopus mutus |
134 |
36 |
|
Lagopus lagopus |
653 |
144 |
|
Lagopus sp. |
318 |
30 |
|
Tetrao urogallus |
16 |
6 |
|
Lyrurus tetrix |
1 |
1 |
|
Grus grus |
1 |
1 |
|
Grus leucogeranus |
6 |
1 |
|
Larus argentatus |
65 |
10 |
|
Charadriiformes indet. |
1 |
1 |
|
Bubo bubo |
4 |
1 |
|
Nyctea scandiaca |
12 |
2 |
|
Strix nebulosa |
1 |
1 |
|
Corvus cornix |
11 |
3 |
|
Corvus corax |
11 |
2 |
|
Aves indet. |
12 |
|
|
Всего |
1996 |
422 |
Краснозобая гагара Gavia stellata (Pontopp.)
Материал: premax — 1, hum — 5 (3)*, ulna — 10 (7), rad — 3 (2), cmc — 4 (2), tib — 2 (1).
Ископаемые локтевые кости в среднем немного меньше современных (табл. 2). Найдена в пределах современной области гнездования.
Таблица 2. Промеры локтевых костей усть-полуйских (F) и современных (R) краснозобых гагар Gavia stellata
|
Статистики |
Промеры |
|||
|
1 |
2 |
3 |
||
|
п |
F |
8 |
7 |
9 |
|
R |
10 |
11 |
11 |
|
|
Х± S.E. |
F |
110.65±1.659 |
10.23±0.310 |
12.97±0.381 |
|
R |
114.12±1.739 |
10.21±0.245 |
13.07±0.438 |
|
|
lim |
F |
102.9-118.0 |
9.2-11.3 |
11.9-15.3 |
|
R |
106.1-122.9 |
8.8-11.9 |
11.0-16.3 |
|
Чернозобая гагара Gavia arctica (L.)
Материал: stern — 1, cor — 1, hum — 1, ulna — 12 (10), rad — 10 (7), cmc — 14 (8), tib - 5 (3), tmt - 4 (3).
Промеренные ископаемые локтевые кости в среднем немного меньше современных (табл. 3). В районе Салехарда — обычная гнездящаяся птица.
Таблица 3. Промеры локтевых костей усть-полуйских (F) и современных (R) чернозобых гагар Gavia arctica
|
Статистики |
Промеры |
|||
|
1 |
2 |
3 |
||
|
л |
F |
10 |
11 |
11 |
|
R |
5 |
5 |
5 |
|
|
Х± S.E. |
F |
117.17±1.603 |
11.59±0.140 |
15.81±0.237 |
|
R |
121.56±3.255 |
12.06±0.316 |
16.20±0.226 |
|
|
lim |
F |
112.0-127.0 |
11.0-12.7 |
14.9-17.2 |
|
R |
109.8-126.7 |
11.2-12.7 |
15.6-16.6 |
|
Чомга Podiceps cristatus (L.)
Материал: hum — 1, fem — 1, tib — 3 (2), tmt — 3 (2).
Северная граница гнездовой части ареала в бассейне Оби проходит по 57-й параллели (Степанян 1990). К.М.Дерюгин (1898) встретил эту птицу 26 июня 1897 близ Устремских юрт (Малая Обь, примерно 64°20' с.ш.). Весенний залёт отмечен в дельту р. Щучья (Данилов и др. 1984).
Лебедь-кликун Cygnus cygnus (L.)
Материал: cran — 2 (1), cor — 4 (3), hum — 7 (3), ulna — 1, syns — 1, fem — 7 (5), tib — 3 (2), tmt — 3 (3).
Немногочисленная гнездящаяся птица.
Тундряный лебедь Cygnus bewickii Yarr.
Материал: cran — 1, premax — 2 (2), hum — 1, ulna — 1, rad — 1, syns — 1, fem — 1, phal — 2 (1).
Многие исследователи приводят для нижней Оби только по одному виду лебедей — либо кликуна (Данилов 1965), либо тундряного (Дерюгин 1898; Шухов 1915), либо просто "лебедя" (Сурина 1967). Тундряный лебедь в районе Салехарда встречается лишь на пролёте, причём его весенняя миграция происходит позднее, чем у кликуна (Брауде 1987; Калякин, Виноградов 1987). Гнездится на южном Ямале.
Белолобый гусь Anser cf. albifrons (Scop.)
Материал: stern-3(3), cmc-1, fem-57(30), tib-71(41), tmt-1; одна голень была сломана и срослась при жизни птицы.
Бедренные кости усть-полуйских белолобых гусей по размерам соответствуют современным (табл. 4; рис. 2), а тибиотарсусы значительно длиннее (табл. 6; рис. 3). Белолобый гусь обычен на весеннем и осеннем пролётах. Выводок встречен на луже близ р. Байдараты, 68° с.ш. (Дерюгин 1898).
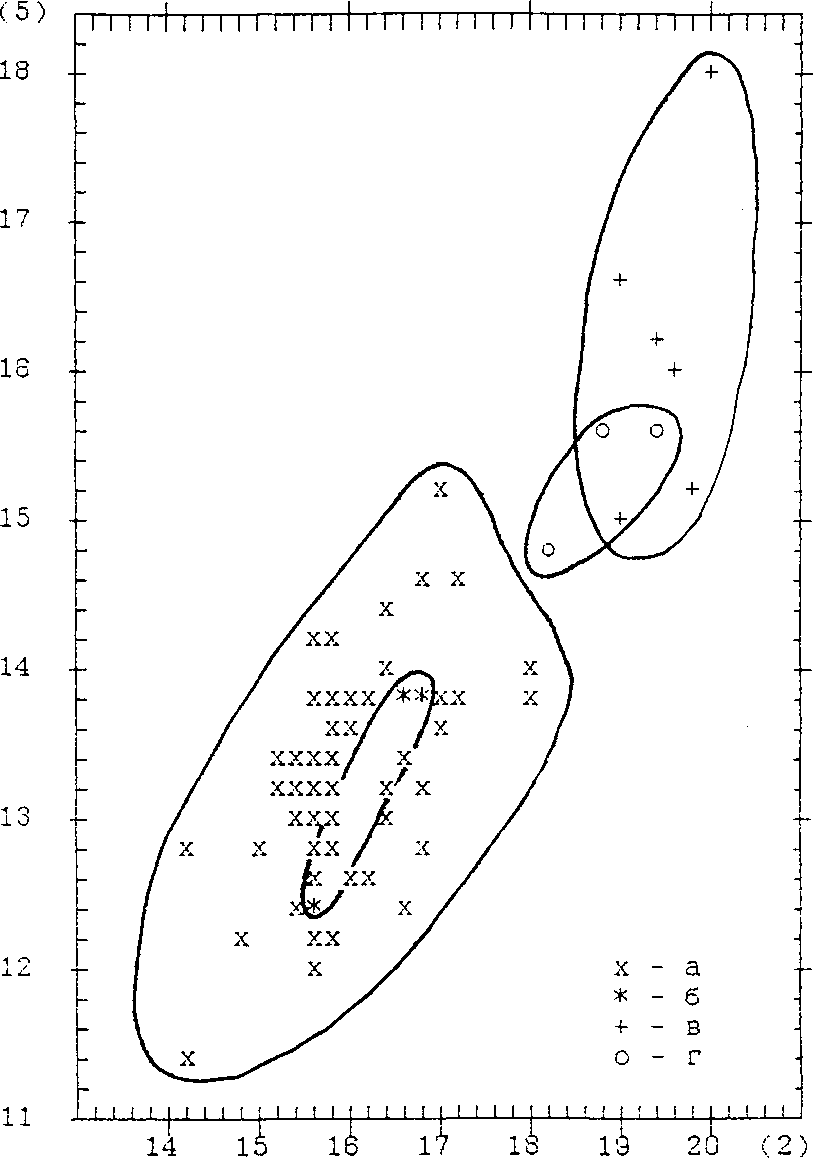
Рис. 2. Распределение бедренных костей белолобого гуся Anser albi-frons и гуменника Anser fabalis по ширине проксимального (2) и высоте дистального (5) эпифизов (размеры в мм).
Обозначения: а — усть-полуйские A. albifrons; б — современные A. albifrons; в — усть-полуйские A. fabalis; г — современные A. fabalis.
Таблица 4. Промеры бедренных костей усть-полуйских (F) и современных (R) белолобых гусей Anser albifrons
|
Статистики |
Промеры |
|||
|
1 |
2 |
3 |
5 |
|
|
п F |
52 |
56 |
51 |
48 |
|
R |
3 |
3 |
3 |
3 |
|
Х± S.E. F |
69.91±0.329 |
15.98±0.106 |
14.54±0.097 |
13.24±0.107 |
|
R |
67.80±1.442 |
16.33±0.367 |
14.77±0.561 |
13.30±0.451 |
|
lim F |
64.7-74.3 |
14.1-18.0 |
13.4-18.0 |
11.3-15.2 |
|
R |
65.0-69.8 |
15.6-16.7 |
13.7-15.6 |
12.4-13.8 |
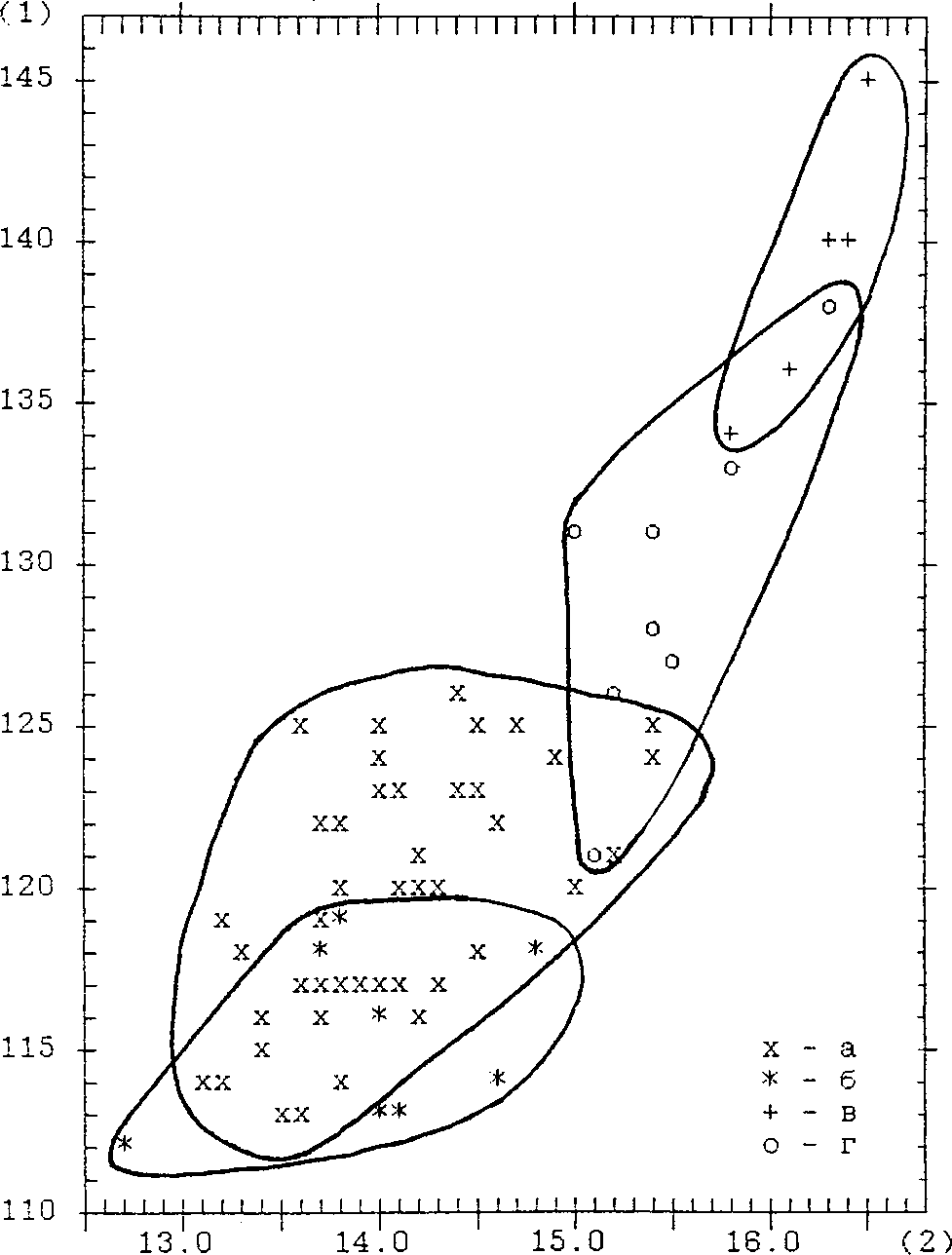
Рис. 3. Распределение тибиотарсусов белолобого гуся Anser albifrons и гуменника Anser fabalis по общей длине (1) и ширине дистального эпифиза (2). Размеры в мм.
Обозначения: а — усть-полуйские A. albifrons; б — современные A. albifrons; в — усть-полуйские A. fabalis; г — современные A. fabalis.
Гуменник Anser cf. fabalis (Lath.)
Материал: premax — 1, cor — 6 (4), scap — 6 (3), clav — 2 (2), stern — 8 (8), hum — 21 (9), rad — 1, cmc — 1, syns — 1, fem — 32 (17), tib — 39 (21), tmt — 4 (3).
Все элементы скелета усть-полуйских гуменников несколько крупнее современных. Особенно разница в размерах заметна в длине костей, по другим параметрам она меньше (табл. 5, 6; рис. 2, 3). То есть скелет (по крайней мере, ноги) современных гусей стал не только меньше, но и массивнее. Немногочисленный гнездящийся вид.
Таблица 5. Промеры бедренных костей усть-полуйских (F) и современных (R) гуменников Anser fabalis
|
Статистики |
Промеры |
||||
|
1 |
2 |
3 |
5 |
||
|
п |
F |
9 |
12 |
11 |
6 |
|
R |
3 |
3 |
3 |
3 |
|
|
Х± S.E. |
F |
81.20±0.922 |
19.38±0.143 |
17.55±0.251 |
16.13±0.426 |
|
R |
75.83±0.917 |
18.77±0.348 |
16.47±0.318 |
15.27±0.285 |
|
|
Пт |
F |
77.8-85.3 |
18.2-20.0 |
16.4-19.0 |
15.0-17.9 |
|
R |
74.0-76.8 |
18.2-19.4 |
15.9-17.0 |
14.7-15.6 |
|
Таблица 6. Промеры голени усть-полуйских (F) и современных (R) белолобых гусей Anser albifrons и гуменников Anser fabalis
|
Статистики |
Промеры |
|||
|
Anser albifrons |
Anser fabalis |
|||
|
1 |
2 |
1 |
2 |
|
|
л F 46 49 5 10 R 8 8 8 9 X+S.E. F 119.56±0.545 14.14±0.087 139.06±1.976 16.44±0.202 R 115.38±0.942 13.96±0.224 129.29±1.814 15.38±0.151 lim F 113.0-125.7 13.1-15.5 133.6-145.2 15.7-17.7 R 112.0-119.5 12.7-14.8 120.7-138.0 14.8-16.3 |
||||
Кряква Anas platyrhynchos L.
Материал: cor — 2 (1), stern — 1, hum — 5 (5), rad — 2 (1), cmc — 2 (1), fem — 1.
К.М.Дерюгин (1898, c. 112) указывает, что эта утка "под Обдорском, безусловно, не встречается". И.Н.Шухов (1915, с. 228) сообщает: "Под селом Обдорским этот вид встречается очень редко". В.И.Абрамов (1959, с. НО) пишет, что "не гнездящаяся в районе Салехарда кряква появляется там во второй половине лета. Всего вероятнее, что это перелинявшие и поднявшиеся на крыло селезни, прилетавшие в недоступные сора на линьку". Л.Н.Добринский встречал у Салехарда выводки (Данилов 1965).
Чирок-свистунок Anas crecca L.
Материал: cor — 1, stern — 1, hum — 41 (23), ulna — 6 (3), tib — 1.
Обычный гнездящийся вид.
Свиязь Anas penelope L.
Материал: cor — 1, stern — 1, hum — 16 (10), ulna — 8 (4), rad — 1, cmc — 7 (6), syns — 1, fem — 1, tib — 4 (2).
Многочисленная гнездящаяся птица.
Шилохвость Anas acuta L.
Материал: stern — 3 (3), hum — 6 (4), ulna — 6 (3), rad — 1, cmc — 3 (2), fem — 3 (2), tib - 2 (1).
Шилохвость — самая многочисленная гнездящаяся утка нижней Оби (Дерюгин 1898; Шухов 1915; Данилов 1965).
Чирок-трескунок Anas querquedula L.
Материал: cor — 2 (2), hum — 4 (2).
К.М.Дерюгин (1898) считал трескунка на нижней Оби редкой птицей. Сейчас это обычный гнездящийся вид (Данилов 1965), хотя большой численности и не достигает (Добринский 1959).
Широконоска Anas clypeata L.
Материал: cor — 2 (1), scap — 1, stern — 1, hum — 1, ulna — 1, cmc — 2 (1), tib — 3 (3), tmt — 1.
Обычный гнездящийся вид.
Хохлатая чернеть Aythya fuligula (L.)
Материал: cor — 4 (4).
Обычный гнездящийся вид.
Морская чернеть Aythya marila (L.)
Материал: cor — 1.
Немногочисленный гнездящийся вид.
Турпан Melanitta fusca (L.)
Материал: hum — 1.
Редкий гнездящийся вид.
Синьга Melanitta nigra (L.)
Материал: hum — 1, fem — 1.
Немногочисленный гнездящийся вид. На осеннем пролете может образовывать большие стаи (Сурина 1967).
Морянка Clangula hyemalis (L.)
Материал: cor — 2 (1), stern — 3 (3), hum — 7 (5), fem — 1, tib — 1, tmt — 1.
Немногочисленный гнездящийся вид.
Гоголь Bucephala clangula (L.)
Материал: cor — 2 (2), hum — 2 (2), ulna — 2 (2), cmc — 1.
Немногочисленная гнездящаяся птица.
Луток Mergas albellus L.
Материал: fem — 1.
Редкая гнездящаяся птица.
Большой крохаль Mergas merganser L.
Материал: cor — 1.
Редкий, возможно, гнездящийся вид. В.Н.Бойков (1965) встречал этих птиц в низовьях Полуя, Л.Н.Добринский (1965) находил выводки в нижнем течении Соби (впадает в Обь в 40 км южнее Салехарда).
Орлан-белохвост Haliaeetus albicilla (L.)
Материал: cran — 4 (4), prem — 2 (2), mand — 4 (4), cor — 9 (6), scap — 5 (4), stern — 1, hum — 15 (8), ulna — 16 (7), rad — 14 (8), cmc — 14 (8), syns — 6 (5), fem — 17 (10), tib — 15 (7), tmt — 15 (10), phal — 6 (2); 1 ulna и 1 tib были сломаны и срослись при жизни птицы.
Измерения бедренных костей и карпометакарпусов показали, что ископаемые орланы-белохвосты были в среднем немного мельче современных (рис. 4), причем, для общей длины os femur, высоты основания головки этой же кости и общей длины os carpometacarpus эти различия статистически значимы (табл. 7, 8).
Обычная гнездящаяся птица.
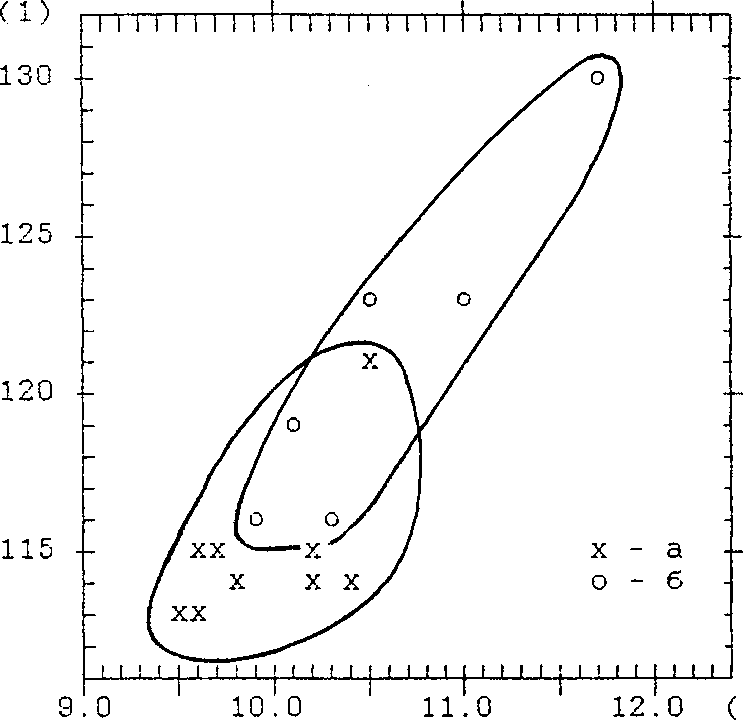
Рис. 4. Распределение бедренных костей орлана-белохвоста Haliaeetus albicilla по общей длине (1) и высоте основания головки (3). Размеры в мм. Обозначения: а — усть-полуйские Н. albicilla; б — современные Н. albicilla.
Тетеревятник Accipiter gentilis (L.)
Материал: hum — 1, ulna — 1, tib — 1, tmt — 1.
В мае, июле, августе и октябре тетеревятников наблюдали в низовьях Полуя (Бойков 1965). Регулярно отмечаются встречи птиц на южном Ямале. В 1979 найдено гнездо на р. Хадытаяха (Данилов и др. 1984).
Мохноногий канюк Buteo lagopus (Pontopp.)
Материал: tmt — 1.
Немногочисленная гнездящаяся птица.
Таблица 7. Промеры карпометакарпусов усть-полуйских (F) и современных (R) орланов-белохвостов Haliaeetus albicilla
|
Статистики |
Промеры |
||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
|
п F 12 13 13 13 14 12 12 R 6 6 6 6 6 6 6 X+S.E. F 112.03±1.417 13.32±0.346 29.75±0.446 9.23±0.305 7.54±0.17 25.73±0.355 110.42±1.500 R 116.72±1.601 14.02±0.471 30.52±0.413 9.40±0.333 7.27±0.179 26.62±0.456 113.72±1.000 lim F 107.0-123.2 11.1-15.4 27.0-32.1 7.0-10.8 6.6-8.5 24.1-27.5 104.5-120.0 R 111.6±123.1 12.9-15.8 28.8-32.0 7.8-10.4 6.6-7.Э 25.2-28.1 109.1-120.1 Р < 0.05 |
|||||||
Таблица 8. Промеры бедренных костей усть-полуйских (F) и современных (R) орланов-белохвостов Haliaeetus albicilla
|
Статистики |
Промеры |
|||||
|
1 |
2 |
3 |
4 |
5 |
||
|
п |
F |
11 |
15 |
12 |
13 |
13 |
|
R |
7 |
7 |
6 |
7 |
7 |
|
|
Х + S.E. |
F |
115.46±0.834 |
25.80±0.392 |
9.97±0.115 |
25.98±0.375 |
22.21±0.391 |
|
R |
121.07±1.693 |
26.64±0.682 |
10.58±0.248 |
27.33±0.571 |
22.30±0.255 |
|
|
lim |
F |
112.5-120.9 |
24.1-28.8 |
9.5-10.7 |
24.1-29.0 |
20.4-25.1 |
|
R |
116.0-129.8 |
25.1-30.7 |
9.9-11.7 |
26.1-30.7 |
21.2-23.1 |
|
|
Р |
< 0.01 |
< 0.05 |
||||
Беркут Aquila chrysaetus (L.)
Материал: cran — 7 (7), premax — 1, mand — 5 (5), cor — 1, ulna — 4 (3), rad — 1, syns — 3 (2), fem — 4 (2), tib — 1, tmt — 1.
К.М.Дерюгин (1898), И.Н.Шухов (1915), Л.H.Добринский (1959) и Н.Н.Данилов (1965) эту птицу для среднего и нижнего течения Оби и южного Ямала не упоминают. Известны весенние и летние встречи беркутов на юге Ямала в 1970-1980-х (Данилов и др. 1984; Калякин 1989). В 1982 отмечено гнездование двух пар на р. Щучьей (Калякин 1989).
Полевой лунь Circus cyaneus (L.)
Материал: tmt — 2 (1).
Немногочислен. Гнёзда находили на р. Щучьей (Шухов 1915; Данилов и др. 1984). Пара отмечена 13 июля 1961 в низовьях Полуя (Бойков 1965); неоднократно полевые луни встречались на р. Собь (Добринский, 1965).
Таблица 9. Промеры элементов скелета усть-полуйских (F) и современных (R, подвид L. т. pleskei) тундряных куропаток Lagopus mutus
|
Кость, её промеры и с |
латистики |
F |
R |
|
Cranium GB |
n |
5 |
12 |
|
X± S.E. |
25.7±0.06 |
24.8±0.20 |
|
|
lim |
25.6-25.9 |
23.6-26.2 |
|
|
Coracoideum Gif |
n |
7 |
7 |
|
X± S.E. |
38.9±0.37 |
38.2±0.61 |
|
|
lim |
37.7-39.8 |
36.0-40.8 |
|
|
Humerus GL |
n |
67 |
16 |
|
X± S.E. |
59.3±0.20 |
58.4±0.35 |
|
|
lim |
55.7-62.9 |
56.2-60.1 |
|
|
Ulna GL |
n |
22 |
14 |
|
X± S.E. |
55.4±0.15 |
56.2±0.45 |
|
|
lim |
53.1-57.7 |
53.2-58.5 |
|
|
Carpometacarpus GL |
n |
1 |
16 |
|
X± S.E. |
33.0 |
32.7±0.26 |
|
|
lim |
33.0 |
30.8-34.3 |
|
|
Femur GL |
n |
20 |
13 |
|
X± S.E. |
54.1 ±0.37 |
52.9±0.49 |
|
|
lim |
51.5-57.1 |
50.7-57.1 |
|
|
Tibiotarsus La |
n |
8 |
12 |
|
X± S.E. |
72.2±0.60 |
68.4±0.87 |
|
|
lim |
70.2-75.8 |
64.4-75.2 |
|
|
Tarsometatarsus GL |
л |
1 |
13 |
|
X± S.E. |
33.0 |
33.3±0.36 |
|
|
lim |
33.0 |
30.6-35.0 |
Сапсан Falco peregrinus Tunst.
Материал: hum — 1, tmt — 1.
Редкая птица. В окрестностях Салехарда сапсана наблюдал только К.М.Дерюгин (1898) 19 августа 1897. Ближайшее известное место гнездования — р. Щучья (Данилов и др. 1984).
Тундряная куропатка Lagopus mutus (Montin)
Материал: cran — 6, cor — 8, hum — 68 (36), ulna — 22, rad — 1, cmcl, fem — 20, tib — 8, tmt — 1.
Промеры черепа и костей поясов конечностей представлены в таблице 9. Сравнение с современными экземплярами L. mutus pleskei с северо-восточного Ямала и материковой Якутии показало, что усть-полуйские тундряные куропатки по средним значениям измеренных параметров несколько крупнее современного подвида. Однако они не имеют значимых отличий от современных ямальских форм, размеры которых несколько больше, чем якутской формы. Кроме того, на Усть-Полуйском городище найдено несколько костей североуральского подвида L. mutus comensis, который, по данным промеров шкурок, отличается от равнинных птиц очень мелкими размерами (см.: Потапов 1985). Это плечевые (GL, мм: 55.7, 55.7, 56.4) и локтевая (GL, мм: 53.1) кости от двух особей.
Ближайшие от Салехарда места гнездования этого вида — Уральские горы и северная часть полуострова Ямал (Добринский 1965; Потапов 1985). Отмечены редкие встречи птиц в низовьях Полуя (Бойков 1965). В коллекции ЗИН РАН имеются экземпляры из "подножия Урала против Обдорска" (самка sad, 20 августа 1897, К.М.Дерюгин) и окрестностей Салехарда (самец, 28 января 1909, Д.Я.Вардроппер). Северосибирскому подвиду, в отличие от других, свойственны регулярные сезонные перемещения, носящие на некоторых территориях, в т.ч. Ямале, характер настоящих перелётов (Потапов 1985), при которых птицы отлетали массовыми стаями южнее, занимая на зимовках территории низовий Оби.
Белая куропатка Lagopus lagopus (L.)
Материал: cran — 31, mand — 2, cor —51, scap — 10, hum — 269 (144), ulna — 91, rad — 3, cmc — 6, fem — 111, tib — 44, tmt — 42.
Данные по промерам основных костей скелета представлены в таблице 10. Усть-полуйские птицы несколько уступают размерами современным материковым белым куропаткам номинального подвида, добытым в Хибинских горах, районе Салехарда и северной Якутии. При сравнении голоценовых костей с современными экземплярами только из района Салехарда (и = 4) видно, что размеры их почти одинаковы. Учитывая большой размах изменчивости подвида, его клинальную направленность (Потапов 1985) и скудность остеологических коллекций, проводить сравнительный статистический анализ голоценовых и современных форм преждевременно. Белая куропатка обычна около Салехарда. В низовьях Оби особенно многочисленна поздней осенью и ранней весной (Дерюгин 1898; Бойков 1965).
Таблица 10. Промеры элементов скелета усть-полуйских (F) и современных (R) белых куропаток Lagopus lagopus
|
Кость, её промеры и статистики |
Усть- полуйские |
Современные |
||
|
L. 1. lagopus |
L 1. pallasi |
|||
|
Салехард |
Все |
|||
|
Cranium GB п |
16 |
1 |
9 |
4 |
|
Х± S.E. |
26.8±0.20 |
25.7 |
27.5±0.28 |
26.3±0.45 |
|
Пт |
25.7-28.6 |
25.7 |
25.7-28.7 |
24.9-27.4 |
|
Coracoideum Gif п |
47 |
- |
14 |
4 |
|
Х± S.E. |
40.7±0.21 |
- |
41.4±0.27 |
39.9±0.14 |
|
Пт |
37.9-45.3 |
- |
39.7-42.8 |
39.5-40.3 |
|
Humerus GL п |
262 |
3 |
25 |
6 |
|
Х± S.E. |
61.9±0.12 |
61.1±1.07 |
62.7±0.36 |
61.0±0.68 |
|
Пт |
57.5-68.9 |
59.3-63.7 |
59.9-65.6 |
58.7-64.1 |
|
Ulna GL п |
91 |
4 |
19 |
7 |
|
Х± S.E. |
57.9±1.02 |
57.7±0.66 |
59.0±0.45 |
59.2±0.33 |
|
Пт |
55.5-61.9 |
56.1-59.8 |
55.2-63.0 |
58.1-60.8 |
|
Carpometacarpus GL п |
6 |
4 |
19 |
7 |
|
Х± S.E. |
34.4±0.38 |
33.9±0.47 |
34.7±0.23 |
34.1±0.21 |
|
Пт |
33.3-35.9 |
32.5-35.1 |
32.5-37.5 |
33.2-34.4 |
|
Femur GL п |
111 |
4 |
22 |
4 |
|
Х± S.E. |
58.1±0.17 |
57.9±0.41 |
59.4±0.42 |
57.2±0.70 |
|
Пт |
54.3-62.9 |
56.8-59.1 |
54.2-62.5 |
55.7-59.4 |
|
Tibiotarsus La п |
44 |
2 |
20 |
3 |
|
Х± S.E. |
76.8±0.36 |
75.7 |
77.4±0.39 |
75.6±0.56 |
|
Пт |
72.7-81.4 |
74.6-76.8 |
73.8-80.6 |
76.1-78.8 |
|
Tarsometatarsus GL п |
40 |
4 |
17 |
7 |
|
Х± S.E. |
39.4±0.21 |
40.0±0.50 |
40.1±0.31 |
40.2±0.24 |
|
Пт |
36.9-42.4 |
38.6-41.3 |
37.7-42.3 |
39.5-41.3 |
Белая или тундряная куропатка Lagopus ex gr. L. lagopus-L, mutus
Материал: cran — 5, vert — 1, syns —11, stern — 96, pelvis — 2/1.
По неполным черепам и их фрагментам, а также по вышеперечисленным остаткам, разделить кости по видам оказалось невозможным. Этот материал включён в рис. 5а.
Глухарь Tetrao urogallus L.
Материал: cran — 1, mand — 1, cor — 1, stern — 1, hum — 3 (2), rad — 1, cmc — 7 (6), tmt — 1.
В настоящее время северные территории Палеарктики населяет один из самых крупных подвидов — северотаежный глухарь Tetrao urogallus obsoletus Snigirewski, 1937 (Потапов 1985). Из-за отсутствия его посткра- ниального скелета в остеологической коллекции ЗИН сравнения проводились с Т. и. uralensis Nasarov, 1887 и Т. и. pleskei Stegmann, 1926.
Среди остатков из Усть-Полуйского городища самцам принадлежит 10 костей (5 особей), самкам — 6 (1 особь). Обломок нижней челюсти происходит от относительно мелкого самца (табл. 11), возможно, от особи в возрасте 1.5-2 лет (по материалам из Печоро-Илычского заповедника, Но, мм: 12.3, однолеток; 12.5-13.6, 2-5 лет). Кости плечевого пояса (cor, hum, cmc) по своим средним и абсолютным размерам очень крупные (табл. 11). Их значения превышают, либо лежат на границе максимальных значений современных экземпляров Т. и. uralensis (cor (GL) 87.6-93.4 мм; hum (Bp) 32.6-35.5; cmc (GL) 68.2-70.8) и T. u. pleskei (hum (Bp) 23.7-25.6 mm; rad (GL) 116.5-120.6; cmc (GL) 69.2-70.2). Это даёт основание полагать, что остатки глухаря принадлежат крупной форме, вполне соответствующей по своим размерам Т. и. obsoletus.
Возможно, в окрестностях Салехарда глухарь гнездится. К.М.Дерюгин (1898) встречал его в долине Оби к северу только до 66° с.ш. А.М.Гын-газов и Н.Г.Шубин (1968) сообщают, что в Западной Сибири глухари проникают в лесотундру, где местами многочисленны. В.Н.Бойков (19656) отмечает гнездование глухарей в "Приобской лесотундре". В коллекции ЗИН РАН имеется молодой самец, добытый у Салехарда 28 января 1909. К западу от Урала, в Канино-Тиманской лесотундре, глухарь регулярно гнездится и зимует (Михеев 1948).
Таблица 11. Промеры черепа и нижней челюсти усть-полуйских (F) и современных (R; Т. и. obsoletus) глухарей Tetrao urogallus
|
Статистики |
Промеры |
||
|
Cranium |
Mandibula |
||
|
GB |
Lp |
Но |
|
|
п F Самцы 1 Самки 1 1 R Самцы 555 Самки 5 5 5 X+S.E. F Самцы - - 12.9 Самки 39.2 45.6 R Самцы 51.010.51 59.810.71 13.210.32 Самки 38.710.33 44.810.30 8.010.10 lim F Самцы - - 12.9 Самки 39.2 45.6 R Самцы 49.8-52.9 57.2-61.3 12.5-14.2 Самки 38.0-40.1 38.0-46.0 7.7-8.3 |
|||
Тетерев Lyrurus tetrix (L.)
Материал: hum — 1. Остатки принадлежат крупной самке (табл. 12).
По К.М.Дерюгину (1898), тетерев по Оби к северу немного не дохо- дит до Салехарда. Н.Н.Данилов (1975) считает возможным гнездование примерно на широте 65°. В долинах рек Полуй и Собь отмечается как редкая птица (Бойков 1965а; Добринский 1965).
Таблица 12. Промеры плечевых костей усть-полуйских (F) и современных (R; L. t. tetrix) тетеревов Lyrurus tetrix
|
Статистики |
Промеры |
|||
|
GB |
Вр |
Bd |
SC |
|
|
п F |
1 |
1 |
1 |
1 |
|
R Самцы |
5 |
5 |
5 |
5 |
|
Самки |
10 |
10 |
10 |
10 |
|
Х + S.E. F |
84.6 |
22.9 |
16.0 |
8.2 |
|
R Самцы |
83.410.44 |
21.910.22 |
15.610.15 |
8.110.89 |
|
Самки |
74.210.47 |
19.510.10 |
16.910.10 |
7.210.05 |
|
lim F |
84.6 |
22.9 |
16.0 |
8.2 |
|
R Самцы |
82.1-84.6 |
21.0-22.4 |
15.0-16.0 |
7.8-8.3 |
|
Самки |
71.8-76.3 |
18.8-20.0 |
13.3-14.3 |
7.0-7.4 |
Таблица 13. Промеры локтевых костей усть-полуйских (F) и современных (R) серебристых чек Larus argentatus
|
Статистики |
Промеры |
|||
|
1 |
2 |
3 |
||
|
п |
F |
7 |
7 |
7 |
|
R |
20 |
20 |
20 |
|
|
XI S.E. |
F |
151.3612.294 |
10.8710.406 |
11.9010.257 |
|
R |
151.8611.162 |
11.5410.113 |
12.6810.156 |
|
|
Пт |
F |
140.0-161.0 |
9.8-13.0 |
11.0-13.3 |
|
R |
142.0-163.6 |
10.6-12.2 |
11.9-13.1 |
|
|
Р |
< 0.01 |
|||
Серый журавль Grus grus (L.)
Материал: cor — 1.
Редкий залётный вид. Северная граница ареала проходит "несколько южнее Салехарда" (Флинт 1987, с. 271).
Стерх Grus leucogerunus Pall.
Материал: cran — 1, mand — 1, cor — 1, ulna — 2 (1), tmt — 1.
В бассейне Оби достоверно гнездится между реками Питляр и Куно-ват и южнее р. Куноват (Сорокин, Котюков 1982). Неоднократно наблюдали этих журавлей севернее — в ряде пунктов того же Шурышкарского района (Брауде 1982), недалеко от Салехарда (Азаров 1977), на юге Ямала (Брауде 1982; Данилов и др. 1984).
Серебристая чайка Larus argentatus Pontopp.
Материал: premax — 1, cor — 1, stern — 4 (4), hum — 19 (10), ulna —18 (9), rad - 6 (4), cmc - 7 (5), fem - 3 (2), tib - 3 (3), tmt - 3 (3).
Измерения локтевых костей показали значимые различия в ширине дистального эпифиза между более мелкими ископаемыми чайками и более крупными современными (табл. 13). Для сравнения использовались кости трёх северных подвидов современных серебристых чаек (по: Юдин, Фирсова 1988): L. a. argentatus, L. a. antelius, L. a. taimyrensis.
Обычный гнездящийся вид.
Филин Bubo bubo (L.)
Материал: cran — 1, hum — 1, fem — 1, tib — 1.
В бассейне Оби гнездится на широте пос. Мужи (65°30' с.ш.) и южнее; изредка осенью и зимой залетает на южный Ямал (Добринский 1959; Данилов и др. 1984). Известна встреча филина в августе 1976 на р. Щучьей (Калякин 1989).
Белая сова Nyctea scandiaca (L.)
Материал: cran — 2 (2), quadr — 1, stern — 1, ulna — 2 (1), cmc — 2 (1), tmt — 4 (2).
По сообщению К.М.Дерюгина (1898, с. 107), "в северных тундрах и под Обдорском белая сова обыкновенна, особенно осенью и зимой". Другие исследователи летом у Салехарда эту сову не отмечали. По-видимому, она встречается здесь только на кочёвках.
Бородатая неясыть Strix nebulosa J.R.Forst.
Материал: ulna — 1.
Летом в низовьях Оби эту сову отмечал только Финш, который с помощью местных жителей 10 июля 1876 нашёл молодых птиц близ Уст-ремских юрт (Дерюгин 1898; Шухов 1915). Зимой встречается до южного Ямала (Данилов и др. 1984). Наблюдалась 30 июня 1979 в пойме Танлова-яхи, левом притоке Щучьей (Калякин 1979). Л.С.Степанян (1990) проводит северную границу гнездовой части ареала в бассейне Оби по 67° с.ш.
Серая ворона Corvus cornix L.
Материал: hum — 2 (2), ulna — 4 (3), cmc — 2 (2), fem — 1, tib — 2 (2).
Обычная гнездящаяся птица.
Ворон Corvus corax L.
Материал: cran — 1, premax — 1, mand — 1, clav — 1, cor — 2 (2), hum — 2 (2), fem — 1, tib — 1, tmt — 1.
Редкая гнездящаяся птица.
Обсуждение
Все виды, установленные по ископаемому материалу, уже отмечались в нижнеобском регионе исследователями XIX-XX вв. Но среди птиц Усть-полуйского городища много форм, характерных для более южных районов, встречающихся сейчас у Салехарда как редкие залётные или редкие гнездящиеся, находящиеся у северных пределов своего распространения (чомга, кряква, большой крохаль, тетеревятник, беркут, глухарь, тетерев, серый журавль, филин, бородатая неясыть). Не исключено, что 2000 лет назад границы областей гнездования птиц были несколько смещены к северу. По палеоботаническим данным, в это время на севере Западной Сибири климат был более тёплым (Волкова и др. 1989), край лесной полосы проходил значительно севернее и в районе устья реки Полуй росли сосновые леса (Мошинская 1953).
Интересные результаты дало измерение костей. Мы промерили локтевые кости краснозобой и чернозобой гагар, бедренные кости и тибио-тарсусы белолобого гуся и гуменника, бедренные кости и карпометакар-пусы орлана-белохвоста, по восемь основных элементов скелета тундряной и белой куропаток, череп и нижнюю челюсть глухаря, локтевые кости серебристой чайки. Ископаемые формы, кроме гусей, тундряной куропатки и глухаря, оказались в среднем немного мельче современных. Правда, эти различия статистически значимы только для общей длины бедра, высоты основания головки бедра и общей длины карпометакарпу-са орлана-белохвоста и ширины дистального эпифиза локтевой кости серебристой чайки. Но за такое короткое для эволюции время (2 тыс. лет) трудно ожидать более существенных изменений. По крайней мере, можно говорить о тенденции к укрупнению костей у ряда видов в рассматриваемый период. У гусей, наоборот, ископаемые птицы были более крупными по сравнению с современными.
Комплекс птиц Усть-Полуйского городища представлен 7 группами. В основном это куриные и водно-болотные птицы (всего более 92% особей), а также дневные хищные, совы и врановые (табл. 14). Видно, что в первую очередь добывались птицы, имеющие промысловое значение или имеющие отношение к определенным культурным традициям населения.
Из воробьиных в небольшом числе присутствуют только врановые, которых могли привлекать пищевые отбросы на поселении. На голоценовых местонахождениях, не связанных с деятельностью человека, воробьиных значительно больше — 30-97% (Барышников и др. 1986; Пантелеев 1993).
Кости куриных составляют 56.2% от всех костей птиц. Подавляющее большинство из них принадлежит белой куропатке — виду, как правило многочисленному среди остатков на памятниках позднего палеолита Северного Урала и Среднего Приуралья (Потапова 1990, 1991), лесной зоны и некоторых памятников лесостепной зоны Русской равнины (комплекс Костенок, Новгород-Северский, Мезин — Зубарева 1950; данные О.Р.По-таповой), южной Сибири (Афонтова Гора III — Тугаринов 1932).
Среди археологических местонахождений голоценового возраста остатки белых куропаток редки. В местонахождении Маяк 2 (ранний бронзовый век, Кольский п-ов) они составляют около 1% костных остатков птиц (Potapova, Sablin 1994; данные О.Р.Потаповой). На Северном Урале они известны из стоянки Канинская на Верхней Печоре, датированной бронзо-железным веком (Кузьмина 1971).
Таблица 14. Распределение остатков птиц из Усть-Полуйского городища по систематическим группам
|
Группа |
видов |
Число: |
% особей |
|
|
остатков |
особей |
|||
|
Gaviidae, Podicipitidae, Charadriidae, Laridae |
5 |
147 |
30 |
7.1 |
|
Anatidae |
19 |
488 |
142 |
33.7 |
|
Accipitridae, Falconidae |
6 |
181 |
22 |
5.2 |
|
Tetraonidae |
4 |
1122 |
217 |
51.4 |
|
Gruidae |
2 |
7 |
2 |
0.5 |
|
Srtrigidae |
3 |
17 |
4 |
0.9 |
|
Corvidae |
2 |
22 |
5 |
1.2 |
|
Aves indet. |
- |
12 |
- |
- |
|
Всего |
41 |
1996 |
422 |
100.0 |
Кости двух видов Lagopus представлены почти всеми элементами скелета. Среди последних большинство составляют плечевые и бедренные кости (см. рис. 5а). Их доминантное положение среди остальных элементов скелета характерно для местонахождений с остатками куропаток на поселениях человека, таких как Абри Фонталес, Эббу и Ле Коломбьер во Франции (Mourer-Chauvire 1983). Находки черепов, грудины и тазовых костей, наряду с другими элементами скелета, свидетельствуют о том, что утилизация птиц происходила на поселении. Сохранность почти всех костей очень хорошая, среди длинных элементов скелета целые кости составляют 88%. Подавляющее большинство костей принадлежит взрослым особям: от молодых птиц происходит лишь 1.5% (на эпифизах обнажена губчатая ткань). На многих костях (плечевых, бедренных) имеются чёткие следы зубов (1.3 % от общего числа остатков куропаток) в виде ямок с вмятыми вовнутрь обломками кости или небольших отверстий (1.6-3.0 мм, 2.0-3.7 мм, 2.7-2.9 мм) на эпифизах. Некоторые кости (0.4%) несут следы насечек на диафизе (бедренная) и сквозных ножевых порезов разной глубины на эпифизах (плечевые). Интересно, что черепа куропаток жителями Усть-Полуя были специально собраны (Адрианов 19366).
Белые куропатки — стайные птицы и держатся вместе большую часть года, за исключением сезона размножения. Объединение выводков в табунки в северных районах ареала происходит в августе. В низовьях Оби табунки с максимальной численностью птиц (80-100 особей) наблюдались в октябре (Бойков 1965). Поздней осенью в тундре начинаются зимние миграции белых куропаток в лесотундровую и север лесной зоны, области, в которых уровень численности птиц настолько высок, что позволяет вести в настоящее время промысловую охоту (Потапов 1985). Осенне-зимний период, по-видимому, являлся основным периодом промысла для усть-полуйского населения.
Белые куропатки привлекали человека, видимо, не только высокой численностью и гастрономическими качествами, но и сравнительной легкостью добычи, не требующей применения стрел или других трудоёмких, но легко утрачиваемых предметов и приспособлений. Известно множество различных способов безружейной массовой добычи этих птиц, например, силками в загородках, слопцами, пастями, сетями, шатрами, вентерями, кымрами, сачками и др. (Силантьев 1898; Колосов, Шибанов 1957). В недавнем прошлом эти способы лова были широко распространены в северных районах. В XIX в. А.А.Силантьев (1898, с. 367) отмечал, что "до последнего времени многие промышленники добывали белых куропаток, совсем не имея даже ружей". Интересно, что ещё сравнительно недавно в тундрах Ленско-Хатангского междуречья и на нижней Колыме практиковался весенний лов куропаток сетями (Романов 1934; Е.Р.Потапов, устн. сообщ.). Технология изготовления сетей из крапивного и тальникового волокон была хорошо известна всем народам, занимающимся рыболовством, со времен неолита-бронзы (Косарев 1987, 1991; История Дальнего Востока... 1989). Поэтому не исключено, что в условиях комплексного хозяйства, характерного для Усть-Полуя, сети могли использоваться весь год: летом — для ловли рыбы и линяющих гусей, а в осенне-зимний период — для ловли куропаток (так поступает, например, местное население нижней Колымы; Е.Р.Потапов, устн. сообщ.). Однако возможно, что для лова куропаток изготавливался и специальный инвентарь. Наряду с наиболее выгодным осенне-зимним промыслом в районе поселения, в отдаленных районах в течение любого времени года куропатки могли забиваться палками, либо охотники просто могли ловить птиц руками, используя специфическую особенность в поведении этих птиц в малонаселенных человеком областях, где они могут подпускать человека на расстояние 2-3 м, а также весной в токовый период (Потапов 1985).
Остатки крупных тетеревиных — глухаря и тетерева — принадлежат взрослым особям обоих полов. Как и на многих других стоянках человека севера Русской равнины, они обычно редки и значительно уступают по числу остатков белым куропаткам.
Вторую важную группу по промыслу составляют пластинчатоклювые, преимущественно гуси, лебеди и речные утки. Их присутствие легко объяснить расположением городища у большой реки — важной пролётной трассы водоплавающих птиц. Их могли добывать как во время гнездования, так и в период линьки и миграций. Судя по числу особей, предпочтение отдавалось чирку-свистунку, белолобому гусю, гуменнику и свиязи (по 8-34% от всех особей семейства). Белолобый гусь мог добываться только на пролёте, так же как и тундряный лебедь.
Интересны находки на городище большого числа костей и особей гуменника — немногочисленной здесь в настоящее время птицы. Редко добывалась шилохвость, которая является ныне в низовьях Оби очень многочисленной.
Соотношение элементов скелета лебедей, белолобого гуся, гуменника, свиязи и шилохвости сходно с белыми куропатками (рис. 5 б, в). Но у ряда
О 5 10 15 20 25 30 35 40
О 5 10 15 20 25 30
cran mand scap clav cor stern hum ulna rad cmc vert pelvis fem tib tmt phal
а
Lagopus
О
5 10 15 20 25 30 35 40
cran mand scap clav cor stern hum ulna rad cmc vert pelvis fem tib tmt phal
б
-
■ Cygnus bewickii
-
□ Cygnus cygnus
О
5 10 15 20 25 30 35
40 45
О
cran
mand
scap clav
cor
stern
hum
ulna
rad
cmc
vert
pelvis fem
tib
Anas penelope
tmt
phal
5 10 15 20 25 30
55 60 65 70 75 80 85
Anas platyrhynchos
Anas crecca
Anas acuta
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85
О 5 10 15 20 25 30 35 40
cran в- mand г scap 1
clav cor □ stern □= hum i ■' .
ulna r-1*™"***-^
rad r—B?—1 cmc vert pelvis fem □=□ tib i■ tmt c=i
0 5 10 15 20 25 30 35 40
-
■ Gavia stall ata
-
□ Larus argentatus
0 5 10 15
cran ■■ mand ■■ © scap ^ clav cor stern ■ hum ^—^—« ulna rad ^■n"^^^*
cmc vert pelvis fem tib —i^— tmt ^——
Phal^--,--
0 5 10 15
1 Haliaeetus albicilla
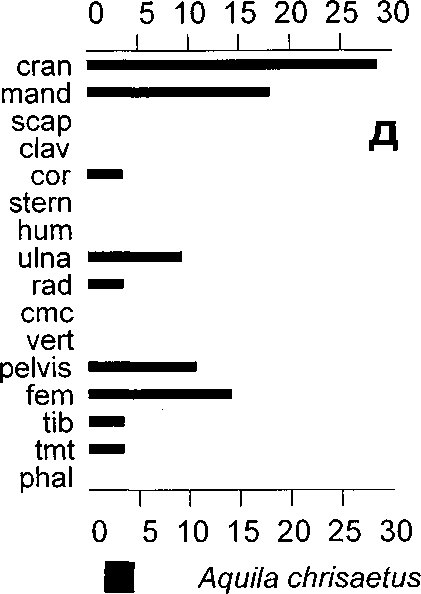
Рис. 5. Соотношение (в %) между сохранившимися элементами скелета у некоторых видов птиц из Усть-Полуйского городища.
В категорию “Cranium” включены остатки верхнечелюстных костей.
крупных гусеобразных либо отсутствуют (лебеди), либо присутствуют в очень небольшом числе (белолобый гусь, гуменник) грудные кости. Грудина, как и некоторые другие несъедобные остатки дичи, использовалась для различных хозяйственных нужд. На поселении обнаружены "ложки", сделанные из грудных костей пластинчатоклювых (4 экз.), гагар (И экз.) и крупных ястребиных (беркутов или орланов-белохвостов; 2 экз.). У грудин аккуратно обрезали киль, закругляли края и зачищали все выступающие на наружной стороне неровности, что придавало изделию ложкообразный вид (см.: Мошинская 19536, табл. XII, 1). Но в передней части грудины пластинчатоклювых имеется естественное отверстие, уходящее в утолщение киля, которое, при срезании последнего, становится сквозным. В костях без естественных отверстий люди просверливали искусственные отверстия, обычно в передней, реже в боковой части. На одной из грудин гагары при срезании киля была оставлена передняя утолщенная часть в виде рукоятки, направленной вниз и вперед, хотя такое расположение очень затрудняет зачерпывание. Очевидно, что все эти "ложки" использовали не для еды, а для каких-то ритуальных целей. В.И.Мошинская (1953) считает, что ложкообразные грудные кости, наряду с лопаточкообразными деревянными ложками, представляют наиболее архаичную форму изделий. Манси ещё недавно употребляли костяные ложки-лопаточки при жертвоприношениях и при снаряжении покойника в потусторонний мир. В ненецком фольклоре центральный персонаж сказки о Порнэ-не — полуженщина-полумедведица — употребляет вместо ложки грудную кость "лебедя" (Там же).
Остатки ряда видов (шилохвость, чирки) представлены, главным образом, дистальными частями конечностей, преимущественно крыльями. Известно, что современное население северных территорий использует крылья некоторых утиных, несущих ярко окрашенные кроющие второстепенных маховых, для украшения домашней утвари и одежды. Во многих деревенских домах России крылья птиц применялись для намазывания масла на сковороду при выпечке блинов, а на нижней Колыме, кроме того, в качестве веника для подметания полов (Е.Р.Потапов, устн. сообщ.). В прошлом крылья шилохвости, чирков и широконоски могли также использоваться в декоративных, ритуальных и других целях.
Немногочисленность остатков гагар и серебристых чаек — обычных птиц побережий, озер и рек лесотундровой зоны, так же как и нырковых уток и крохалей, свидетельствуют о второстепенном характере их добывания. Соотношение элементов скелета этих видов, сходное с белыми куропатками (рис. 5 г), даёт возможность предполагать их полную утилизацию на поселении. Из гагар, возможно, добывали жир, который был всегда большой ценностью для населения Севера, а шкурки (красивые, тёплые, прочные) могли идти на пошив одежды (Колосов и др. 1983). Так, население на Внешних Гебридах и Оркнейских островах (стоянки Юдал и Ваккой), добывало гагар попутно с олушей исключительно ради жира с начала I тыс. н. э. до средневековья (см.: Serjeantson 1988). Остатки чаек на многих известных стоянках мезолита-раннего средневековья северного побережья Скандинавии и Шотландии обычно редки или ма- лочисленны (Serjeantson 1988; A.K.Hufthummer, pers. comm.). Являясь перелётными, чайки и гагары могли добываться в районе городища только весной и летом, с мая по август (Флинт 1982; Юдин, Фирсова 1988).
Остатки сов принадлежат трём крупным видам, но преобладает белая сова. Вероятно, на сов, как и на куропаток, существовал специальный промысел. Так, по сообщению К.М.Дерюгина (1898), осенью и зимой, когда белые совы под Салехардом особенно многочисленны и сильно жиреют, местные жители искусно ловили их, употребляя как лакомство в пищу, а из крыльев делали опахала от комаров. На Ямале самоеды (ненцы) ловили белых сов капканами, состоящими из высокого шеста и петли (Шухов 1915), на Присаду и на гнёздах (Житков 1912). Для индейцев Великих Равнин совы были символом сверхъестественной мистической силы; им также приписывали лекарственные свойства. Их содержали в неволе как предсказателей событий, а перья (особенно виргинского филина) использовали для специфических церемоний и танцев. Некоторые племена использовали в пищу не всех сов, а только определённые виды. Так, среди остатков 6 видов сов на стоянках арикара порезы на костях были найдены только у виргинского филина (Parmalee 1977а).
На жертвенном месте городища найдено довольно большое число костей и особей орланов-белохвостов и беркутов. Среди остатков беркутов оказалось необычно много черепов, в числе которых присутствует аккуратно срезанная верхняя часть черепной коробки. Орланы представлены почти всеми частями скелета (рис. 5 д, е) и, в отличие от беркутов, видимо, захоранивались целыми. Разное соотношение элементов скелета может свидетельствовать о различном использовании этих видов.
Среди костей орланов-белохвостов оказались голень и локтевая кость от 1 или 2 особей (из 10, найденных на поселении), которые были сломаны и срослись при жизни птицы. В природе птицы, имеющие повреждения скелета, редки. Так, среди остатков из Усть-Полуйского городища подобный случай ещё отмечен на одной бедренной кости белой куропатки (0.5% от числа особей куропаток) и одной голени белолобого гуся (2% от особей гусей). В изученной части коллекции (несколько тысяч костей) из бинагадинских асфальтов, естественной ловушки среднего плейстоцена, были обнаружены только 2 особи птиц (кряква и степной орёл) с зажившими переломами костей (Бурчак-Абрамович 1949, 1968). Однако среди утиных зажившие повреждения костей конечностей могут встречаться чаще (13% из 256 скелетов уток: Музей Естественной Истории, Университет Канзаса; Tiemeier 1941).
Хищной птице со сломанными крылом и ногой выжить особенно трудно или почти невозможно — для этого необходимы особо благоприятные условия (обилие легко добываемой пищи, отсутствие врагов и др.). Эти обстоятельства свидетельствуют о том, что на поселении люди могли содержать в неволе некоторых птиц (по крайней мере, орланов). Интересно, что среди 145 скелетов ара, содержащихся в неволе для ритуальных целей индейцами Пуэбло, остатки которых найдены на стоянках в Аризоне и Нью Мексико, 13% особей имели зажившие травмированные конечности (Brothwell 1993).
Содержание в неволе диких птиц, в том числе хищных, широко практиковалось у многих народов. "Орлов”, взятых птенцами, держали селькупы, кеты, айны (Соколова 1972; Косарев 1981, 1991). Последние выращивали "орлов" при доме для принесения их в жертву. У ряда племён индейцев Калифорнии хищные птицы содержались в неволе для приманки при организации охоты на орлов, являющейся значительным ритуальным событием и для всех индейцев Великих Равнин (Parmalee 1977b). Беркут и белоголовый орёл, играющие наиболее важную роль в ритуальных церемониях индейцев Великих Равнин, были найдены в 60% из 51 стоянок племени арикара в Южной Дакоте и составили около 9% от общего числа остатков птиц (Parmalee 1977а,b).
Были и другие цели содержания хищных птиц. Михалон Литвин, путешествуя в первой половине XVI в. по юго-западной Руси, записал: "Орлят запирают в клетки для перьев к стрелам" (Аристов 1866, с. 6-7).
У многих народов, в т.ч. сибирских, существовал культ орла. Орёл так или иначе ассоциировался с солнцем, то отождествляясь с ним, то представляясь его хозяином, и в качестве такового творцом, верховным божеством, либо священной птицей верховного божества, и являлся благодетелем и покровителем как отдельных людей, так и целых родов, “птицей счастья” (Штернберг 1925, с. 718). У обских угров тотемический характер культа часто перекликался с представлением о душе-птице (четвёртой душе человека), живущей на голове в волосах (Чернецов 1959; Косарев 1981). Скульптурное изображение орла часто встречается на бытовых предметах и культовом оружии из Усть-Полуйского городища: гребнях, пряжках, подвесках, ложках, рукоятке ножа, клевцах (Мошинская 1953; Чернецов 1953а). На 9 предметах из 12, имеющих изображение птиц, в реалистическом и стилизованных стилях изображены орёл или орлы, клюющие голову лося (Адрианов, Архив МАЭ, с. 727-729, К-1, оп. 2). У целого ряда сибирских народов месяц март называется “месяцем орла”.
Солнечная птица Карс (орёл) являлась важнейшей частью обряда, посвященного "верхнему" небесному миру. Этот обряд очень древний, и его происхождение связывают с Индией или югом Ирана (Штернберг 1925; Чернецов 1947). Небесный Карс должен сидеть на священном мировом дереве, на котором растут солнце и луна. В условиях Сибири мировое дерево заменялось берёзой или лиственницей.
В.Н.Чернецову (1959) удалось присутствовать при обряде инициации в роде Крылатого Старика (орла) на Оби. Юношу по достижении им определенного возраста вели на священное родовое место, где его заставляли влезть на дерево, на котором "обитает" крылатый предок рода. В XX в. н.э. потомки обских угров вполне обходятся без реальной птицы. Но в древности, когда традиции, религия, шаманство были более сильны, на дереве вполне мог сидеть живой орлан или беркут.
Являясь тотемной птицей, орёл у многих сибирских народов считался неприкосновенным. У якутов мёртвый орёл, как сородич и покровитель, хоронился. Нечаянно убивший орла человек должен был съесть его всего, кроме головы, поджарив на углях (Штернберг 1925). Возможно, большое количество черепов беркутов, собранных на поселении, косвенно указы- вают на какой-то отбор определенных частей птицы, при котором часть посткраниальных скелетов беркутов могла не сохраниться.
Находки целых скелетов и большого количества черепов "орлов" в Усть-Полуйском городище документально свидетельствуют о существовавшем культе этих птиц у обских угров, об их жертвоприношении (сохранялись большей частью головы), и возможно, захоронении, по крайней мере со П-1 вв. до н.э., что вполне согласуется с современными этнографическими данными о традиционных верованиях ряда народов, связанных с культом орла.
