Предварительные данные по изменчивости двух микросателлитных локусов у соболя (Martes zibellina L.) Среднего Приамурья
 Среднего Приамурья")
Автор: Шишацкая Д.И., Фрисман Л.В.
Журнал: Региональные проблемы @regionalnye-problemy
Рубрика: Биология. Экология. Геоэкология
Статья в выпуске: 2 т.17, 2014 года.
Бесплатный доступ
Проведен анализ изменчивости микросателлитных локусов Ма-2 и Ма-3 в 5 географических выборках соболя таежной зоны хребтов Малый Хинган, Буреинский и Сихотэ-Алинь. Семь аллелей Ма-2 локуса и 4 аллеля Ма 3 локуса было обнаружено. Ожидаемая гетерозиготность в локальных выборках варьирует от 0,364 до 0, 801 (среднее - 0,620). Для обоих локусов в каждой из выборок (кроме Ма2 из «Каменушки») наблюдается соответствие равновесному состоянию по Харди-Вайнбергу. Уровень попарной генетической дифференциации по рассматриваемым микросателлитным локусам варьирует от 0,021 до 0,023. Nm =11,11, указывает на значительный миграционный поток между рассматриваемыми локальными сообществами. Однако в распределении некоторых аллелей обнаружена географическая изменчивость.
Соболь, популяция, генетическая изменчивость, микросателлиты
Короткий адрес: https://sciup.org/14328884
IDR: 14328884 | УДК: 575.174.015.3:
Preliminary data on genetic variation of the sable (Martes zibellina L., 1758) in the Middle Priamurie: the analysis of two microsatellite loc
The variation of microsatellite loci Ма 2 and Ма 3 was examined in 5 geographical samples of the sable Martes zibellina from the taiga zone of Smaller Khingan, Bureinsky and Sikhote-Alin ridges. It was found seven alleles in Ma-2 locus and four alleles - in Ma 3. The expected heterozygosity varies from 0.364 to 0.801 (on average - 0.620). In both loci of every geographical sample (except Ma 2 from “Kamenushka”) it is observed a correspondence to the equilibrium state according to the Hardy-Weinberg equilibrium. The level of pair-wise genetic differentiation F (ST) at two microsatellite loci varies from 0.021to 0.023. Nm = 11.11, that reflects a significant migration flow between these local communities. However, some alleles have a different distribution in the 5 geographical samples.
Текст научной статьи Предварительные данные по изменчивости двух микросателлитных локусов у соболя (Martes zibellina L.) Среднего Приамурья
Соболь (Martes zibellina Linnaeus, 1758) - один из самых ценных промысловых видов на территории России. На протяжении нескольких веков соболь по праву считается одним из национальных символов России [3].
В далеком прошлом соболь заселял территорию от Тихоокеанского побережья до Скандинавского полуострова. Высокая ценность пушнины соболя и стабильный на протяжении столетий спрос на нее привели к нерациональному промыслу и практически полному истреблению природных популяций. На европейском севере он исчез еще несколько веков назад. Распространение соболя к началу XX в. носило мозаичный характер. А к началу 30-х гг. соболь сохранился только в отдельных труднодоступных участках разрозненными очагами. Так в ряде регионов Западной Сибири он исчез полностью [8]. Одной из мер для сохранения этого вида явилась интродукция соболя на угнетенные территории. Около 27% - современного ареала соболя занимают популяции, образованные таким путем [1]. Искусственное расселение соболя во многих районах ареала вида -один из примеров успешного восстановления ресурсов ценных пушных зверей промысловой фауны страны [6]. Всего в Советском Союзе было выпущено около 20000 соболей. К концу XX в. численность соболя восстановилась практически на всем его ареале.
Современный ареал соболя охватывает таежную зону Евразии от северного Предуралья до Тихоокеанского побережья и прилежащих островов. Лишь незначительные по численности популяции этого вида существуют на территории Японии, Китая, Кореи и Монголии. Проведенная в прошлом веке массовая реаклиматизация, высокая хозяйственная значимость и широкое распрос транение соболя во всех зоогеографических таежных зонах Евразии обуславливают многогранность проводимых в настоящее время исследований. Так, краниометрические исследования [14] изменчивости соболя после 40 лет акклиматизации обнаружили, что по некоторым неметрическим признакам черепа особей из популяционных группировок акклиматизантов сходны с черепами автохтонных животных, а по другим признакам - с черепами животных исходных территорий. Математическое моделирование на основе динамики численности популяций позволило получить оценку оптимального промыслового изъятия [13]. Анализ ядерных и митохондриальных генов выявил генетическую изменчивость соболя, филогеографическую структуру и позволил получить характеристики межвидовой гибридизации [2, 9, 15]. Анализ микросателлитных локусов в промышленных популяциях соболя [4] показал перспективность привлечения этого метода для исследования генетической структуры современных природных популяций данного вида. Распределение аллелей микросателлитных локусов в природных популяциях [10, 11] обнаружило высокий уровень генетического популяционного разнообразия соболя Уральского региона - зоны симпатрии и гибридизации соболя и лесной куницы. Несомненно, что древность существования популяций в данном регионе и интродукция животных из Восточной Сибири также могли сказаться на уровне генетического разнообразия. Высокий уровень генетического разнообразия показан у соболя Сихотэ-Алиня. Обнаруженное в этом регионе разнообразие может иметь аборигенные корни, т.к. существование здесь лесного рефугиума в периоды плейстоценовых оледенений не подлежит сомнению. Другой источник разнообразия - инт- родукция животных из Предбайкалья, которая была проведена в прошлом веке. Пищулина с соавторами отмечают высокую степень генетических отличий между популяциями соболя Северного Урала и Камчатки, при существенно более низком уровне генетического разнообразия последней [10]. Популяции соболя в Центральной Сибири оказались достаточно однородны. В то же время, соболь западной части Хоккайдо подразделен на 2 генетически различающиеся группы, чьи ареалы не полностью изолированы [19].
В силу большой сложности получения материала, исследование природных популяций обычно опирается на суммарные выборки как в пространстве (по регионам), так и во времени. Такой подход дает усредненные характеристики внутривидового разнообразия сообщества конкретного региона, не позволяя отследить внут-рипопуляционные изменения, которые являются базой для эколого-генетического исследования. Настоящая работа - это первая попытка подробного исследования генетической популяционной структуры соболя Среднего Приамурья, в формирование которой внесли вклад как обитатели исходных природных сообществ региона, так и интродуценты.
Работа основана на анализе сборов одного сезона (зима 2012-2013 гг.). Главной целью настоящей работы является анализ генетической популяционной структуры соболя Среднего Приамурья. В связи с этим были поставлены следующие задачи:
-
1. Провести анализ двух микросателлитных локусов в выборках одного сезона для пяти географических точек Среднего Приамурья, локализованных в отрогах хребтов Малый Хинган, Буреинский и Сихотэ-Алинь.
-
2. На основе анализа полученных генетических коэффициентов оценить уровень популяционной изменчивости и дифференциации соболя в регионе.
Материалы и методы
В работе использованы образцы тканей от 131 соболя, добытых охотниками в зимний сезон 2012-2013 гг. в пяти географических районах Среднего Приамурья (рис. 1):
-
1. Еврейская автономная область, Облученский район, бассейн р. Каменушка - 36 образцов (далее «Каменушка»).
-
2. Хабаровский край, Верхнебуреинский район, верховья р. Сутырь - 24 образца (далее «Сутырь).
-
3. Хабаровский край, Верхнебуреинский район, пересечение р. Тырма и ее притоков Нигида и Копуны -28 образцов (далее «Тырма»),
-
4. Хабаровский край, Верхнебуреинский район, бассейн р. Маган - 13 образцов (далее «Маган»),
-
5. Хабаровский край, Нанайский район бассейн р. Манома -30 образцов (далее «Манома»),
ДНК выделяли из мышечной ткани с использованием стандартного метода солевой экстракции [16], включающего лизис клеток протеиназой К в присутствии додецилсульфата натрия, очистку ДНК хлоридом натрия и изопропиловым спиртом и осаждение этиловым спиртом. Количественное определение ДНК проводили при помощи спектрофотометра «НаноДроп 2000». Использовали амплификационную смесь следующего состава из расчета 10 pl на 1 образец (50-100 ng ДНК): 10 ПЦР буферКСГ, 1,5 мМ MgCl2, 0,2 мМ dNTP, 0,5 мМ прямой и обратный праймеры, 0,5u Taq ДНК полимераза (И.Л. Кац, личное сообщение).
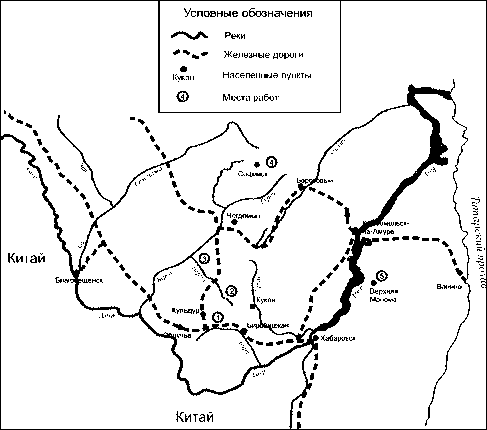
Рис. 1. Локализация мест сбора материала: 1 - «Каменушка», 2 - «Сутырь», 3 - «Тырма», 4 - «Маган», 5 - «Манома»
Амплификацию проводили в следующем режиме: 8 циклов, включающих 1 мин денатурации ДНК при температуре 94° С, отжиг праймеров 30 с- при Х° С и 30 с синтеза новых цепей при 72° С; 21 цикл, включающий 30 с при температуре 94° С, 30 с при Х° С и 15 с при 72° С; элонгация 3 мин при 72° С. X - температура отжига для индивидуальной пары праймеров (табл. 1). Участки ДНК, соответствующие выбранным локусам, амплифицировали с помощью праймеров, флуоресцентно меченных по 5'-концу красителем TAMRA производства ООО «Бигль» (Санкт-Петербург). Режим амплификации (с изменением Т отжига для Ма 2) и последовательности примененных праймеров взяты из рабо-
Таблица 1
Характеристика исследованных микросателлитных локусов
Фрагментный анализ проводили на восьмиканальном автоматическом генетическом анализаторе Applied Biosystems 3500 Genetic Analyzer с длиной капилляра 50 см, с использованием гель-полимера POP 7, в присутствии маркера молекулярного веса LIZ 600. Для идентификации аллелей использовали программу GeneMapper (ver. 4.1) «Applied Biosystems» (США). Оценку гетерозиготности и частот аллелей, а также статистические тесты на соответствие наблюдаемых частот генотипов по каждому локусу равновесному состоянию по Харди-Вайнбергу проводили с использованием программы GenePop V4 [12]. Построение UPGMA дендрограммы осуществляли с помощью программы Biosys-1 [18].
Результаты и их обсуждение
С целью изучения генетической структуры природной популяции соболя Martes zibellina в Среднем Приамурье проведено исследование изменчивости двух микросателлитных локусов Ма 2 и Ма 3. Полученные данные приведены в табл. 2. На 131 исследованную особь по двум микросателлитным локусам выявлено 11 аллелей. Среднее количество аллелей на локус 5,5.
Для обоих локусов в каждой из выборок (кроме Ма 2 из локалитета «Каменушка») наблюдается соответствие равновесному состоянию по Харди-Вайнбергу.
Как видно из табл. 2, в локусе Ма 2 обнаружено семь аллелей. Они представлены пятью гомозиготными (164164, 166-166, 168-168, 170-170 и 172-172) и семнадцатью гетерозиготными генотипами. Из возможных у семи аллелей двадцати одного гетерозиготного генотипа не были обнаружены четыре следующих: 162-172, 162-174, 170-174 и 172-174. В каждой из географичес ких выборок представлено по пять либо шесть аллелей. Пять из семи обнаруженных аллелей (164,166, 168,170, 172) оказались общими для всех рассматриваемых географических выборок. Наиболее часто во всех выборках, кроме самой малочисленной - «Маган», представлен аллель 164. При этом частота его встречаемости не превышает 0,361. Аллели 162 и 174 имеют мозаичное распределение с частотой встречаемости, не превышающей 0,083. В выборке «Сутырь» отсутствует аллель 162, а в выборках «Маган» и «Каменушка» отсутствует аллель 174.
Для локуса Ма 3 обнаружено четыре аллеля, представленных в трех гомозиготных (131-131, 133-133,135-135) и четырех гетерозиготных (129-131, 131-133, 131-135, 133-135) генотипах. В каждой из географических выборок представлено по три аллеля этого логоса. Географическое распределение аллелей логоса Ма 3 более упорядочено, чем у Ма 2. Аллель 131 локуса Ма 3 является основным в выборках соболей всех рассматриваемых локальностей, встречаясь здесь с частотой от 0,643 до 0,769. Стоит отметить, что выборка «Манома», наиболее географически удаленная от других точек и отделенная рекой Амур, маркирована собственным уникальным аллелем 129 с частотой встречаемости- 0,017. В то же время здесь отсутствует аллель 135, обнаруженный в выборках левобережья Амура с частотами 0,042-0,143.
Интересно, что количественный состав аллелей (семь - для Ма 2 и четыре - для Ма 3) в исследованной нами суммарной природной выборке совпадает с данными, полученными для промышленных популяций [5].
Для представления картины географической структурированности соболиного сообщества рассматриваемой территории были посчитаны роджеровские [17] генетические дистанции (табл. 3).
Таблица 2
Аллельный состав микросателлитных локусов, частоты встречаемости и показатели гетерозиготности соболя Среднего Приамурья
|
Локус, аллели Н0/НеР |
«Каменушка» |
«Сутырь» |
«Тырма» |
«Маган» |
«Манома» |
|
Ма2 (N) |
36 |
24 |
28 |
13 |
30 |
|
162 |
0,083 |
0,000 |
0,036 |
0,077 |
0,017 |
|
164 |
0,361 |
0,348 |
0,321 |
0,192 |
0,300 |
|
166 |
0,194 |
0,152 |
0,143 |
0,154 |
0,283 |
|
168 |
0,139 |
0,239 |
0,304 |
0,231 |
0,150 |
|
170 |
0,197 |
0,152 |
0,071 |
0,083 |
0,067 |
|
172 |
0,125 |
0,087 |
0,071 |
0,269 |
0,150 |
|
174 |
0,0000 |
0,022 |
0,054 |
0,000 |
0,033 |
|
н0/не |
0,611/0,786 |
0,625/0,768 |
0,787/0,775 |
0,769/0,801 |
0,867/0,778 |
|
Р |
0,0046 ± 0,0008 |
0,3378±0,0102 |
0,8597±0,0087 |
0,3445 ±0,0102 |
0,1374±0,0094 |
|
МаЗ |
|||||
|
129 |
0,000 |
0,000 |
0,000 |
0,000 |
0,017 |
|
131 |
0,667 |
0,604 |
0,643 |
0,731 |
0,767 |
|
133 |
0,250 |
0,354 |
0,214 |
0,154 |
0,217 |
|
135 |
0,083 |
0,042 |
0,143 |
0,115 |
0,000 |
|
н0/не |
0,500/0,486 |
0,458/0,484 |
0,464/0,535 |
0,385/0,429 |
0,400/0,364 |
|
Р |
0,1399±0,0145 |
0,1991±0,0733 |
0,3509±0,1258 |
1,0000 ±0,171 |
1,0000±0,0791 |
Примечание: N - объем выборки; Н /Н - наблюдаемая и ожидаемая гетерозиготность; Р - вероятность соответствия 0 гипотезе - соответствие распределению Харди-Вайнберга
Роджеровские [16] генетической дистанции соболя Среднего Приамурья
Таблица 3
|
Географические выборки |
«Каменушка» |
«Сутырь» |
«Тырма» |
«Манома» |
«Маган» |
|
«Каменушка» |
0,092 |
0,097 |
0,096 |
0,136 |
|
|
«Сутырь» |
0,104 |
0,130 |
0,166 |
||
|
«Тырма» |
0,147 |
0,130 |
|||
|
«Манома» |
0,152 |
UPGMA дендрограмма (рис. 2), построенная на основании этих дистанций, иллюстрирует наибольшее сходство между географически близкими выборками западной части ареала («Каменушка», «Сутырь», «Тырма») и позволяет предположить некоторую обособленность самой северной из четырех левобережных («Маган») и правобережной («Манома») как от группы западных выборок, так и между собой.
Дистанция
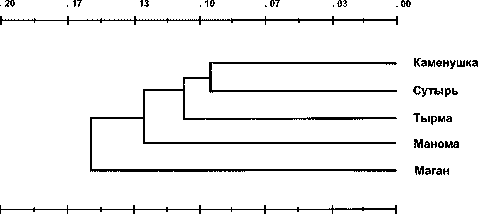
Рис. 2. UPGMA- дендрограмма соболя Среднего Приамурья
По рассматриваемым локусам, используя параметры F-статистик С. Райта [20], была определена степень подразделенности рассматриваемых локальных сообществ приамурского соболя. Значения этих параметров представлены в табл. 4.
Усредненные показатели величин F(IS) и F(IT) отражают незначительность доли инбридинга особи как по отношению к субпопуляции, к которой она принадлежит (F(IS)), так и к по отношению к сообществу всего исследованного региона (F(IT)). Более того, обнаруженная по локусу Ма 3 отрицательная величина F(IS) указывает на 1,5%-ный избыток гетерозиготных особей для этого локуса в среднем на субпопуляцию, по сравнению с теоретически ожидаемым при равновесном состоянии по Харди-Вайнбергу.
Таблица 4
Значения параметров F-статистик С. Райта по локусам Ма 2 и Ма 3 у соболя Среднего Приамурья (усредненная для 5 географических выборок)
|
Локус, среднее значение по двум локусам |
F(IS) |
F(IT) |
F(ST) |
|
Ма2 |
0,057 |
0,077 |
0,021 |
|
МаЗ |
-0,015 |
0,009 |
0,023 |
|
Среднее значение |
0,031 |
0,052 |
0,022 |
Показатель подразделенности F(ST), то есть уровень различий в генетической структуре исследованных локальных сообществ соболя варьирует незначительно: от 0,021 (Ма 2) до 0,023 (Ма 3). Среднее значение составляет 0,022 (2,2%). Следовательно, около 97,8% всего генетического разнообразия приамурского соболя можно найти внутри каждой субпопуляции и только 2,2% приходится на разнообразие между локальными сообществами.
-
С . Райт [20] использовал следующие градации для оценки значений F(ST):
-
- от 0 до 0,05 - малая генетическая дифференциация;
-
- от 0,05 до 0,15 - промежуточная дифференциация;
-
- от 0,15 до 0,25 - высокая степень генетической дифференциации;
-
- и более 0,25 - очень высокая степень генетической дифференциации.
В настоящем исследовании F(ST) = 0,022 (2,2%) отражает слабую степень генетической дифференциации субпопуляций соболя исследованных регионов Приамурья.
На основе полученных данных по F(ST) произведен расчет оценки потока генов Nm по формуле 1:
где N- эффективный размер популяции, а т - коэффициент миграции.
В результате расчета получили N m= 11,11 на поколение, что указывает на значительный миграционный поток между локальными сообществами рассматриваемой территории.
Итак, полученные значение показателя подразделенности (F(ST=0,022) и потока мигрантов (более 11 особей на поколение) позволяют рассматривать соболя лесных территорий хребтов Малый Хинган, Буреинский и Сихотэ-Алинь в рамках единой популяции. Вероятность соответствия структуры этой популяции равновесному состоянию по Харди-Вайнбергу, оцененное по суммарной выборке 5 локалитетов для локуса Ма 2=0,1 и для Ма 3=0,317. Обнаруженная географическая изменчивость аллельного состава и представленная в UPGMA дендрограмме картина взаимосвязей локальных сообществ указывают на ее некоторую пространственную структурированность.
Насколько стабильна обнаруженная структура популяции соболя Среднего Приамурья в пространстве и во времени покажут результаты дальнейших исследований.
Авторы выражают искреннюю благодарность Л.В. Капитоновой и П.В. Будилову за помощь в сборе материала, Л.Н. Спиридоновой за конструктивную и доброжелательную критику работы.
Настоящее исследование выполнено при частичной финансовой поддержке проектов № 12-1-ОБН-05 и 12-1-П30-14 Программы Фундаментальных исследований ДВО РАН «Дальний Восток».
Список литературы Предварительные данные по изменчивости двух микросателлитных локусов у соболя (Martes zibellina L.) Среднего Приамурья
- Грязнухин А.Н. Результаты реакклиматизации соболя в Якутии//Фауна и экология наземных позвоночных таежной Якутии. Якутск, 1980. С. 43-78.
- Жигилева О.Н., Политов Д.В., Головачева И.М., Петровичева С.В. Генетическая изменчивость соболя Martes zibellina L., лесной куницы M. martes L. И их гибридов в Западной Сибири: полиморфизм белков и ДНК//Генетика, 2014. Т.50, № 5. С. 581-590.
- Зырянов А.Н. Соболь Средней Сибири. Экология, промысел, охрана. Красноярск: «Сибирские промыслы», 2009. 240 с.
- Каштанов С.Н., Рубцова Г.А., Лазебный О.Е. Исследование генетической структуры промышленной популяции соболя (Martes zibellina Linnaeus,1758) по микросателитным маркерам//Вестник ВОГиС, 2010. Т. 14, № 3. С. 426-431.
- Каштанов С.Н., Афанасьев К.И., Потапов С.Г., Лазебный О.Е. Микросателлитный анализ двух промышленных популяций соболя (Martes zibelli-na L.)//Генетика. 2011. Т. 47, № 12. С. 1622-1628.
- Кириков С.В. Промысловые животные, природная среда и человек. М.: Наука, 1966. 348 с.
- Малярчук Б.А., Петровская А.В., Деренко М.В. Внутривидовая структура соболя (Martes Zibelli-na L.) по данным изменчивости нуклеотидных последовательностей гена цитохрома b митохондриальной ДНК//Генетика. 2010. Т. 46, № 1. С. 73-78.
- Монахов Г.И., Крючков В.С., Монахов В.Г., Шурыгин Б.В. Результаты интродукции восточносибирских соболей в Енисейской Сибири и бассейне р. Васюган//Промысловая териология. М.: Наука, 1982. С. 136-148.
- Петровская А.В. Генетическая структура популяций соболя (Martes zibellina L.) в Магаданской области по данным об изменении митохондриальной ДНК//Генетика. 2007. Т. 43, № 4. С. 530.
- Пищулина С.Л. Мещерский И.Г., Симакин Л.В., Каштанов С.Н., Лазебный О.Е., Рожнов В.В. Сравнительная оценка генетического своеобразия соболя на западе и на востоке ареала.//Междунар. совещ. IX Съезд Териологического общества при РАН. г. Москва, 1-4 февраля 2011 г. М., 2011. С. 371.
- Пищулина С.Л. Взаимодействие популяций лесной куницы и соболя в зоне симпатрии: генетический аспект: автореф. дис. … канд. биол. наук. Москва, 2013. 24 с.
- Раймонд М., Руссе Ф., 1995 GENEPOP (версия 1.2): Популяционная генетика: рабочая программа.
- Ранюк М.Н., Монахов В.Г. Изменчивость краниологических признаков в популяциях соболя (MARTES ZIBELLINA), возникших в результате акклиматизации//Экологический журнал. 2011. Т. 90, № 1. С. 82-96.
- Ревуцкая О.Л., Неверова Г.П., Фрисман Е.Я. Оценка оптимального промыслового изъятия охотничьих животных на территории Среднего Приамурья России//Региональные проблемы. 2008. № 9. С. 34-38.
- Рожнов В.В., Пищулина С.Л., Мещерский И.Г., Симакин Л.В., Лазебный О.Е., Каштанов С.Н. Генетическая структура соболя (Martes zibelli-nа L.) Евразии -анализ распределения митохондриальных линий//Генетика. 2013. Т. 49, № 2. С. 251-258.
- Aljanabi SM, Martinez I. Universal and rapid salt-extraction of high quality genomic DNAb for PCR-based techniques//Nuclec Acids Research. 1997. Vol. 25, No 22. Р. 4692-4693.
- Rogers J.S. Measures of genetic similarity and genetic distance//Studies in Genetics VII. Univ. Texas Publ, 1972. Vol. 7213. P. 145-153.
- Swofford D.R., Selander R.B. Biosys-1: a FORT-RAN program for the comprehensive analysis of electrophoretic data in population genetic and syste-matic//J. Heredity. 1981. Vol. 72, № 4. P. 281-283.
- Tomoko Nagai, Takahiro Murakami and Ryuichi Masuda. Genetic Variation and Population Structure of the SableMartes zibellina on Eastern Hokkaido, Japan, Revealed by Microsatellite Analysis/Mammal study, 2012. Vol.37. P. 323-330.
- Wright S. Evolution and the genetics of population/Variability within and among Natural Populations. Chicago: University of Chicago Press, 1978. Vol.4. 590 p.
