Применение методов тональной аудиометрии для оценки слуха серых тюленей в воздушной и водной среде

Автор: Пахомов М. В., Зайцев А. А., Литвинов Ю. В., Трошичев А. Р.
Журнал: Вестник Мурманского государственного технического университета @vestnik-mstu
Рубрика: Биологические науки
Статья в выпуске: 2 т.26, 2023 года.
Бесплатный доступ
Антропогенная шумовая нагрузка на традиционные места обитания морских млекопитающих продолжает возрастать в связи с активным освоением Арктической зоны. Изучение слуха арктических тюленей имеет большое значение для разработки мер по снижению степени воздействия техногенных шумов. В ходе исследования группу из десяти серых тюленей разного возраста и пола обучили нажимать на специальную клавишу при демонстрации звука чистого тона с частотой из стандартного и расширенного набора частот, рекомендованных для проведения тональной аудиометрии. Эксперименты проводили при размещении клавиши и источника звука в воздушной и водной (на глубине 2 м) среде. Основным показателем являлось время реакции тюленя на демонстрируемый стимул. Для каждого тюленя были составлены индивидуальные аудиограммы для слуха в воздухе и воде с использованием 32 частот. Анализ полученных данных показал, что время реакции может служить индивидуальным показателем при исследовании слуха серых тюленей, отражающим их возрастные особенности. По времени реакции на звуки с разной частотой тона можно установить биологически значимые и индифферентные частоты для серых тюленей.
Серый тюлень, аудиограмма, слух в воздухе, слух под водой, grey seal, audiogram, hearing in the air, hearing underwater
Короткий адрес: https://sciup.org/142237545
IDR: 142237545 | УДК: 500.7 | DOI: 10.21443/1560-9278-2023-26-2-121-130
Текст статьи Применение методов тональной аудиометрии для оценки слуха серых тюленей в воздушной и водной среде
e-mail: , ORCID:
e-mail: , ORCID:
Степень влияния антропогенного шумового загрязнения на морских млекопитающих является дискуссионным вопросом. Ряд авторов указывает, что морские млекопитающие покидают зоны с повышенным уровнем техногенного акустического шума; примером может служить снижение численности ластоногих в районе расположения ветроэлектростанций ( Wilson et al., 2010 ) или сокращение численности морских котиков в районе лежки вблизи от Пеликан Пойнт (Намибия) в результате увеличения морских и наземных перевозок ( Martin et al., 2022 ). Однако многие авторы публикуют результаты наблюдений за лежбищами ластоногих вблизи аэропортов или портов, где уровень шума достаточно высок ( Johnson et al., 1989; Bonner, 1983 ). Явные патологические изменения слуха у морских млекопитающих фиксируются при воздействии мощных источников звуковых сигналов, таких как сонары ( Harris et al., 2018 ) и подводные взрывы ( Wiggins et al., 2021 ), тем не менее в таких случаях сложно отличить травмирующее действие взрывной волны от воздействий именно звуковых колебаний.
Антропогенная нагрузка на традиционные места обитания морских млекопитающих, прежде всего арктических тюленей, продолжает возрастать в связи с активным освоением Арктической зоны. В указе Президента РФ от 26.10.2020 г. № 645 "О Стратегии развития Арктической зоны Российской Федерации и обеспечения национальной безопасности на период до 2035 г."1 предполагается интенсивное развитие судоходства и соответствующей портовой инфраструктуры региона на всем протяжении Северного морского пути. Согласно этому документу регион может пострадать из-за возрастающего антропогенного воздействия и сопутствующих климатических и экологических изменений, что, несомненно, увеличит шумовую нагрузку и на морских млекопитающих.
Таким образом, изучение слуха морских млекопитающих имеет большое значение для разработки мер по снижению степени воздействия техногенных шумов, оптимизации маршрутов и правил судоходства в арктических морях, правил по строительству и эксплуатации морской и прибрежной инфраструктуры в Арктическом регионе. Одним из наиболее информативных и наименее травмирующим методом исследования слуха является тоновая аудиометрия, так как, во-первых, в ходе ее применения собрана база данных медицинских и ветеринарных исследований, во-вторых, указанный метод отлично адаптируется с методами регистрации поведенческих реакций, используемых при работе с морскими млекопитающими. В научной литературе представлены исследования слуха серых тюленей в воздушной среде ( Ruser et al., 2014 ), ларг ( Sills et al., 2014 ) и кольчатых нерп ( Sills et al., 2015 ) в воздушной и водных среде. Основным показателем слуха в этих исследованиях служил пороговый уровень звукового давления, при котором животное выказывало поведенческую реакцию ( Sills et al., 2014; 2015 ) или регистрировались вызванные потенциалы спинного мозга ( Ruser et al., 2014 ). Следует отметить, что обе методики требуют либо долговременной высококвалифицированной тренерской подготовки исследуемого животного, либо применения наркоза, как при изучении слуха серых тюленей.
При применении тоновой аудиометрии основным показателем являлось время реакции на предъявляемый звуковой стимул, т. е. время между началом демонстрации звукового сигнала и нажатием на специальную клавишу. В предыдущих исследованиях зрения ( Пахомов и др., 2022 ) и слуха ( Литвинов и др., 2019; 2022 ) серых тюленей было установлено, что время реакции является достаточно надежным показателем сенсомоторной реакции исследуемого животного на стимулы разной модальности.
В ходе исследований ( Литвинов и др., 2022 ) тюлени были обучены реагировать нажатием на клавишу при демонстрации чистого тона на заданной частоте. Работы проводились в воздушной среде с использованием 10 типовых частот (125, 250, 500, 1 250 Гц; 3, 8, 12,5, 16, 20 кГц), наиболее часто применяемых при изучении слуха ластоногих ( Ruser et al., 2014; Sills et al., 2014; 2015 ). Экспериментально установлено, что время реакции на звуковой раздражитель может быть показателем сенсомоторной реакции серых тюленей. Полученные данные продемонстрировали индивидуальные, возрастные, половые различия испытуемых серых тюленей во времени реакции на звуки с разной частотой тона и послужили основой для исследования слуха серых тюленей не только в воздушной, но и в водной среде, с использованием как стандартного, так и расширенного набора частот, рекомендованных для проведения тональной аудиометрии (ГОСТ Р МЭК 60645-1-2017).
Целями настоящего исследования являются: 1) установление для каждого исследуемого тюленя показателей времени реакции на звуковой стимул чистого тона с частотами из наборов стандартного и расширенного диапазонов, используемых в тональной аудиометрии; 2) получение данных о восприятии набора звуковых стимулов в воздушной среде и аналогичных наборов звуковых стимулов в водной среде; 3) определение индивидуальных отличий во времени реакции на звуковой стимул в зависимости от частоты тона при восприятии в воздушной и водной среде; 4) сравнительный анализ индивидуальных показателей тюленей и выявление возрастных и половых отличий.
Материалы и методы
Экспериментальные работы проведены в аквакомплексе Мурманского морского биологического института РАН (ММБИ РАН) в Кольском заливе Баренцева моря.
В качестве экспериментальных животных использовались серые тюлени ( Halecoerus grupus Fabricius, 1791), которые подразделялись на три возрастных группы: 15-летние – 4 самки (тюлени под номерами 1–4); 5-летние – 3 самки (номера 5–7) и 1 самец (номер 8); 6-месячные самцы (номера 9 и 10). Все животные принадлежали к одной популяции; 15- и 5-летние тюлени в разное время были отловлены в районе Айновых островов в период окончания постнатальной линьки; 6-месячные тюлени были рождены самкой под номером 4 (тюлень 9 – в 2019 г., тюлень 10 – в 2021 г.). Эксперименты со взрослыми тюленями и молодым тюленем 9 проводились с 2020 г., с молодым тюленем 10 – с 2022 г.
Работы выполняли по парадигме "стимул – реакция", где стимулом служил звук чистого тона на различных частотах, а реакцией – нажатие тюленем специальной клавиши. Верная реакция подкреплялась пищевым поощрением в виде кусочка рыбы, вес одной порции рассчитывался индивидуально для каждого тюленя.
На момент начала экспериментальных работ все тюлени были физически здоровы, девиаций в поведении не имели; с ними проводились тренировки по выработке и закреплению навыка "стартовая позиция", когда тюлень должен находиться в определенной позиции и не покидать ее до подачи тренером команды (сигнала) на какое-либо действие. В ходе подготовительных тренировок команда тренера заменялась на звуковой сигнал экспериментальной установки, по которому тюлень должен был нажать на клавишу. В надводном положении тюлень располагался вертикально, по шею погружался в воду и удерживал морду на расстоянии не более 1 см от клавиши (или вплотную к ней), ожидая подачи звукового сигнала. Данная позиция была отработана с помощью команды "Таргет!" и поощрения требуемой позиции. В подводном положении тюлень находился в горизонтальном положении на глубине 2 м и также удерживал морду на расстоянии не более 1 см от клавиши (или вплотную к ней), ожидая подачи звукового сигнала. Данная позиция формировалась на базе уже отработанной позиции в надводном положении, но теперь подкреплялось вертикальное "зависание" тюленя на уровне клавиши. Выработка этой позиции упрощалась тем, что подобное зависание является характерным паттерном поведения серого тюленя, которое в естественных условиях используется для засадной охоты ( Bouveroux et al., 2014 ). Также у всех исследуемых животных был сформирован навык "концентрации на выполняемой задаче", т. е. подаваемые тренером и экспериментальной установкой команды выполнялись четко и максимально быстро.
У 15- и 5-летних тюленей большинство навыков были отработаны в ходе предыдущих научных и научно-практических работ; 6-месячных тюленей в разное время обучали необходимым навыкам (начиная с 3-месячного возраста).
Для проведения исследования была разработана экспериментальная установка (рис. 1). Тюленям демонстрировались 32 чистых тона из стандартного (125, 160, 200, 250, 315, 400, 500, 630, 750, 800, 1 000, 1 250, 1 500, 1 600, 2 000, 2 500, 3 000, 3 150, 3 200, 4 000, 5 000, 6 000, 6 300, 8 000 Гц) и из расширенного набора частот (9 000, 10 000, 11 200, 12 500, 14 000, 16 000, 18 000, 20 000 Гц), рекомендованных для проведения тональной пороговой аудиометрии (ГОСТ Р МЭК 60645-1-2017)2. Звуковой сигнал создавался в генераторе частоты на основе микросхемы AD9833 по команде с микроконтроллера Arduino UNO R3 и проходил предусиление на цифровом широкодиапазонном усилителе на базе чипа TDA2050, а затем усиливался на усилителе Ural BV 2.70. Звуки на частотах до (и включая) 1 000 Гц воспроизводились на подводной колонке Aquasonic AQ339 (2а); звуки на частотах свыше 1 000 Гц воспроизводились с помощью акустического излучателя на основе пъезоэлектрической диафрагмы CBC2065BAL (2b). Блок динамиков (2) и клавишу (3) размещали на дистанции 1 м друг от друга в воздухе на высоте 30 см от уровня воды и под водой на глубине 2 м.
На каждой тренировке с тюленем проводились работы как в воздушной среде, так и в водной; при этом очередность размещения оборудования в воздушной и водной среде изменялась. Если на первой тренировке сначала оборудование размещалось над водой, а затем погружалось на глубину 2 м, то на следующей сначала проводились подводные испытания, а по их завершении – надводные, и наоборот. Для тюленей 1–9 вопрос первого размещения оборудования решался с помощью порядкового номера исследуемого животного: четные номера – первое размещение оборудования в воздушной среде, нечетные номера – в водной. Для молодого тюленя 10 было выбрано первое размещение оборудования в воздушной среде.
Частоты были разделены на три блока: низкие (125–800 Гц); средние (1 000–8 000 Гц); высокие (9 000–20 000 Гц). Сначала с тюленем отрабатывался блок средних частот, затем низких, затем высоких. В каждом блоке каждая частота звучала дважды. Последовательность частот генерировалась случайным образом, при этом одна и та же частота не воспроизводилась дважды подряд. Для генерации случайных чисел использовался интегрированный в Arduino UNO R3 генератор псевдослучайных чисел с источником в виде электромагнитных помех на первом аналоговом входе.

Рис. 1. Схема экспериментальной установки: А – в надводном положении; Б – в подводном положении; 1 – блок питания, генерации и усиления звукового сигнала; 2 – блок динамиков; 2а – низкочастотный динамик; 2b – высокочастотный динамик; 3 – клавиша для взаимодействия с тюленем;
4 – блок управления; 5 – пульт управления; 6 – блок контейнеров с рыбой
Fig. 1. Experimental installation scheme: A – in air, B – in water; 1 – power, generation and amplifying unit;
2 – acoustic speaker’s complex; 2a – low frequency speaker; 2b – high-frequency speaker; 3 – the key for interacting with the seal; 4 – control unit; 5 – control plate; 6 – a case of containers with fish
Интенсивность звука была константна и составляла 50 Дб (отн. 20 мкПа) на расстоянии 1 м от источника на всем диапазоне используемых частот; уровень громкости подстраивался системой автоматически для каждой частоты в соответствии с калибровочными данными для воздушной и водной среды. Такой уровень звукового давления был выбран потому, что, во-первых, естественный уровень акустического шума в районе проведения эксперимента составлял в среднем 35 дБ (отн. 20 мкПа) над водой и 38 дБ (отн. 20 мкПа) под водой на глубине 2 м; во-вторых, при таком уровне звукового давления звук воспринимается именно слуховой системой и не имеет тактильного воздействия, что показано в ряде исследований ( Levanen et al., 2001; Yau et al., 2010; Bernard et al., 2022 ).
Работы проводились два раза в день утром и вечером, когда уровень естественного акустического шума минимален и не превышает 40 Дб (отн. 20 мкПа) над и под водой.
Экспериментальная установка работала в автоматическом режиме по следующей схеме. Оператор перед началом каждого опыта оценивал состояние тюленя и окружающей среды, если все было в норме, то принимал решение о начале работы. В контейнеры (5) заряжались 10 порций рыбы; на смартфоне в блоке управления (4) выбирался тюлень, с которым будет проводиться исследование, и по нажатию кнопки на пульте управления (5) запускалась программа опыта. После запуска оператор отходил от вольера, чтобы тюлень не мог его видеть. Система отсчитывала 30 с, давая человеку возможность удалиться, и запускала первый цикл опытов. Установка в случайном порядке выдерживала интервал времени в диапазоне 10–30 с и подавала звуковой сигнал заданной частоты в течение 3 с; если тюлень в это время нажимал на клавишу (3), то система подавала звуковой бридж-сигнал положительного подкрепления (долгий свист), затем запускала механизм открывания первого контейнера (в дальнейших циклах номер контейнера соответствовал номеру цикла), выдавая тюленю кусочек рыбы. Далее система делала задержку в работе на 10 с, давая тюленю возможность съесть кусочек рыбы, а при подводной работе вдохнуть воздуха, и начинала новый цикл.
Если тюлень нажимал на клавишу, когда звуковой стимул не транслировался, то система подавала звуковой бридж-сигнал наказания (два коротких свиста) и начисляла к времени ожидания "штрафные" 10 с. Пропуск звукового сигнала не наказывался; система выдерживала интервал времени в диапазоне 10–20 с и снова проигрывала тот же звуковой сигнал.
Проведя 10 циклов, система делала перерыв в опыте, переходила в режим ожидания и посылала оператору голосовое послание, дублируя его миганием индикаторной лампы на пульте управления (5). Оператор возвращался к вольеру, оценивал состояние тюленя и принимал решение о продолжении или окончании опытов.
Система самостоятельно прекращала опыт в двух случаях: 1) если тюлень начинал бессистемно нажимать или удерживать нажатой клавишу, то каждые 2 с ему начислялись "штрафные" 10 с; при наборе суммарного времени ожидания более 120 с система прекращала опыт и посылала оператору соответствующее звуковое сообщение; 2) если тюлень более 5 раз подряд игнорировал подачу звукового сигнала, то система также прекращала текущий опыт и сигнализировала оператору.
Результаты и обсуждение
С каждым исследуемым тюленем для каждой демонстрируемой частоты проводили по 100 опытов в воздушной и водной среде. Результаты в виде средних значений времени реакции на звуковой стимул представлены на рис. 2.
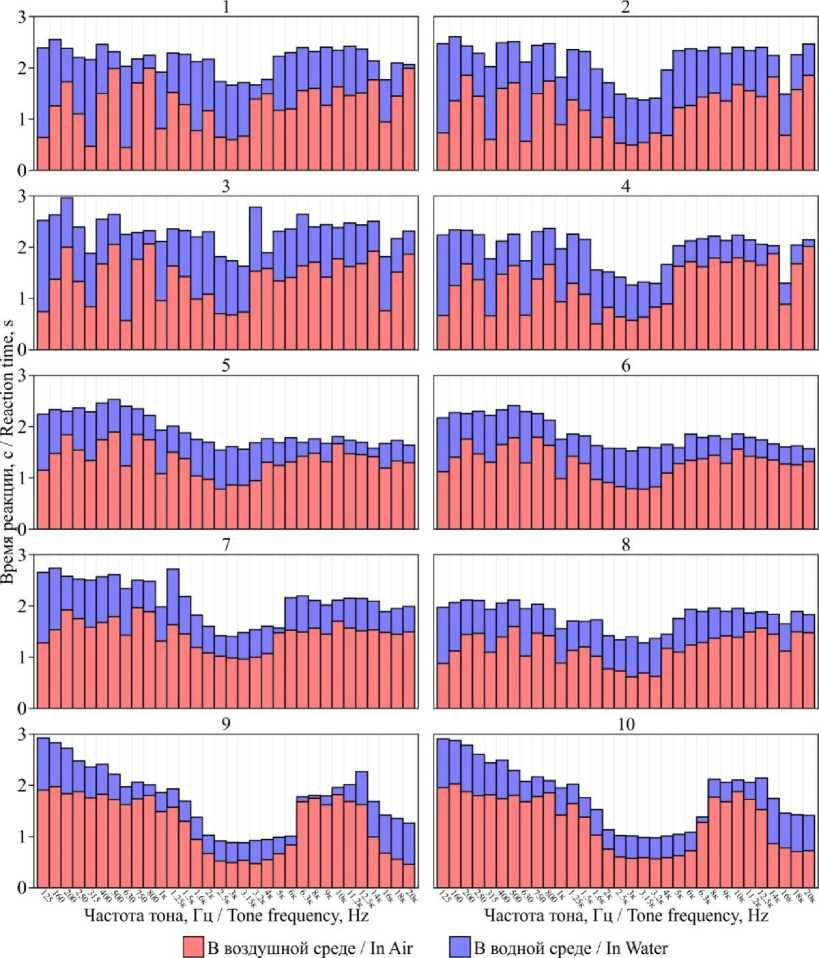
Рис. 2. Индивидуальные аудиограммы серых тюленей Fig. 2. Individual audiograms of grey seals
Индивидуальные аудиограммы исследованных серых тюленей показали, что животные подразделяются на пять групп.
В первую группу входили 15-летние самки, при этом для выборок на частоте 4 кГц они подразделялись на две подгруппы: 1) тюлени 1 и 3; 2) тюлени 2 и 4.
Во вторую группу входили 5-летние самки; их внутригрупповые отличия на всем диапазоне частот были минимальны; от группы 15-летних самок они отличались большим временем реакции и более плавным изменением времени реакции при использовании соседних частот.
Третью "группу" представлял 5-летний самец, который отличался от тюленей второй группы более коротким временем реакции, от тюленей первой группы - более плавным изменением времени реакции при использовании соседних частот (характерным для тюленей его возраста).
Молодые 6-месячные тюлени образовывали четвертую группу, наибольшим отличием от остальных групп являлась большая величина стандартного отклонения на всех частотах [кроме частот 2-4 кГц и 14 кГц и выше, где величина стандартного отклонения была соизмерима с другими группами (диапазон 2-4 кГц) или была меньше (диапазон от 14 кГц и выше), в этих же диапазонах время реакции было значительно ниже, чем у тюленей других групп].
Экспериментально установлено, что начиная с частоты 12,5 кГц у 15-летних самок время реакции возрастает, 5-летних - незначительно снижается, 6-месячных - резко уменьшается. Данные тренды различаются по причине снижения с возрастом верхнего предела воспринимаемых частот, что описано в ряде исследований ( Schusterman et al., 2002; Cunningham et al., 2016; D’Agnese et al., 2021 ).
Время реакции всех исследованных тюленей в воздухе меньше, чем время реакции под водой, но при этом разница значений в большинстве случаев константна и составляет около 1 с (у 15-летних тюленей), 0,5-0,7 с (у 5-летних, независимо от пола) и 0,2-0,5 с (у 6-месячных тюленей). Меньшую разницу между временем реакции в воздухе и под водой у молодых тюленей можно объяснить их меньшим весом и линейными размерами, что позволяет быстрее передвигаться под водой. Анализ видеопротоколов показал, что время между началом подачи звукового сигнала и моментом, когда тюлень начинает движение, в среднем одинаково для идентичных частот в воздушной и водной среде.
Следует отметить, что на определенных частотах время реакции в воздухе и под водой значительно различалось.
Так, 15-летние тюлени очень быстро (относительно соседних частот) реагировали на звуки с частотами 125, 315 и 630 Гц в воздухе и менее быстро - под водой (кроме тюленей 3 и 4, у которых время реакции на данных частотах под водой значительно отличались от соседних частот). У 5-летних и 6-месячных тюленей такой тенденции выявлено не было. Данную особенность можно объяснить разницей в размерах: у 15-летних тюленей ушной обхват черепа на момент начала эксперимента составлял в среднем 560 ± 15 мм, у 5-летних самок - 520 ± 11 мм, у 5-летнего самца - 540 мм, у 6-месячных - 450 и 470 мм. Эти различия, вероятно, приводили к разным размерам слухового канала, поэтому у взрослых тюленей данные частоты резонировали и, соответственно, усиливались.
В ходе экспериментов выявлена более быстрая (относительно соседних частот) реакция 15-летних тюленей и 5-летнего самца на звук с частотой тона 16 кГц как в воздухе, так и подводой. Этот факт можно объяснить реакцией на хлопки ластами, характерные для половозрелых самцов серого тюленя в период размножения, частотный пик которых приходится именно на 16 кГц ( Hocking et al., 2020 ). Данные звуки имеют биологическое значение для половозрелых 15-летних самок, связанное с поиском партнера для спаривания, а для 5-летнего самца, находящегося в начале полового созревания ( Boness et al., 1979; Beest et al., 2019 ), - с возможностью избежать встречи с крупными половозрелыми самцами, чем и обусловлено быстрое время реакции на звук чистого тона с частотой 16 кГц.
При использовании средних частот 1-4 кГц у всех тюленей в воздушной и водной среде отмечается самое быстрое время реакции. В этом диапазоне находятся гортанные рыки, издаваемые серыми тюленями как на суше, так и в воде, и щелчки, фиксируемые только под водой. Ряд авторов ( Asselin et al., 1993; Miksis-Olds et al., 2016; Nowak, 2021 ) предполагают, что именно эти звуки являются основными коммуникативными сигналами, имеющими амплитудно-темпоральную модуляцию на индивидуальной для каждой особи частоте (форманте), обычно находящейся в пределах 2-3,5 кГц для серых тюленей ( Stansbury et al., 2019 ).
Изменения на данных частотах у 5-летних тюленей выражены нечетко, в то время как у 6-месячных и 15-летних на данном диапазоне частот время реакции значительно отличается от более низких и более высоких частот. У 6-месячных тюленей наиболее быстрая реакция фиксируется при применении частот 2-6 кГц, 15-летние тюлени 1 и 3 демонстрируют наиболее быструю реакцию на частотах 3-3,2 кГц, тюлени 2 и 4 имеют более широкий диапазон - 1,6-4 кГц. При этом оба молодых тюленя были рождены и выкормлены 15-летней самкой 4, а самка 2 рожала щенка, но не выкармливала его (Zaytsev et al., 2021). Также у рожавших самок время реакции на частоты 3; 3,1 и 3,2 кГц меньше, чем у яловых самок 1 и 3. Данный факт можно объяснить физиологическими изменениями тюленей 2 и 4, связанными с рождением и выкармливанием щенков. Согласно ряду исследований, посвященных вокализации серых тюленей в период лактации (Caudron et al., 1998; Stansbury et al., 2021), коммуникация самки со щенком происходит на частотах 3–4 кГц.
Заключение
В результате исследования получены индивидуальные аудиограммы серых тюленей в воздушной и водной среде. Аудиограммы, основанные на времени реакции, имеют зависимость от возрастов испытуемых тюленей: для молодых тюленей характерен более ровный график, время реакции на соседние частоты возрастает и убывает более плавно, чем у взрослых тюленей, на аудиограммах которых наблюдаются скачкообразные переходы. Под водой относительная разница во времени реакции на соседние частоты ниже, чем в воздушной среде на аналогичные частоты. При использовании частот среднего спектра, которые серые тюлени используют для коммуникации, время реакции значительно меньше, чем остальных частот. В ходе экспериментов выявлены частоты (125, 315 и 630 Гц), время реакции на которые при восприятии в воздушной среде значительно меньше, чем на соседние частоты, что особенно четко проявляется у 15-летних тюленей, слабо выражено у 5-летних тюленей и не наблюдается у 6-месячных тюленей. При восприятии данных частот в воде резкие отличия во времени реакции наблюдаются только у рожавших 15-летних самок. В верхней части спектра предъявляемых частот внимание привлекает 16 кГц, быстрая реакция на которую четко выражена у всех 15-летних тюленей как в воздушной среде, так и в водной; снижение скорости реакции выражено у 5-летнего самца; быстрая реакция не наблюдается ни у 5-летних самок, ни у молодых тюленей.
Полученные данные и опыт могут служить основой для проведения дальнейших исследований с использованием электрографических методов фиксации времени реакции, исключающих влияние различий во времени, затрачиваемом тюленями на совершение движения нажатия на клавишу: электромиографии или метода вызванных потенциалов спинного мозга ( Ruser et al., 2014 ).
Конфликт интересов
