Предварительные результаты изучения взаимосвязи структуры кариотипа комаров-звонцов рода Chironomus (Diptera, Chironomidae) с факторами среды в условиях Центрального Кавказа
 с факторами среды в условиях Центрального Кавказа")
Автор: Кармоков Мухамед Хусенович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Общая биология
Статья в выпуске: 2-2 т.18, 2016 года.
Бесплатный доступ
Представлены предварительные результаты изучения взаимосвязи структуры кариотипа комаров-звонцов рода Chironomus (Diptera, Chironomidae) с факторами среды в условиях Центрального Кавказа. Полученные данные позволяют заключить, что наибольшая степень связи структуры кариотипа прослеживается с такими факторами среды как температура середины апреля, июля, температура среднегодовая, лесообразующая порода, высота над уровнем моря и прозрачность воды. Разные генотипические сочетания обнаруживают различную степень корреляции с факторами среды. В целом структура кариотипа демонстрируют существенную отрицательную корреляцию с температурными факторами среды.
Политенные хромосомы, хромосомный полиморфизм, окружающая среда, температурный режим
Короткий адрес: https://sciup.org/148204494
IDR: 148204494 | УДК: 595.771:574.52
Preliminary results of the study of interrelation of karyotype structure of non-biting midges of genus Chironomus (Diptera, Chironomidae) with environment factors in the Central Caucasus
The preliminary results of the study of interrelation of karyotype structure of non-biting midges of genus Chironomus (Diptera, Chironomidae) with environmental factors in the conditions of the Central Caucasus are presented. The obtained data allow us to conclude that the largest degree of interrelation of karyotype structure can be traced to such environment factors as, temperature in the middle of April and July, t average annual, forest-forming species, altitude and transparency of water. Different genotypic combinations exhibit varying degrees of correlation with environmental factors. In general, karyotype structure shows a significant negative correlation with temperature factors.
Текст научной статьи Предварительные результаты изучения взаимосвязи структуры кариотипа комаров-звонцов рода Chironomus (Diptera, Chironomidae) с факторами среды в условиях Центрального Кавказа
Личинки комаров-звонцов, являясь важнейшим компонентом бентоса, вызывают большой интерес и с теоретической, и с практической точек зрения. При благоприятных условиях они могут преобладать над другими донными беспозвоночными как наиболее ценный кормовой объект для бентосоядных промысловых рыб [1]. Также известна другая важная роль личинок хирономид в донных сообществах, как активных участников процессов самоочищения.
Экологическая кариология – недавно возникшее направление науки и, несмотря на большое теоретическое и прикладное значение, пока мало разработанное [2]. Исследование реакции генома на воздействие факторов среды весьма актуально при проведении генетического мониторинга природных популяций в условиях роста антропогенной нагрузки на водные экосистемы и для реализации комплексного подхода в эволюционном анализе популяций. Уже накоплены обширные сведения по выявлению зависимости уровня инверсионного полиморфизма личинок хирономид, особенно видов рода Chironomus , от экологических факторов различной природы: глубины обитания [3, 4], кислородного и температурного режимов водоема [3, 5-8], солености воды [7, 9, 10], природных и антропогенных химических факторов [11-14]. Исследования действия температуры на структуру и функциональную активность политенных хромосом хирономид проводились в разные сезоны года [15-17], в условиях вечной мерзлоты Исландии, Гренландии, арктической Аляски, Финляндии, Якутии, севера России [18-22]. Изучение инверсионного полиморфизма позволило оценить степень и характер воздействия на кариотип хирономид антропогенных факторов, в частности радиации, вызывающей появление в кариофонде видов редких последовательностей, сохраняющихся в ряду поколении [20]. Также было обнаружено, что отсутствие серьезных вариаций полиморфизма в природных популяциях Chirono-mus plumosus (Linnaeus, 1758) , Ch. balatonicus Devai et al., 1983 и Camptochironomus tentans (Fabricius), 1805 [23] указывало на устойчивость структуры кариотипа в ряду поколении [20]. Также было обнаружено, что отсутствие серьезных вариаций
полиморфизма в природных популяциях Chirono-mus plumosus (Linnaeus, 1758) , Ch. balatonicus Devai et al., 1983 и Camptochironomus tentans (Fabricius), 1805 [23] указывало на устойчивость структуры кариотипа в ряду поколений и отсутствие факторов, провоцирующих проявление дифференциальной адаптивности генотипов.
В большей части предыдущих работ модельным видом исследований был избран голарктически распространенный вид Ch. plumosus . В нашем же исследовании таким видом был выбран близкородственный вид Ch. nuditarsis Str. (Keyl, 1961) , так как у него, в отличие от Ch. plumosus , на Центральном Кавказе гораздо более широкий высотный диапазон распространения ( Ch. nuditarsis – 204-1789 м над ур. м.; Ch. plumosus – 183-558 м над ур. м.) [24] и, следовательно, возможно, более разнообразными проявлениями взаимосвязи кариотипа c факторами среды. Стоит также отметить, что данный вид ранее не использовался в качестве модельного.
Цель работы: выявить существует ли взаимосвязь структуры кариотипа Ch. nuditarsis с факторами среды в условиях Центрального Кавказа.
Материал и методы . Для проведения исследования были определены 4 стации, где по ранее опубликованным данным [24] обитает Ch. nuditarsis : 1. В черте с. Белая Речка, заболоченный ручей под пологом леса на первой надпойменной террасе, 43025’24.23’’, 43028’41.29’’, 736 м. над ур. м.; 2. Верхнее Голубое озеро, 43013’31.71’’, 43033’43.59’’, 913 м. над ур. м.; 3. 4-5 км ниже п. Верхний Баксан, Комсомольское озеро, 43019’32.58’’, 42048’19.40’’, 1454 м. над ур. м.; 4. В черте п. Эльбрус, долговременная лужа, подпитываемая подпорными водами р. Баксан, 43014’54.34’’, 42038’12.72’’, 1789 м. над ур. м. Первые две стации относятся к терскому, а вторые две - к эльбрусскому варианту поясности в соответствии с концепцией высотно-поясной структуры А.К. Темботова [25].
Отбор проб проводили с помощью гидробио-логичекого скребка, параметры 34х18х48 см. Прозрачность воды и глубина водоема определялась с помощью диска Секки. Температуру воды измеряли в середине каждого сезона года, во второй декаде января, апреля, июля и октября, с помощью цифрового термометра. При проведении исследований также учитывали характер грунта дна водоема и трофность. Личинок для кариологического анализа фиксировали в смеси 96% этилового спирта и ледяной уксусной кислоты (3:1). Давленые препараты политенных хромосом готовили по рутинной этил-орсеиновой методике [17, 26]. Исследование препаратов хромосом проводили с применением микроскопа проходящего света Carl Zeiss Axio Imager.A2. Для статистического анализа (кластерный анализ) использовался пакет программ
STATISTICA 10. Использовали непараметрические методы статистики, т.к. полученные данные не соответствовали нормальному распределению. Генетические дистанции между изученными популяциями вычислялись по критерию Нея [27]. Картирование плеч хромосом A, E и F проводили по Кейлу [28], а для плеч C и D по Деваи с соавторами [29]. Для выявления степени связи особенностей кариотипа с факторами среды определяли частоты различных генотипические сочетания в четырех выбранных стациях.
Таблица 1. Данные измерений параметров среды в 4-х стациях исследований Центрального Кавказа за 2015г.
|
Стации / температуры воды |
toC янв. |
toC апр. |
toC июль |
toC окт. |
toC сред./ год |
Тип раст. |
Высота над ур. м. |
Макс глуб. м |
Прозр воды, м |
Троф-ность |
Грунт дна |
|
п. Эльбрус |
+1,7 |
+11,2 |
+15,3 |
+12,3 |
+10,1 |
сосна |
1789 |
0,52 |
до дна |
мезо-троф |
серый ил |
|
Комсооль-ское озеро |
+2,5 |
+16,5 |
+20,3 |
+14,2 |
+13,3 |
береза |
1454 |
3-4 |
>3 |
олиго-троф |
желтый ил |
|
Верхнее Голубое озеро |
+1,2 |
+12,5 |
+16,5 |
+10,3 |
+10,2 |
ольха |
913 |
8-9 |
0,7 |
мезо-троф |
черный ил и листвен. опад |
|
Белая речка |
+3,1 |
+13,2 |
+17,5 |
+10,1 |
+10,9 |
ольха бук |
736 |
0,45 |
до дна |
мезо-троф |
черный ил и листвен. опад |
Таблица 2. Частоты генотипических сочетаний последовательностей дисков Ch. nuditarsis 4-х популяций Центрального Кавказа (n – количество изученных особей)
|
Популяции/ Генотипические сочетания |
ndt A1.1 |
ndt A1.2 |
ndt B1.1 |
ndt B1.2 |
ndt B2.2 |
ndt C1.1 |
ndt D1.1 |
ndt E1.1 |
ndt F1.1 |
ndt F1.2 |
ndt F2.2 |
ndt G1.1 |
ndt G1.2 |
ndt G2.2 |
|
п. Эльбрус, n=20 |
1 |
0 |
0,400 |
0,550 |
0,050 |
1 |
1 |
1 |
0,300 |
0,400 |
0,300 |
0 |
0 |
1 |
|
Комсомольское озеро, n=36 |
0,889 |
0,111 |
0,611 |
0,361 |
0,028 |
1 |
1 |
1 |
0,111 |
0,528 |
0,361 |
0,361 |
0,389 |
0,250 |
|
Верх. Голубое озеро, n=20 |
1 |
0 |
0,950 |
0,050 |
0 |
1 |
1 |
1 |
0,150 |
0,200 |
0,650 |
0,700 |
0,300 |
0 |
|
Белая речка, n=17 |
1 |
0 |
0,764 |
0,118 |
0,118 |
1 |
1 |
1 |
0,118 |
0,588 |
0,294 |
0,588 |
0,176 |
0,236 |
Таблица 3. Данные расчета ранговой корреляции Спирмена между показателями среды и генотипическими сочетаниями.
|
Параметры среды/ Генотип. сочетания |
A1.1 |
A1.2 |
B1.1 |
B1.2 |
B2.2 |
C1.1 |
D1.1 |
E1.1 |
F1.1 |
F1.2 |
F2.2 |
G1.1 |
G1.2 |
G2.2 |
|
t январь |
0,316 |
-0,738 |
0 |
-0,400 |
-0,400 |
-0,258 |
-0,258 |
-0,258 |
0,200 |
-0,800 |
0 |
0 |
-0,600 |
-0,400 |
|
t апрель |
-0,316 |
-0,949 |
-0,800 |
-0,400 |
-0,400 |
-0,775 |
-0,775 |
-0,775 |
-0,200 |
-0,800 |
-0,800 |
-0,800 |
-1,000 |
-0,400 |
|
t июль |
-0,316 |
-0,949 |
-0,800 |
-0,400 |
-0,400 |
-0,775 |
-0,775 |
-0,775 |
-0,200 |
-0,800 |
-0,800 |
-0,800 |
-1,000 |
-0,400 |
|
t октябрь |
0,316 |
0,105 |
-0,400 |
0,800 |
0,800 |
0,258 |
0,258 |
0,258 |
0,600 |
0,400 |
-0,400 |
-0,400 |
-0,200 |
0,800 |
|
t среднегод. |
-0,316 |
-0,949 |
-0,800 |
-0,400 |
-0,400 |
-0,775 |
-0,775 |
-0,775 |
-0,200 |
-0,800 |
-0,800 |
-0,800 |
-1,000 |
-0,400 |
|
лесообраз. порода |
-0,947 |
-0,316 |
-0,400 |
-0,800 |
-0,800 |
-0,775 |
-0,775 |
-0,775 |
-1,000 |
-0,400 |
-0,400 |
-0,400 |
-0,200 |
-0,800 |
|
высота над ур. м. |
0,632 |
0,632 |
0,200 |
1,000 |
1,000 |
0,775 |
0,775 |
0,775 |
0,800 |
0,800 |
0,200 |
0,200 |
0,400 |
1,000 |
|
маским. глубина |
-0,632 |
0,211 |
-0,600 |
0,200 |
0,200 |
-0,258 |
-0,258 |
-0,258 |
-0,400 |
0,400 |
-0,600 |
-0,600 |
0,000 |
0,200 |
|
прозрачность |
-0,500 |
-0,389 |
-0,949 |
0,105 |
0,105 |
-0,544 |
-0,544 |
-0,544 |
-0,211 |
-0,105 |
-0,949 |
-0,949 |
-0,632 |
0,105 |
|
трофность |
0 |
0,544 |
0,775 |
-0,258 |
-0,258 |
0,333 |
0,333 |
0,333 |
-0,258 |
0,258 |
0,774 |
0,774 |
0,775 |
-0,258 |
|
характер грунта |
-0,500 |
0,056 |
0,316 |
-0,739 |
-0,739 |
-0,272 |
-0,272 |
-0,272 |
-0,738 |
-0,211 |
0,316 |
0,316 |
0,316 |
-0,738 |
Примечания : параметры среды, демонстрирующие наибольшую степень связи с показателями хромосомного полиморфизма выделены полужирным шрифтом, p <0,05.
Результаты исследований. Были получены предварительные данные по динамике температуры воды в течение года, а также глубине, прозрачности и трофности исследованных водоемов (табл. 1). В рамках исследования были изучены характеристики структуры кариотипа и хромосомного полиморфизма личинок
Chirono-mus nuditarsis из 4-х стации исследования (табл. 2). Данные по п. Эльбрус взяты из ранее опубликованной работы [24]. Для выявления возможной связи параметров среды и частот различных генотипических сочетаний в программе Staistica 10 произвели расчет ранговой корреляции Спирмена между показателями среды и генотипическими сочетаниями (табл. 3). Также на основе данных о частотах генотипических сочетаний личинок Ch. nuditarsis из 4-х стации были рассчитаны величины генетических дистанций между указанными популяциями по критерию Нея [27] (табл. 4.).
Таблица 4. Величины генетических дистанций, между изученными популяциями вычисленные по критерию Нея [27]
|
Белая речка |
п. Эльбрус |
Комсомол. озеро |
Верх. Голубое озеро |
|
|
Белая речка |
0 |
|||
|
п. Эльбрус |
0,127 |
0 |
||
|
Комсомол. озеро |
0,020 |
0,094 |
0 |
|
|
Верх. Голубое озеро |
0,036 |
0,219 |
0,054 |
0 |
Используя полученные данные о величинах генетических дистанции между изученными популяциями, нами была построена дендрограмма генетических дистанции по методу ближайшего соседа в евклидовом пространстве (рис. 1).
Обсуждение результатов. Полученных данных в настоящий момент недостаточно для того, чтобы говорить о выявленных закономерностях, так как измерения параметров среды проводились только в течение одного года, однако, можно сделать некоторые предварительные выводы:
-
1. Наибольшая степь связи структуры кариотипа прослеживается с такими факторами среды как, t середины апреля, июля, t среднегод., лесообразующая порода, высота над уровнем моря и прозрачность воды.
-
2. Частоты разных генотипических сочетаний обнаруживают различную степень корреляции с факторами среды. В целом структура кариотипа демонстрируют существенную отрицательную корреляцию с температурными факторами среды.
-
3. Частоты генотипических сочетания имеют существенную положительную корреляцию с комплексом факторов связанных с высотой расположения над уровнем моря пункта исследования.
-
4. Между частотами генотипических сочетаний ndtA1.1, ndtB1.1, ndtF2.2, ndtG1.1, ndtG1.2 и уровнем трофности выявлена существенная положительная
-
5. Вычисленные показатели генетических дистанций указывают на близость популяции Белая речка, Комсомольское озеро и Верхнее Голубое озеро, в то время как наиболее высокогорная популяция п. Эльбрус выпадает в отдельный кластер.
-
6. Имеющиеся данные пока не позволяют выявить степень связи между факторами среды и показателями кариотипа внутри отдельно взятых популяций, для чего необходимы повторные измерения параметров среды в течение более длительного времени.
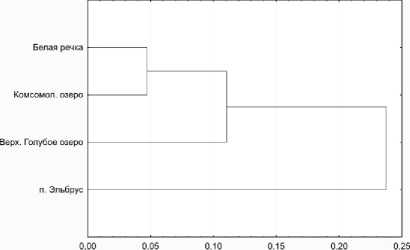
Рис. 1. Дендрограмма генетических дистанций между изученными популяциями, построенная по методу ближайшего соседа в евклидовом пространстве
корреляция, которая позволяет предварительно говорить о возможности использования частот данных сочетаний как индикаторов трофности того или иного водоема.
В дальнейшем планируется продолжение начатых исследований степени связи факторов среды и структуры кариотипа комаров-звонцов уже с использованием автоматических регистраторов температуры для получения наиболее полной картины динамики температурного режима в стациях исследования.
Работа выполнена в рамках госзадания «Оценка биоин-дикационного потенциала ресурсно-значимых беспозвоночных для практики мониторинговых исследований лесных экосистем Северного Кавказа».
Список литературы Предварительные результаты изучения взаимосвязи структуры кариотипа комаров-звонцов рода Chironomus (Diptera, Chironomidae) с факторами среды в условиях Центрального Кавказа
- Константинов, А.С. Биология хирономид и их разведение//Труды Саратовского отделения ВНИОРХ. -Саратов: ВНИОРХ, 1958. Т. 5. 358 с.
- Полуконова, Н.В. Эколого-кариологическая оценка последствий действия экологических факторов на хирономид (Chironomidae, Diptera)/Н.В. Полуконова, И.А. Федорова//Поволжский экологический журнал. 2006. № 2/3. С. 164-175.
- Ильинская, Н.Б. Зависимость уровня инверсионного полиморфизма от типа водоема, сезона и года наблюдений у мотыля Chironomus plumosus L. (Diptera, Chironomidae)/Н.Б. Ильинская, Н.А. Петрова, И. Матена//Общая генетика. 1999. Т. 35, №8. С. 1061-1070.
- Krieger-Wolff, E. Chironomiden aus der Umgebung von Freiburg i. Br. (mit Besonderer Berucksichtingung der Gattung Chironomus)/E. Krieger-Wolff, W. Wuelker//Beitr. Naturk. Forsch. Sudw-Dischl. 1971. Bd. 30. S. 133-145.
- Ильинская, Н.Б. Сезонная динамика хромосомного полиморфизма у мотыля Chironomus plumosus L. (Diptera, Chironomidae)/Н.Б. Ильинская, Н.А. Петрова, С.Ю. Демин//Генетика. 1988. Т. 24, №8. С. 1393-1401.
- Шобанов, Н.А. Кариофонд Chironomus plumosus (L.) (Diptera, Chironomidae). IV: Внутри-и межпопуляционный полиморфизм//Цитология. 1994. Т. 36, №11. С. 1129-1145.
- Michailova, P. Structural changes in the polytene chromosomes of Chironomus valkanovi Michailova, Cricotopus silvestris Staeg. and Cricotopus varius Fabr. (Dipreta, Chironomidae) under the influence of various factors//Comp. Rend. Acad. Bulg. Sci. 1979. Vol. 32. P. 377-380.
- Matena, I. Chromosomal polymorphism in a Chironomus plumosus (L.) (Diptera, Chironomidae) population from pond in Southern Bohemia: a long term studi//Heredity. 1991. Vol. 115, №2. P. 145-152.
- Балушкина, Е.В. Функционирование популяций хирономид в гипергалинных озерах Крыма/Е.В. Балушкина, Н.А. Петрова//Труды Зоол. ин-та АН СССР. 1989. Т. 205. С. 129-138.
- Michailova P. Untersuchungen uber den Chromo-somenpolymorphisms bei Chironomus salinarius Kieff., Chironomus valkanovi Michailova und Chirono-mus anchiatus Michailova (Diptera, Chironomidae) von der bulgarischen Schwartzmeerkuste//Zool. Anz. Leipzig. 1973. Bd. 191. S. 348-364.
- Белянина, С.И. Влияние ботулинического токсина на генетическую активность гигантских хромосом/С.И. Белянина, Н.П. Чеснокова, А.О. Колбенев, Т.А. Невважай//Бактериально-вирусные инфекции. -Саратов: Изд-во Сарат. гос. мед. ун-та, 1993. Ч. 1. С. 90-94.
- Полуконова, Н.В. Клеточные механизмы адаптации хирономид в условиях длительного воздействия низких температур/Н.В. Полуконова, А.В. Сокова, М.С. Козлов//Окружающая среда и гигиена: Мат-лы науч.-практ. конф. -Саратов: Изд-во Сарат. гос. мед. ун-та, 2002 а. С. 78-80.
- Полуконова, Н.В. Эколого-кариологический подход к исследованию популяций хирономид севера/Н.В. Полуконова, А.В. Сокова, М.С. Козлов//Проблемы экологии и биоразнообразия: Тез. докл. Междунар. конф. молодых ученых. -Борок, Институт биологии внутренних вод РАН, 2002 б. С. 69.
- Белянина, С.И. Реакция генома эукариот на воздействие факторов среды на примере хирономин (Diptera: Chironomidae)/С.И. Белянина, Н.В. Полуконова, Ю.В. Белоногова и др.//Современные проблемы популяционной экологии: Мат-лы Междунар. науч.-практ. экол. конф. -Белгород: Изд-во Белгород. гос. ун-та, 2006. С. 21-22.
- Ильинская, Н.Б. Изменчивость политенных хромосом слюнных желез личинок Chironomus plumosus в разные месяцы года/Н.Б. Ильинская, Ф.Л. Максимова//Цитология. 1976. Т. 18, №6. С. 747-751.
- Ильинская, Н.Б. Изменение рисунка и числа дисков в политенных хромосомах Chironomus plumosus в разные сезоны года/Н.Б. Ильинская, Ф.Л. Максимова//Цитология. 1978. Т. 20, №3. С. 291-296.
- Демин, С.Ю. Изменение компактности политенных хромосом из разных органов личинок мотыля Chironomus plumosus/С.Ю. Демин, Н.Б. Ильинская//Цитология. 1988. Т. 30, №4. С. 407-415.
- Белянина, С.И. Кариотипический анализ хирономид (Chironomidae, Diptera) фауны СССР: дис. … д-ра биол. наук. -М., 1983. 418 с.
- Филинкова, Т.Н. Хирономиды родов Chironomus Meigen и Camptochironomus Kieffer (Diptera, Chironomidae) Полярного Урала, Ямала и Приобской лесотундры (морфология и кариотипические особенности: Автореф. дис. … канд. биол. наук. -СПб., 1995. 16 с.
- Кикнадзе, И.И. Кариофонды хирономид криолитозоны Якутии: Триба Chironomini/И.И. Кикнадзе, А.Г. Истомина, Л.И. Гундерина и др.//Новосибирск: Наука. Сибир. отд-ние, 1996. 166 с.
- Кикнадзе, И.И. Кариотипы и хромосомный полиморфизм сибирских видов хирономид (Diptera, Chironomidae)/И.И. Кикнадзе, А.Г. Истомина//Сибирский экологический журнал. 2000. №4. С. 445-460.
- Wuelker, W. Karyosystematics and morphology of northern Chironomus (Diptera, Chironomidae): Freshwater species with larvae of the salinarius-type/W. Wuelker, M.G. Butler//Entomol. Scand. 1983. Vol. 14, №1. P. 121-136.
- Гундерина, Л.И. Внутривидовая дифференциация цитогенетической структуры природных популяций Chironomus plumosus L. -центрального вида группы видов-двойников (Chironomidae: Diptera)/Л.И. Гундерина, И.И. Кикнадзе, В.В. Голыгина//Генетика. 1999. Т. 35, № 2. С. 193-202.
- Кармоков, М.Х. Роды Chironomus Meigen, 1803 и Camptochironomus Kieffer, 1918 (Diptera, Chironomidae) Центрального Кавказа и Предкавказья: систематика, распространение и хромосомный полиморфизм: дис. … канд. биол. наук. -Саратов, 2013. 321 с.
- Соколов, В.Е. Млекопитающие Кавказа: Насекомоядные/В.Е. Соколов, А.К. Темботов. -М., 1989. 548 с.
- Демин, С.Ю. Кариотип комара Chironomus entis из группы plumosus в европейской части СССР/С.Ю. Демин, Н.А. Шобанов//Цитология. 1990. Т. 32. № 10. С. 1046-54.
- Nei, M. The genetic distance between populations//American Naturalist. 1972. № 106. P. 283-292.
- Keyl, H.-G. Chromosomenevolution bei Chironomus. II. Chromosomenumbauten und phylogenetische Beziehungen der Arten//Chromosoma. 1961. Bd. 13. S.464-514.
- Devai, Gy. Standardization of chromosome arms B, C, and D in Chironomus (Diptera, Chironomidae)/Gy. Devai, M. Miskolczi, W. Wuеlker//Advances in Chironomidology: Acta. Biol. Debr. Oecol. Hung. 1989. Vol.2. Pt.1. P. 79-92.
