Правые и левые формы ели в потомствах плюсовых деревьев в Пермском крае

Автор: Рогозин Михаил Владимирович, Голиков Анатолий Матвеевич, Комаров Святослав Александрович
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Ботаника
Статья в выпуске: 2, 2012 года.
Бесплатный доступ
Обнаружено, что в потомстве плюсовых деревьев из естественных насаждений преобладают левые (светолюбивые), а в потомстве из искусственных насаждений - правые (теневыносливые) формы. Потомства первых и вторых ценопопуляций, при разделении их на группы с положительной и отрицательной реакцией на повышение густоты выращивания, имеют различное соотношение этих форм и разный эволюционный ответ на изменение густоты выращивания в испытательных культурах, выравнивая доли правых и левых форм.
Ель сибирская и финская, плюсовые деревья, потомство, правые и левые формы
Короткий адрес: https://sciup.org/147204596
IDR: 147204596 | УДК: 630.5(075),
The right and left forms of spruce progenies of plus trees in the Perm region
It was found that the progeny of plus-trees from natural stands dominated by leftists (love of light-tions), and progeny from plantations - right (shade-tolerant) form. First and second offspring of populations, with the division into groups with positive and negative-term response to the increasing density of cultivation, have a different relationship between these different forms and the evolutionary response to changes in density of growth in test cultures, aligning the proportion of left and right forms.
Текст научной статьи Правые и левые формы ели в потомствах плюсовых деревьев в Пермском крае
О дисимметрии в потомствах сосны и ели
Известно, что у сосны и ели к признакам, связанным с конкурентной выносливостью и продуктивно-стью,следует отнести альтернативные фены – правое (D) и левое (L) вращение филлотаксиса (мест прикрепления хвои) на стволовом побеге, которое получило название «энантиоморфы» [Хохрин,1977].
Так, изучение 28-летних плантационных культур ели с густотой посадки 1 и 4 тыс. экз. на 1 га показало, что в густых посадках правые формы превосходили левые по высоте на 7–8 % и по объему ствола – на 33%. В редких посадках, наоборот, левые формы имели лучшие показатели по высоте на 4–7% и по объему ствола на 10–23%. Поэтому при низкой плотности популяции преимущества имеют L-формы, а при высокой – D-формы. Их адаптивная разнокачественность, по-види-мому, связана со световой конкуренцией, и левые формы как более светолюбивые имеют преимущества в разреженном древостое, а D-формы как более теневыносливые – в густых насаждениях. Предполагается, что L-формы лучше используют прямой солнечный свет и более сухие условия, а D-формы – рассеянный свет и влажные условия [Голиков, Бурый, 2008].
Материнские деревья ели в высокополнотных насаждениях чаще продуцирует потомство с преоб- ладанием правых форм, которые наиболее адаптированы к повышенной густоте произрастания. В потомстве деревьев, произрастающих в низкопол-нотных древостоях, наоборот, чаще встречаются левые формы [Голиков, 2010].
Объекты и методы
В наших исследованиях в 2011 г. изучена частота этих форм в 24-летних потомствах плюсовых деревьев ели на одном из двух участков испытательных культур, в кв. 41 Ильинского лесничества, где посадки проводились на суглинистой почве по схеме 2.5×1.0 м. Рост этих же потомств изучали еще на одном участке культур в Юго-Камском лесничестве, где посадку проводили на супесчаной почве и более густо, с размещением растений в раскорчеванных полосах тремя рядами по схеме 0.7×0.7 м.
Среди 453 семей ели, представленных в этих парных испытаниях потомства, по результатам их роста в 21-23 года, были выделены три группы по их реакции на повышение густоты выращивания (плотности стояния):
-
1 – положительная: увеличение средней высоты в густых культурах до 115% от контроля при средней высоте в редких культурах 92%;
-
2 – отрицательная: понижение высоты в густых культурах в среднем до 91% от высоты в редких культурах 113%;
-
3 – семьи-универсалы, растущие одинаково хорошо и в редких, и в густых культурах (116 и 113% от контроля соответственно).
Общая численность всех групп составила 146 семей. Остальные 307 семей проявили не столь контрастные и стабильные реакции. Значительная часть семей представляет собой потомство плюсовых деревьев из культур Ф.А. Теплоухова.
Для изучения филлотаксиса хвои в натуре мы выбирали одно нормально развитое растение из семьи и фотографировали со штатива последний годичный прирост на вершинке дерева; номер семьи записывали в блокнот и сразу снимали его на фото. В камеральных условиях по характеру закрутки хвоинок определяли форму дерева (правая или левая) и последствия повреждения кроны вредителями в баллах: 0 – повреждения нет; 1 – центральный побег искривлен и снизил прирост; 2 – центральный побег погиб и замещен.
При расчете достоверности различий между долями использована формула
-
tф Т ( Р 2 Н p 1 )/ V m 22 6 m 2 2 2 ,
где
m p Т
Р 2 (1 Н Р ) n Н 1
n – числорастений;
р – долялевыхформ(илидоляповрежденных побегов).
Результаты и их обсуждение
Результатысведенывтабл.1,изкоторойследу-ет,чтоестественныепопуляциипроизводятпотом-ковсповышеннойдолей(57%)левых(светолюби-вых)форм,акультуры–потомковспреобладани-ем (61.3%) правых (теневыносливых) форм. Раз-личиестатистическидостоверно t ф =14при t 0.01 =2.7.
Таблица1
Доля правых и левых форм ели в испытательных культурах
|
Ценопопуляция |
Числорасте-ний,шт. |
L-формы, % |
D-формы, % |
Повреждения крон,%. |
|
Естественные:Гайва,Пермь,Очер |
64 |
57.0 |
43.0 |
75 |
|
Культуры:Вещагино,Сепыч1,Сепыч2,Очер |
82 |
38.7 |
61.3 |
68 |
Полученные различия в преобладании левых формвпотомствеестественныхнасажденийипра-выхформвпотомствекультурТеплоуховасоот-ветствуютусловиямразвитияихродителей:весте-ственных древостоях в относительно редком, а в культурахТеплоухова–вболееплотномстоянии. Однаковкультурахуплюсовыхдеревьевбылиоб-наруженывсреднемна8.3%болеедлинныешиш-кивсравнениисшишкамидеревьевизестествен-ныхдревостоев,ипоэтомупризнакуельвкульту-рах Теплоухова более «европейская», а в есте-ственныхдревостоях,гдешишкикороче,ельболее «сибирская»[Рогозин,Разин,2011].Поэтомуобъ-яснить только плотностью популяций либо при-надлежностьюихкразнымвидамелиразличияв дисимметрииихпотомковпоканельзя.
В исследованиях А.М. Голикова [2004, 2007] выяснено, что в больших репрезентативных вы-боркахизсотендеревьеввпопуляцияхсоотноше-ниеLиD-формстремитсяк1:1,аихчисленные колебаниявразныхценозахинасажденияхневы-ходятзапределыот38до62%.Этойжезаконо-мерностиподчиняетсяварьированиелевыхипра-вых энантиоморф каквпределахразныхсемей сосныобыкновеннойиелиевропейской,такив потомстверазличныхпопуляцийэтихвидов.Дан-наянорматесносвязанасизвестнымвсимметрии правиломзолотогосеченияилизолотойпропорци-ей,гдеменьшаячастьотноситсякбольшей, как большаякцелому,т.е.0.38197:0.61803=0.61803: (0.38197+0.61803)или38:62=62:(38+62).Эта нормаявляетсяважнымкритериемдлябиомонито- ринга на семейном ипопуляционномуровне.На нарушение равночисленного соотношения энан-тиоморфвпотомстведостоверноевлияниеоказы-вает уровень гетерозиготности и экологические условияпроизрастанияматеринскихдеревьев.Ве-роятно,соотношениеихвпотомствезависитиот густотыдревостоев[Голиков,1985,2004,2007].
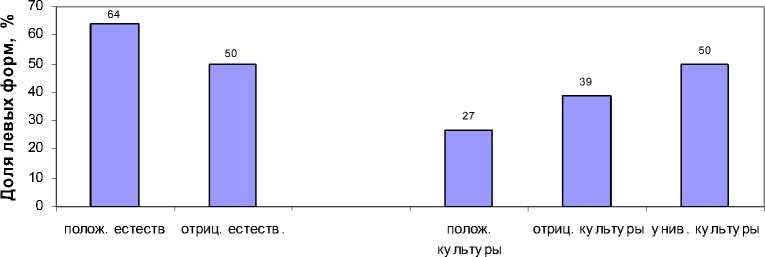
Унасполучилисьследующиерезультаты(рис.1):
1.ВпотомствекультурТеплоухова(справатри столбика)потомствасположительнымотношени-ем кплотнойпосадке имеют низкую (27%), а потомствасотрицательнойреакциейнагустоевы-ращивание–болеевысокуюдолюлевыхформ (39%),чтосогласуетсясвыводамиА.М.Голикова [2010].
-
2 . В потомстве естественных ценозов (слева) картинаиная:потомства–любителиплотноговы-ращивания имеют высокую долю левых форм (64%), апотомствасотрицательнойреакциейна густоевыращивание–равночисленноесоотноше-ниеправыхилевыхформ(по50%).
-
3 .В потомстве из культур удалось выделить группусуниверсальнымбыстрымростомвобоих испытательных культурах, наиболее ценнуюдля селекции; впотомствахизестественныхпопуля-цийтакихуниверсальныхсемеймывыявилимало ианализироватьихнестали.Дляэтойгруппысе-мей-универсаловсоотношениеэнантиоморфоказа-лосьравным–50/50.
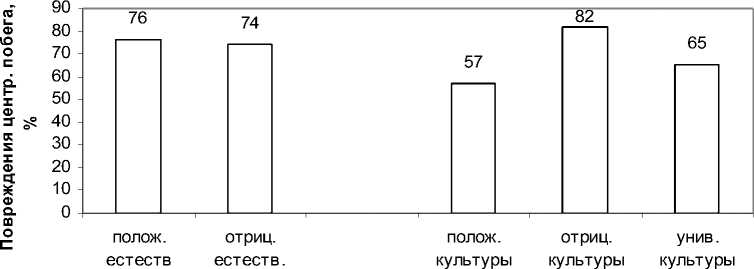
Повреждения центрального побега вредителя-мивпотомствеизкультурвсреднемнесколько ниже(68%),чемвпотомствеизестественныхна- саждений (75%). Различие статистически достоверно tф = 14 при t0.01 = 2.7 (рис. 2, табл. 2).
Гр уппы семей с разной реакцией на повышение густоты выращ ивания
Рис. 1 . Доля левых (светолюбивых) форм ели у семей из естественных и искусственных насаждений в группах семей с разной реакцией на повышение густоты выращивания в испытательных культурах
Группы се ме й с разной реакцие й на повышение густоты выращивания
Рис. 2. Повреждение центрального побега ели у потомств из естественных насаждений и из культур Теплоухова в группах потомств с разной реакцией на повышение густоты выращивания
Таблица 2
Различия между долями левых форм и повреждениями центрального побега в потомстве естественных популяций (группы 1, 2 и 3) и в потомстве культур (группы 4,5 6 и 7)
|
Группа |
Ценопопуляции |
Реакция на густоту |
Число растений, шт. |
Доля левых форм, р |
m p |
Сравниваемые группы |
Различие, t ф |
Повр. крон, доля р |
m p |
Сравниваемые группы |
Различие, t ф |
|
1 |
Гайва |
полож. |
33 |
0.64 |
0.007 |
1-2 |
9.3 |
0.76 |
0.005 |
||
|
2 |
Пермь, Очер |
отриц. |
31 |
0.50 |
0.008 |
0.74 |
0.006 |
- |
|||
|
3 |
Итого, в среднем |
64 |
0.57 |
0.007 |
0.75 |
0.004 |
3-7 |
14 |
|||
|
4 |
Верещагино |
полож |
15 |
0.27 |
0.014 |
4-5 |
5.7 |
- |
- |
||
|
5 |
Сепыч 1, 2; Очер |
отриц |
33 |
0.39 |
0.007 |
5-6 |
7.8 |
0.82 |
0.004 |
5-6 |
17 |
|
6 |
Сепыч 1 |
универс. |
34 |
0.50 |
0.007 |
4-6 |
10.9 |
0.65 |
0.006 |
||
|
7 |
Итого, в среднем |
82 |
0.387 |
0.002 |
0.68 |
0.003 |
Заключение
Таким образом, в потомстве естественных насаждений преобладают левые (светолюбивые), а в потомстве культур Теплоухова – правые (теневыносливые) формы. Потомства первых и вторых цено-популяций, при разделении их на группы с положительной и отрицательной реакцией на повышение густоты выращивания, имеют достоверно различное соотношение этих форм и разное направление эволюционного ответа на изменение густоты выращивания в испытательных культурах, выравнивая доли правых и левых форм. Семьи с универсальной способностью быстрого роста как в редких, так и в густых испытательных культурах имеют равные доли правых и левых форм (по 50%). Не исключено, что изменение численного равновесия энантиоморф – это ответная эпигенетическая реакция потомства на изменение густоты выращивания. Дальнейшие исследования с использованием дисимметрического подхода позволят понять механизм микроэволюции лесных популяций различной плотности.
Список литературы Правые и левые формы ели в потомствах плюсовых деревьев в Пермском крае
- Голиков А.М. Формы сосны обыкновенной и их селекционное значение в условиях Псковской области: автореф. дис. канд. с.-х. наук: 06.03.01.Свердловск, 1985. 18 с.
- Голиков А.М. Влияние экологических факторов на наследственную неравноценность дисимметричных форм плюсовых деревьев сосны обыкновенной//Тр. СПбНИИЛХ. 2004. Вып.2(12). С. 117-131.
- Голиков А.М. Рост и формовая структура потомства ели европейской в зависимости от гетерозиготности деревьев и условий произрастания//Лесоведение. 2007. № 4. С. 51-58.
- Голиков А.М., Бурый Н.Л. Влияние густоты посадки на рост и конкурентные отношения энантиоморф сосны и ели в 28-летних плантационных культурах//Рациональное природопользьвание и перспективы устойчивого развития лесного сектора экономики: материалы конф. Нов-ГУ им. Ярослава Мудрого. Великий Новгород, 2008. С. 78-81.
- Голиков А.М. Теоретическое и прикладное значение экологодиссимметрийного подхода в исследовании формовой и генетической структуры популяций видов хвойных//Наука о лесе ХХI века: материалы междунар. науч.-практ. конф. Гомель, 2010. С. 157-160.
- Рогозин М.В., Разин Г.С. Лесные культуры Теплоуховых в имении Строгановых на Урале: история, законы развития, селекция ели. Пермь, 2011. 192 с.
- Xохрин А.В. Внутривидовая диссимметрическая изменчивость древесных растений в связи с их экологией: автореф. дис. д-ра биол. наук: 03.00.05. Свердловск, 1977. 49 с.
