Применение бактерий со сверхэкспрессией гена CADA для биоконверсии лизина в кадаверин

Автор: Ахова А.В., Федоненко П.М., Шумков М.С., Ткаченко А.Г.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 1, 2024 года.
Бесплатный доступ
Сконструирован штамм Escherichia coli со сверхэкспрессией гена cadA, кодирующего лизиндекарбоксилазу, и проведена оценка его способности конвертировать лизин в кадаверин в зависимости от рН среды. Клетки, сверхэкспрессирующие лизиндекарбоксилазу, получены на основе штамма E. coli BL21DE3, трансформированного плазмидой pET19b, несущей ген cadA из E. coli MC4100. Бактерии культивировали в 5 мл бульона LB при 37ºС без перемешивания, сверхэкспрессию cadA запускали добавкой 1 мМ изопропил-β-D-тиогалактопиранозида (при ОП600=0.6), через 2 ч. клетки отмывали и переносили в среду с рН 7.4 или 4.0 с добавкой 5 г/л L-лизина гидрохлорида. Количество синтезированного кадаверина определяли методом ТСХ с предварительной дериватизацией дансилхлоридом. Максимальная скорость биоконверсии наблюдалась при культивировании клеток, сверхэкспрессирующих cadA в среде с рН 4.0 (2.8±0.7 ммоль кадаверина/г АСБ в час). Скорость биоконверсии в базовых условиях (рН 7.4, базовая экспрессия) была в 6.8 раз ниже. Конечная концентрация кадаверина при культивировании в среде с рН 7.4 и/или в условиях отсутствия сверхэкспрессии гена cadA составила 0.8-1 мМ, в условиях сверхэкспрессии гена в нейтральной среде накапливалось до 1.2 мМ кадаверина, а в кислой среде - 1.8 мМ.
Кадаверин, лизин, декарбоксилаза, сверхэкспрессия, цельноклеточные биокатализаторы
Короткий адрес: https://sciup.org/147243409
IDR: 147243409 | УДК: 579.22 | DOI: 10.17072/1994-9952-2024-1-54-60
The use of bacteria with overexpression of the CADA gene for the bioconversion of lysine into cadaverine
The article is devoted to construction of an Escherichia coli strain with overexpression of the cadA gene encoding lysine decarboxylase, as well as to assessment of its ability to convert lysine to cadaverine depending on the pH of the medium. The overexpressing strain was constructed on the base of E. coli BL21DE3 strain transformed with the pET19b plasmid carrying the cadA gene from E. coli MC4100. Bacteria were grown in 5 mL of LB broth at 37 °C without stirring, overexpression of cadA was induced by the addition of 1 mM isopropyl-β-D-thiogalactopyranoside (at OD600 = 0.6), after 2 hours the cells were washed and transferred to a medium with pH 7.4 or 4.0 with the addition 5 g/L L-lysine hydrochloride. The amount of synthesized cadaverine was quantified by TLC with preliminary derivatization with dansyl chloride. The maximum rate of bioconversion was observed when cells overexpressing cadA were cultivated in a medium with pH 4.0 (2.8 ± 0.7 mmol cadaverine/g dry weight per hour). The rate of bioconversion under basal conditions (pH 7.4, basal expression) was 6.8 times lower. The final concentration of cadaverine when cultivated in a medium with pH 7.4 and/or in the absence of overexpression of the cadA gene was 0.8-1 mM; under conditions of overexpression of the gene, up to 1.2 mM of cadaverine accumulated in a neutral medium, and 1.8 mM in an acidic medium.
Текст научной статьи Применение бактерий со сверхэкспрессией гена CADA для биоконверсии лизина в кадаверин
Для клонирования и экспрессии рекомбинантных генов в клетках бактерий разработаны специальные генно-инженерные системы. В общем случае, рекомбинантный ген вводится в бактериальную клетку в составе векторной молекулы, которая – с целью повышения копийности – сохраняется в цитоплазме в состоянии эписомы. Генетически модифицированные подобным образом микроорганизмы обычно используют для наработки рекомбинантных белков, но также возможно их применение в качестве цельноклеточных биокатализаторов.
В частности, бактериальные клетки, способные к сверхэкспрессии лизиндекарбоксилазы за счет генетической модификации путем введения векторных молекул, пытаются использовать для биосинтеза кадаверина (1.5-диаминопентана). Особый интерес биосинтез кадаверина представляет потому, что на основе данного диамина могут быть получены пластики с улучшенными свойствами, а эффективный метод химического синтеза этого соединения не разработан [Kind et al., 2014; Huang et al., 2021]. В настоящее время существует два основных направления использования генетически модифицированных микроорганизмов для биосинтеза 1.5-диаминопентана: они либо выступают в качестве цельноклеточных биокатализаторов одностадийной реакции конверсии лизина в кадаверин, либо применяются для синтеза целевого вещества из простых субстратов в процессе ферментации [Ma et al., 2017; Nærdal, 2017].
Одним из первых был получен биокатализатор для конверсии лизина в кадаверин на основе клеток Escherichia coli , сверхэкспрессирующих ген cadA [Nishi et al., 2006], кодирующий индуцибельную изоформу лизиндекарбоксилазы в клетках E. coli , проявляющую максимальную активность при рН 5–6. В дальнейшем на основе клеток, сверхэкспрессирующих cadA , были разработаны биокатализаторы с дополнительными модификациями, направленными на повышение выхода продукта [Kim et al., 2015; Ma et al., 2015]. Также реализованы разработки на основе сверхэкспрессии гена ldcC , кодирующего конститутивную лизиндекарбоксилазу E. coli , максимально активную в среде с нейтральным рН [Oh et al., 2015; Shin et al., 2018].
Основным источником генов лизиндекарбоксилаз для создания биокатализаторов служит E. coli , хотя спектр используемых микроорганизмов постепенно расширяется. Например, созданы генно-модифицированные штаммы E. coli , сверхэкспрессирующие лизиндекарбоксилазы из клеток Klebsiella pneumonia , Ral-stonia eutropha , Pseudomonas aeruginosa и Hafnia alvei [Oh et al., 2015; Kim et al., 2016; Kim et al., 2019].
С целью создания биокатализаторов для синтеза кадаверина из лизина, как правило, применяют генно-инженерные конструкции, обеспечивающие сверхэкспрессию лизиндекарбоксилазы в ответ на добавку индуктора, в частности, сиcтемы на основе плазмид рET и рКЕ [Kim et al., 2015; Oh et al., 2015]. Альтернативным подходом является помещение гена, кодирующего лизиндекарбоксилазу, под контроль сильного конститутивного промотора, что обеспечивает его постоянную сверхэкспрессию [Qian, Xia, Lee, 2011; Ting et al., 2021].
Цель работы – конструирование штамма E. coli , сверхэкспрессирующего ген cadA , и оценка его способности конвертировать L-лизин в кадаверин в зависимости от рН среды.
Материалы и методы
Объекты и условия культивирования
В качестве объекта исследования использован штамм E. coli BL21DE3, трансформированный сконструированной нами плазмидой pET19b_cadA.
Бактериальные клетки, сохраняемые на скошенном агаре LB, переносили в 5 мл бульона LB (рН 7.4) с добавкой 1% глюкозы и культивировали при 37ºС без перемешивания в течение 16–18 ч. В среду для культивирования клеток, несущих плазмиду pET19b_cadA, добавляли 100 мкг/мл ампициллина. Полученную культуру разводили до плотности ОП 600 = 0.1–0.15 в 20 мл свежего бульона LB (рН 7.4) с добавкой (при необходимости) 100 мкг/мл ампициллина и культивировали при 37ºС без перемешивания. По достижении культурой плотности ОП 600 = 0.6 для индукции сверхэкспрессии гена cadA добавляли 1 мМ изопропил-β-D-тиогалактопиранозида (ИПТГ), после чего продолжали культивирование в описанных выше условиях в течение 2 ч. Затем бактериальную культуру центрифугировали (16 000 g, 3 мин.), и бактериальные клетки переносили в бульон LB с рН 7.4 или рН 4.0, содержащий 5 г/л L-лизина гидрохлорида (общий объем культуры составлял 5 мл). Культивирование проводили в описанных выше условиях.
Определение оптической плотности культуры и концентрации абсолютно сухой биомассы
Оптическую плотность измеряли при 600 нм (ОП 600 ) в кювете с длиной оптического пути равной 1 см с использованием спектрофотометра UV-1280 (Shimadzu, Япония). Концентрацию абсолютно сухой биомассы (АСБ) в культуре рассчитывали, используя предварительно полученный калибровочный график зависимости данного показателя и оптической плотности культуры (ОП 600 ).
Конструирование штамма E. coli со сверхэкспресссией гена cadA
Нуклеотидную последовательность гена cadA амплифицировали с применением праймеров cadANdeI 5’-GT CatATG TGTTGGGAGGGGCCTTTTTTAC-3’ и cadABamH 5’- GGATCC GGCAAGCCACTTCCCTTG-3’ с использованием в качестве матрицы хромосомы E. coli MC4100 (сайты рестрикции подчеркнуты; различия с нуклеотидной последовательностью E. coli W3110, GenBank AP009048.1 показаны строчными буквами). Полученный ПЦР-продукт длиной 2 226 пар нуклеотидов клонировали в вектор pGEM-T easy (Promega, США) согласно рекомендациям производителя. pGEM-T easy-вектор со вставкой секвенирова-ли с использованием стандартных праймеров М13F/R, плазмиду c подтверждённой последовательностью гена cadA выделяли из клеток методом щелочного лизиса [Maniatis, Fritsch, Sambrook, 1982] и обрабатывали эндонуклеазами рестрикции NdeI и BamHI (Fermentas, Литва). Полученный ген cadA очищали элек-трофоретически, экстрагировали из агарозного геля с применением набора DNA Extraction Kit (Fermentas, Литва) и лигировали в вектор pET19b (Novagen, США), предварительно также обработанный эндонуклеазами NdeI и BamHI. Полученную конструкцию pET19b_cadA проверяли с помощью ПЦР и секвенирования с использованием стандартных праймеров T7Pro/Term, после чего трансформировали в штамм E. coli BL21DE3 (Invitrogen) для экспрессии рекомбинантных белков.
Количественный анализ кадаверина осуществляли методом тонкослойной хроматографии с предварительной дериватизацией дансилхлоридом [Akhova, Tkachenko, 2022].
Статистический анализ результатов проводили с использованием пакета программ Statistica 6.0 (StatSoft Inc, США). Числовые данные в тексте представлены в виде среднее ± стандартное отклонение, на рисунке столбцы соответствуют среднему значению, отрезки (разбросы) – стандартной ошибке среднего.
Результаты и их обсуждение
В данном исследовании проведена оценка способности бактерий E. coli , характеризующихся сверхэкспрессией гена cadA , кодирующего лизиндекарбоксилазу, осуществлять биконверсию лизина в кадаверин. Для создания продуцента структурная часть гена cadA была слита с Т7 lac -промотором в составе плазмидного вектора pET19b (рис. 1). Полученную плазмиду pET19b_cadA трансформировали в штамм BL21DE3, представляющий собой производное E. coli BL21, в хромосому которого введен фаг λ DE3, несущий ген РНК-полимеразы Т7 под контролем laсUV промотора. Экспрессия последнего запускается добавкой изопропил-β-D-1-тиогалактопиранозида (ИПТГ), что приводит к наработке РНК-полимеразы фага Т7, обеспечивающей считывание целевого гена в составе эписомального вектора. Обязательными условиями сверхэкспрессии гена являлись присутствие плазмиды pET19b_cadA в клетках и добавка ИПТГ в среду культивирования. Источником гена cadA послужил штамм МС4100.
Затем было проведено сравнение способности продуцировать кадаверин клетками с базовым уровнем экспрессии лизиндекарбоксилазы, и клетками, сверхэкспрессирующими ген cadA , а также исследовано влияние рН среды на процесс биоконверсии (рис. 2).
Максимальной скоростью биоконверсии лизина в кадаверин характеризовались клетки, сверхэкс-прессирующие ген cadA, при культивировании на среде с рН 4.0. На второй час скорость биоконверсии в данных условиях составила 2.8±0.7 ммоль кадаверина/г АСБ в час, после чего наблюдалось снижение скорости процесса. В базовых условиях (рН среды 7.4, штамм без плазмиды) скорость биоконверсии равнялась 0.41±0.19 ммоль/г АСБ×ч, т.е. была почти в 7 раз меньше. Сверхэкспрессия гена cadA обеспе- чивала более высокую продукцию кадаверина и при культивировании в нейтральной среде, но скорость продукции была ниже по сравнению с клетками, выращиваемыми при рН 4.0.
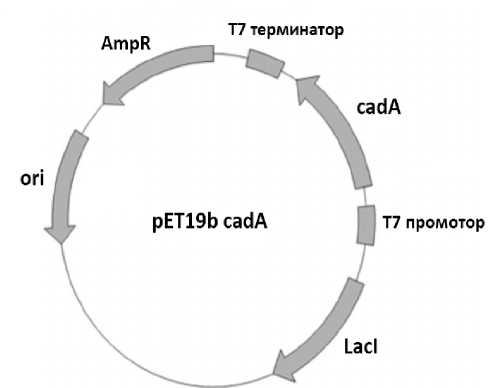
Рис. 1. Схема плазмиды pET19b_cadA.
AmpR – ген, кодирующий β-лактамазу (резистентность к ампициллину); LacI – ген, кодирующий репрессор lac -оперона; ori – район начала репликации вектора
[Diagram of the pET19b_cadA plasmid.
AmpR is a gene encoding β-lactamase (ampicillin resistance); LacI is a gene encoding a repressor of the lac operon; ori is the origin of replication]
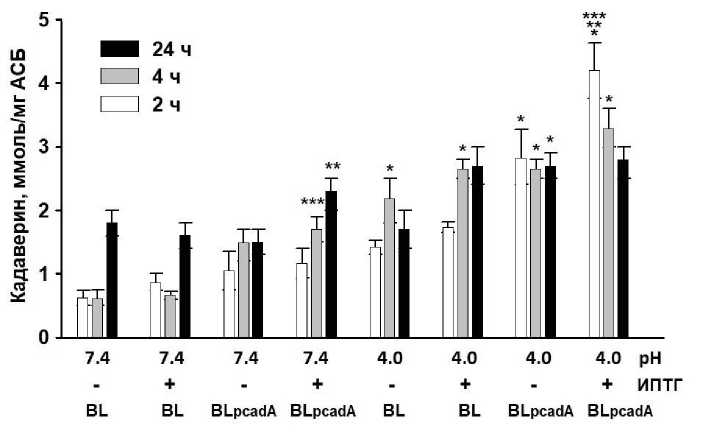
Рис. 2. Влияние рН среды и сверхэкспрессии гена cadA на продукцию кадаверина клетками E. coli BL21DE3.
«BL» – клетки, не несущие плазмиду pET19b_cadA, «BLpcadA» – клетки, несущие плазмиду pET19b_cadA. «+» – добавка 1 мМ ИПТГ (индуктора сверхэкспрессии), «–» – без добавки ИПТГ.
* – статистически значимое отличие от соответствующей культуры, выращиваемой на среде с рН 7.4;
** – статистически значимое отличие от соответствующей культуры без добавки ИПТГ;
*** – статистически значимое отличие от культуры штамма, не несущего плазмиду, при культивировании в аналогичных условиях (t-тест, р ≤0.05, n≥3)
[The effect of the pH of the medium and overexpression of the cadA gene on the production of cadaverine by E. coli BL21 DE3 cells.
“BL” – cells that do not carry the pET19b_cadA plasmid, “BLpcadA” – cells that carry the pET19b_cadA plasmid. “+” – the addition of 1 mM of IPTG (inducer of overexpression), “–” – without the addition of IPTG.
-
* – statistically significant difference from the corresponding culture grown on a medium with a pH of 7.4;
-
* * – statistically significant difference from the corresponding culture without the addition of IPTG;
-
* ** – statistically significant difference from the culture of a strain not carrying a plasmid when cultivated under similar conditions (t-test, p≤0.05, n≥3)]
Смещение рН среды в кислую сторону приводило к ускорению образования кадаверина как в условиях сверхэкспрессии, так и при базовой экспрессии гена cadA (в отсутствие плазмиды, без добавки ИПТГ). Увеличение лизиндекарбоксилазной активности в данном случае можно рассматривать как защитный ответ клетки на кислотный стресс, вызванный снижением рН среды [Meng, Bennett, 1992; Kanjee, Houry, 2013]. Кроме того, можно предположить, что повышение концентрации ионов водорода в среде сопровождалось смещением внутриклеточного рН в область, близкую к оптимальным значениям для лизиндекарбоксилазы CadA, что способствовало повышению скорости катализируемой ею реакции.
Можно также отметить тенденцию к увеличению продукции кадаверина клетками, несущими плазмиду сверхэкспрессии, при выращивании в среде с рН 4.0 без добавки индуктора ИПТГ, однако различия между трансформированными и не трансформированными плазмидой клетками не были статистически значимыми.
Установлено, что при всех вариантах условий культивирования конечная концентрация кадаверина в среде не превышала 2 мМ. При рН среды 7.4 и/или в условиях отсутствия сверхэкспрессии гена cadA итоговая концентрация кадаверина была в интервале 0.8–1 мМ. Сверхэкспрессия гена в нейтральной среде приводила к накоплению 1.2 мМ кадаверина, в кислой среде – к накоплению почти 1.8 мМ. Можно заключить, что данная концентрация является предельной для исследованного организма и лизиндекарбоксилазы CadA, выделенной из клеток E. coli MC4100. Прекращение процесса биоконверсии лизина в кадаверин, несмотря на достаточное количество субстрата в среде, может быть связано с ингибированием фермента по принципу обратной связи. Накопление кадаверина в среде до высоких концентраций оказывает в целом неблагоприятное воздействие на бактериальные клетки. Ранее показано, что добавка в среду культивирования 0.3–0.5 М гидрохлорида кадаверина вызывала частичный лизис клеток в культуре E. coli К12 [Qian, Xia, Lee, 2011]. Однако в процессе декарбоксилирования лизина с образованием кадаверина затрачивается ион водорода, а сам кадаверин, выделяющийся в среду, проявляет свойства основания. Это приводит к защелачиванию среды и смещению рН в область, неоптимальную для проявления активности фермента.
Таким образом, для повышения выхода целевого продукта, по-видимому, необходимо осуществлять культивирование штамма-продуцента в условиях поддержания оптимального значения рН и обеспечивать удаление конечного продукта реакции биоконверсии. Кроме того, решить данную проблему позволит поиск природных лизиндекарбоксилаз, толерантных к высокой концентрации конечного продукта, или получение таких ферментов методами белковой инженерии.
Заключение
Сконструирован штамм E. coli , сверхэкспрессирующий ген cadA , выделенный из E. coli MC4100. Скорость биоконверсии лизина в кадаверин в условиях сверхэкспрессии была в 6.8 раза выше, чем в базовых условиях. Скорость процесса биоконверсии и конечная концентрация кадаверина были выше при постановке реакции при рН 4.0 по сравнению со средой с нейтральным рН. Дальнейшая работа будет направлена на повышение продуктивности процесса биоконверсии с применением полученного продуцента, что может быть осуществлено за счет подбора оптимальных условий протекания реакции декарбоксилирования, в частности, варьирования состава среды культивирования, начальной плотности культуры продуцента, стабилизации продуцента за счет иммобилизации.
Список литературы Применение бактерий со сверхэкспрессией гена CADA для биоконверсии лизина в кадаверин
- Akhova A.V., Tkachenko A.G. Cadaverine biosynthesis in Escherichia coli adaptation to hydrogen peroxide // Applied Biochemistry and Microbiology. 2022. Vol. 58, № 5. Р. 582-589. DOI: 10.1134/S0003683822050039.
- Huang Y. et al. Green chemical and biological synthesis of cadaverine: recent development and challenges // RSC Advances. 2021. Vol. 11, № 39. P. 23922-23942. DOI: 10.1039/d1ra02764f.
- Kanjee U., Houry W.A. Mechanisms of acid resistance in Escherichia coli // Annual Review of Microbiology. 2013. Vol. 67. P. 65-81. DOI: 10.1146/annurev-micro-092412-155708.
- Kim H.J. et al. Optimization of direct lysine decarboxylase biotransformation for cadaverine production with whole-cell biocatalysts at high lysine concentration // Journal of Microbiology and Biotechnology. 2015. Vol. 25, № 7. P. 1108-1113. DOI: 10.4014/jmb.1412.12052.
- Kim H.T. et al. High-level conversion of l-lysine into cadaverine by Escherichia coli whole cell biocata-lyst expressing Hafnia alvei l-lysine decarboxylase // Polymers (Basel). 2019. Vol. 11, № 7. P. 1184. DOI: 10.4014/jmb.1602.02030.
- Kim J.H. et al. Functional study of lysine decarboxylases from Klebsiella pneumoniae in Escherichia coli and application of whole cell bioconversion for cadaverine production // Journal of Microbiology and Biotechnology. 2016. Vol. 26, № 9. P. 1586-1592. DOI: 10.3390/polym11071184. "
- Kind S. et al. From zero to hero - production of bio-based nylon from renewable resources using engineered Corynebacterium glutamicum // Metabolic Engineering. 2014. Vol. 25. P. 113-123. DOI: 10.1016/j.ymben.2014.05.007.
- Ma W. et al. Enhanced cadaverine production from L-lysine using recombinant Escherichia coli co-overexpressing CadA and CadB // Biotechnology Letters. 2015. Vol. 37, № 4. P. 799-806. DOI: 10.1007/s10529-014-1753-5.
- Ma W. et al. Advances in cadaverine bacterial production and its applications // Engineering. 2017. Vol. 3, № 3. P. 308-317. DOI: 10.1016/J.ENG.2017.03.012.
- Maniatis T., Fritsch E.F., Sambrook J.K. Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, 1982. 545 p.
- Meng S.Y., Bennett G.N. Nucleotide sequence of the Escherichia coli cad operon: a system for neutralization of low extracellular pH // Journal of Bacteriology. 1992. Vol. 174, № 8. P. 2659-2669. DOI: 10.1128/jb.174.8.2659-2669.1992.
- Nsrdal I. et al. L-lysine production by Bacillus methanolicus: genome-based mutational analysis and l-lysine secretion engineering // Journal of Biotechnology. 2017. Vol. 244, P. 25-33. DOI: 10.1016/j.jbiotec.2017.02.001.
- Nishi et al., 2006. US7189543B2 patent.
- Oh Y.H. et al. Development of engineered Escherichia coli whole-cell biocatalysts for high-level conversion of L-lysine into cadaverine // Journal of Microbiology and Biotechnology. 2015. Vol. 42, № 11. P. 14811491. DOI: 10.1007/s10295-015-1678-6.
- Qian Z.G., Xia X.X., Lee S.Y. Metabolic engineering of Escherichia coli for the production of cadaverine: a five carbon diamine // Biotechnology and Bioengineering. 2011. Vol. 108, № 1. P. 93-103. DOI: 10.1002/bit.22918.
- Shin J. et al. Characterization of a whole-cell biotransformation using a constitutive lysine decarboxylase from Escherichia coli for the high-level production of cadaverine from industrial grade l-lysine // Applied Biochemistry and Biotechnology. 2018. Vol. 185, № 4. P. 909-924. DOI: 10.1007/s12010-018-2696-4.
- Ting W.W. et al. Whole-cell biocatalyst for cadaverine production using stable, constitutive and high expression of lysine decarboxylase in recombinant Escherichia coli W3110 // Enzyme and Microbial Technology. 2021. Vol. 148. P. 109811. DOI: 10.1016/j.enzmictec.2021.109811.
