Принцип целесообразности и моделирование биосистем

Автор: Коросов Андрей Викторович
Журнал: Ученые записки Петрозаводского государственного университета @uchzap-petrsu
Рубрика: Биология
Статья в выпуске: 7 (101), 2009 года.
Бесплатный доступ
Системный подход, моделирование, целесообразность
Короткий адрес: https://sciup.org/14749599
IDR: 14749599
Текст статьи Принцип целесообразности и моделирование биосистем
Цель научного биологического исследования состоит в поиске закономерностей, для которых наиболее конкретной и ясной формой выражения выступает количественная модель. Модельные коэффициенты пропорциональности (параметры) могут получить биологическую интерпретацию как ключевые характеристики функционирования биосистемы. К сожалению, практика моделирования все еще слабо развита в среде эколого-биологических исследований. Причина, на наш взгляд, состоит в недоверии биологов к возможностям математики показать нечто большее, чем видно непосредственно по результатам простейших количественных обобщений. Иными словами, причина – в непонимании данного инструмента исследования. В этой связи мы видим смысл в популяризации системного подхода, последовательное проведение которого не только приводит к конструированию количественной модели изучаемого явления, но и к существенно более стройному структурированию имеющейся информации, к более глубокому пониманию самой проблемы, возможности выражать биологическое знание в точных числовых утверждениях [13].
Системный подход (как общенаучную методологию) можно представить в виде семи принципов мышления (системности, целесообразности, организации, иерархии, эмерджентности, целостности, развития), первый из которых был детально рассмотрен нами ранее [14]; на очереди – второй. Смысл настоящего сообщения состоит в том, чтобы продемонстрировать разнообразие целевых установок на разных этапах модельного исследования биосистем.
Принцип целесообразности можно сформулировать так:
Границы, содержание и поведение системы определяются целью.
Система – это тот специально ограниченный набор взаимосвязанных объектов (элементов), с помощью которого достигается определенная цель.
Эти определения имеют двоякий смысл. С одной стороны, они относятся к природным объектам (биосистемам), существование кото-
рых поддерживается благодаря их «целесообразной» конструкции и функционированию. Система – это то, что выполняет функцию. Внешне эта функция выглядит как цель, то, «для чего» существует система. Системная функция предписывает системе определенный состав (структуру) и динамику (организацию). Система состоит только из тех элементов, которые участвуют в выполняемой функции. Однако эти онтологические определения не только не точны, но и прямо мешают выполнять процедуры моделирования [15].
Принцип целесообразности вскрывает технологию процесса познания объекта природы, обеспечивая эффективное (в том числе и практичное) системное его описание. « Система есть средство достижения цели » [25; 68]. Этот тезис логически вытекает из принципа системности: коль скоро система есть алгоритм мысли об объектах природы [14], то композиция системы в первую очередь должна определяться причиной, вызывающей эту мысль, то есть целью размышления о биосистеме. Таким образом, на структуру представления объекта природы (на его фрейм и модель) накладывает отпечаток не только строение и динамика самого познаваемого объекта, но и средства его описания, обусловленные имеющейся информацией и доминирующей теорией (парадигмой). В состав моделируемой системы включают только те элементы, которые, на наш взгляд , и осуществляют системную функцию. Системное описание биологического объекта (в форме фрейма или модели) всегда неполно, тенденциозно, но целесообразно.
Итак, чтобы изучить целесообразное поведение биосистем, следует построить их целесообразные модели – но в первую очередь необходимо явно отделить видные нам «цели» природных объектов от «целей» исследователя. (В скобках следует заметить, что термин «целесообразность» в бытовом понимании ближе к понятию «практичность». В нашем изложении термин более соответствует понятию «целенаправленность», когда структура и поведение объекта «сообразуются» с некоторой предзаданной целью.)
ПРИРОДНАЯ ДИНАМИКА
В отношении объектов природы понятие цели используется в том случае, если это бихевиоральные (и социальные) системы, имеющие центр организации поведения – мозг [27]. Для биосистем, не имеющих специфических центров управления, применим термин объективной цели. «У естественных неодушевленных систем есть только объективные цели… Будущее реальное состояние системы мы представляем себе как ее объективную цель» [25; 101]. «У компенсации есть “цель”. Ею является конечное нормальное функционирование, то есть получение определенного приспособительного результата» [1; 24]. Факт существования биосистемы означа- ет, что она достигает свои цели, то есть приспособлена к среде; факт элиминации означает, что цель не достигнута. Критерием достижения цели служит «приспособительный результат», способность пройти естественный отбор.
В более широком плане можно говорить о следующих четырех формах природных процессов. 1. Порождение разнообразных условий и факторов среды, с которыми сталкиваются особи (осуществляется в пределах биогеоценоза и биосферы). 2. Развитие, онтогенез особей (формирование сомы и функций). 3. Функционирование особей популяции (включая процессы размножения). 4. Естественный отбор особей (выживание потомков особей в данной среде).
В биогеоценозе формирование абиотических факторов определяется физико-химическими и геолого-географическими процессами, не требующими для своего описания понятия целесообразности. «Физика… не знает ничего о цели» [6; 232]. В функционировании биотической части биогеоценоза можно усмотреть объективную цель в виде «стремления» всех видовых популяций ценоза к неограниченному росту и возникающей на этой базе конкуренции, ведущей к полноценному использованию ресурсов среды. Так формируется та или иная динамическая структура доминирования в ценозе, в целом независимая от конкретного видового состава – идентичное строение могут иметь сообщества, состоящие из разных видов [31]. С одной стороны, формирование ценоза из видов, обладающих разными «врожденными» способностями, есть чисто стохастический (комбинаторный, «физический») процесс, который не должен поэтому ассоциироваться с «целью» ценоза. С другой стороны, само возникновение видов, утилизирующих свободные ресурсы (осваивающих незанятые ниши), есть эволюционный адаптивный и поэтому – объективно целесообразный процесс. (Более детальное рассмотрение адаптивных черт биоценозов и популяций выходит за рамки статьи.) Для описания подобной «целесообразной» стабилизации экологических характеристик сообщества или популяции часто эффективно используется телеономическая терминология вроде «для того чтобы выжить, популяция должна…», а в математические формулы вводятся такие телеономические конструкции, как максимальная плотность популяции, К (уравнение Лоттки-Вольтерра [8]), эффективная численность популяции [29], доступный объем энергии [3] и др. По большому счету, любая биологическая константа представляет собой реализованную цель – конечную характеристику целесообразной биосистемы.
Онтогенез есть по большей части реализация наследственной информации, заложенной в геноме и цитоплазме зиготы, модифицированной условиями среды. Его объективная цель «материально» выражена набором наследственных факторов и является в форме адаптированного организма, целесообразно реагирующего на внешние факторы на любой стадии своего развития. «Цель» онтогенеза гораздо более отчетливо задана, чем «цели» ценоза или популяции. В экологических исследованиях описание онтогенеза обычно ограничивается характеристикой аллометрических законов роста тела особей [22] и их частей [33] в форме уравнений регрессии. При этом также иногда бывают полезными такие телеономические конструкции, как «максимальный размер особи», W∞ [18].
Функционирование особи отчетливо целесообразно, если обеспечивает ее существование. Эта цель предписывается самим строением биосистемы. Птица летит, рыба плывет, сердце перекачивает кровь – цель функционирования состоит в автономной реализации возможностей данной конструкции биосистемы (органа, организма, популяции). Функционирование есть постоянный процесс реагирования организма на изменение внешних и внутренних факторов (толерантность и гомеостаз). Реактивность есть внутреннее качество изучаемой биосистемы, прямое следствие ее устройства. Таким образом, цель организма – осуществлять потенции своей организации. «…Внутренняя цель может быть инструктирована просто как программа поведения, обеспечивающая стабильность системы относительно параметров выживания» [27; 101]. «Цель системы – некоторая область в пространстве состояний, которую необходимо достичь в процессе функционирования» [32; 121]. «Биологическая система не может самостоятельно определить свое назначение или цель, но она может выбрать функции для достижения запрограммированной цели» [28; 77]. Такое понимание приближает нас к формулировке целей моделирования, имеющей яркое телеономическое звучание. Например, при формальном описании гомеостаза и поведения в качестве цели выступает некое «штатное» (нормальное) значение важной характеристики внутренней среды, на поддержание которой направлено функционирование биосистемы [14]. Примерами таких величин являются температура тела гомойотермых животных, концентрация углекислоты и глюкозы в крови и пр. Рассматривая любое явление более широко, нельзя не увидеть, что естественное разнообразие особей выполняет множественность реализаций видового типа гомеостаза. Это значит, в частности, что для описания «видовой цели» следует применять статистическую характеристику.
Процесс естественного отбора (в контексте изложенного выше) состоит в том, что далеко не все особи (варианты реализации организменной жизни) в конечном итоге адаптивны, то есть способны выжить и успешно размножиться. Множество особей не может оставить свой генетический след в будущих поколениях популяции. В этом контексте лишь часть особей достигает эволюционных целей, другая же – не дости- гает, элиминируется. Естественный отбор протекает в среде биогеоценоза, которая сама по себе не постоянна, изменчива. Это значит, что требования, предъявляемые к организмам, постоянно меняются, как и векторы отбора. Иными словами, для особей отсутствует постоянная однозначная эволюционная цель. Возможность достижения цели обеспечивается диапазоном адаптивного реагирования. Выход уровня внешних факторов за рамки этого толерантного диапазона ведет к элиминации особей, их цели функционирования не достигаются. Экологи очень редко обращаются к практике моделирования с учетом естественного отбора [21]. В большой степени это оправданно, поскольку эволюционные процессы собственно экология не изучает. В то же время без учета эффектов отбора описание естественных биосистем становится неверным. Например, обычная практика экологического исследования популяций состоит в том, чтобы изученные в природе характеристики всех особей популяции включать в общую выборку и вычислять общие статистические параметры, считая их нормой. Однако часть особей, отловленных из общей совокупности, не смогла бы пройти отбор (погибла бы до размножения). Следовательно, в действительности эта часть выборки не представляет норму функционирования вида – это не адаптивные, не целесообразные формы, которые поэтому не могут представлять филум (эволюционирующий вид). Исправить эту некорректность можно только явно разделяя целесообразных и нецелесообразных особей и изучая их по отдельности; примеры тому есть в литературе [34].
Представленные четыре аспекта видения объектов природы с точки зрения их целевых устремлений, естественно, не исчерпывают проблемы целесообразности жизни. Мы стремились лишь показать, что интерпретация любых природных явлений может включать понятие цели. В то же время данное описание живого предпринято с неявно заданных позиций синтетической теории эволюции. С точки зрения номогенеза, игнорирующего отбор как эволюционный фактор [4], те же феномены получили бы совсем другую интерпретацию [19]. Взяв на вооружение разные парадигмы, исследователи будут строить отличающиеся словесные описания. Создавая модель, то есть придавая количественную форму неким (в том числе представленным выше) биологическим высказываниям, следует помнить, что эта модель унаследует теоретический образ биосистемы. Следование распространенной на данный момент теории всегда выступает в роли одной из целей исследования.
МОДЕЛИРОВАНИЕ
Одним из простых и эффективных способов детального количественного описания динамики биосистем является построение имитационных моделей в среде Excel [12], [23]. Генеральная цель такого моделирования состоит в вычислении значений коэффициентов пропорциональности, выражающих важные соотношения между компонентами моделируемой биосистемы и имеющих отчетливые биологические интерпретации. Структура процесса моделирования достаточно сложна, причем не столько в техническом отношении (Excel максимально упрощает задачу), сколько в идеологическом – нужно отчетливо представлять себе цели всех этапов работы. Наша задача состоит не в детальном описании алгоритма построения моделей (см. подробнее [13]), но в выявлении скрытых целей этого процесса. Их подразделение мы построили по схеме, принятой выше для природных эволюционирующих объектов.
-
1. Формирование массива исходных данных.
-
2. Подбор вариантов конструкций и построение (перестройка) модели. 3. Реализация (расчеты по) модели. 4. Настройка параметров модели (методом подбора).
Массив исходных данных должен хорошо соответствовать изучаемому процессу, в явной форме (или через градации) отображать состояния биосистемы, стадии ее развития, внутреннюю дифференциацию на компоненты (виды, популяции, группы, особи) и природное окружение (факторы среды). Здесь рассматривается только моделирование, то есть предполагается, что тема, содержательные задачи научного исследования, выбор парадигмы для ее решения, общее описание процесса выполнены ранее. Понятно, что в формулировании научного вопроса заложен механизм целеполагания, определено направление, в котором должна двигаться исследовательская мысль. «...Умный вопрос – это уже добрая половина знания. Ведь Платон правильно говорит: “Тот, кто о чем-то спрашивает, уже представляет себе в самом общем виде то, о чем он спрашивает, а иначе как бы он смог узнать правильность ответа, когда он будет найден”» [5; 298]. Тем более важно подобрать такие массивы данных, которые затем однозначно должна воспроизвести модель. Цель этого этапа состоит по сути в подготовке результатов моделирования. «Целями являются предпочитаемые исходы...» [28; 70]. При подборе и фильтрации эмпирических данных подспудно решается вопрос, какой должна быть модель, чтобы воспроизвести имеющуюся информацию. По сути дела, располагая набором эмпирических данных, мы имеем решение, не зная пути к нему. Моделирование есть поэтому «регрессивное рассуждение», при котором «исходным пунктом анализа является то, что требуется доказать, ибо допускается, что задача уже решена…» (Папп, цит. по: [26; 136]). Подготовка данных для моделирования есть особенный, важнейший этап, во многом определяющий его успешность. Подбирая данные, мы придаем цель всему процессу исследования, в явной форме обозначаем то, к чему должна свестись эта работа. Данная цель предопределяет весь процесс детального исследования системного объекта. «Единством цели… объясняется то, что, приобретая знания, нельзя упустить из виду ни одной части, а также нельзя сделать никакого случайного добавления или остановиться на неопределенной величине совершенства, не имеющей a priori определенных границ. …Целое расчленено, а не нагромождено. …Полное целесообразное единство есть совершенство» [11; 486, 413]. (Важные соображения о финансовой и ресурсной стоимости исследовательских работ хоть и влияют на целеполагание, но не относятся собственно к логике моделирования.)
Построение модели (и обычно последующая ее перестройка) есть, с одной стороны, чисто технический процесс (ввод формул и данных в среду ПК). В то же время это глубокий и сложный системный процесс разработки блок-схемы и алгоритма модели. Множество возможных формул должно быть подобрано, исходя из имеющейся парадигмы и опыта решения аналогичных задач. Создание формализованного (блок-схема) и количественного (формулы) вариантов требует четкого понимания того, какие компоненты модели могут в ближайшем будущем измениться, какие должны обрести биологический смысл. Общее влияние цели на системное мышление (на реализацию системных принципов) состоит в том, что исследователь последовательно рассматривает разные варианты описания системы – включать или не включать те или иные элементы в модель, учитывать или не учитывать тот или иной уровень иерархии и т. п. Процесс конструирования модели – это процесс выбора альтернативных путей. Общее направление этапа постройки и перестройки определяет цель – создание в недрах компьютера работающей модели (множества формул, правильно выполняющих расчеты). (Имитационную систему невозможно построить, если не проводить предварительные расчеты, отлаживая обязательные связи между компонентами – формулами и данными, не обращая пока внимания на собственно результаты счета.) Здесь важно отметить следующее. Поскольку наши знания не могут безусловно претендовать на соответствие природе [15], одно и то же явление позволительно описывать с помощью самых разных типов моделей. Например, для изучения закономерностей динамики популяций мелких млекопитающих временной ряд численности можно разложить на гармоники [10], разделить на зимний и летний этапы функционирования [17] или представить множеством параллельных зависимых траекторий существования групп разновозрастных животных [13]. Таким образом, апробированный опыт автора есть одна из неявных целей моделирования.
Расчеты по модели – это отдельный процесс, хотя он осуществляется и на 2-м, и на 3-м этапах моделирования. Основываясь на внешней информации о состоянии среды и на стартовых
(начальных) характеристиках изучаемой биосистемы, модель призвана вычислить некие значения, которые далее интерпретируются как модельные аналоги признаков, наблюдаемых в природе. «Под моделью некоторого объекта понимается другой объект… который обладает существенными для целей моделирования свойствами и в рамках этих целей полностью заменяет исходный объект» [7; 12]. «Назначение любой из моделей организации поведения… чтобы их “поведение” воспроизводило то, что мы наблюдаем в действительности» [20; 141]. Как и в случае с функционированием организма, модель выдает только то, что она может вычислить, на что ориентирована ее конструкция. Ее объективная цель состоит в самореализации. «Общая характеристика особенности функционирования и структуры систем… < – это> наличие… целей функционирования систем, которые определяют ее целевое назначение» [30; 9]. Цель модели состоит в том, чтобы функционировать (причем по возможности так же, как и ее реальный прототип). При этом простые детерминистические (алгебраические) модели дают в прогонах одинаковые результаты. В более сложных стохастических и автоматных моделях однозначного воспроизведения результатов не наблюдается (при одинаковых стартовых значениях). В такие модели конструктивно заложена некоторая автономность траекторий жизни, имманентная изменчивость. Определенную свободу функционирования можно придать и алгебраическим моделям, если вносить в расчеты белый шум или варьировать объемы исходной информации. Однако стохастические модели должны давать статистически однородные результаты.
Настройка параметров (и структуры) модели во многом аналогична отбору (искусственному). Содержание процесса моделирования – это воплощения характеристик реального объекта в характеристики идеального объекта, перенос особенных черт строения и функционирования с прообраза на электронный дубликат. В математическом смысле цель процедуры сводится к тому, чтобы из множества возможных значений отобрать такие коэффициенты пропорциональности в модельных формулах, чтобы расчетный и эмпирический массивы данных по возможности совпали. С помощью специальных математических алгоритмов (обычно – оптимизации) параметры модели многократно и направленно модифицируются, и на каждом таком шаге проверяется соответствие двух массивов значений переменных – рассчитанных моделью и наблюдавшихся в природе. «Цель действий состоит в том, чтобы свести к минимуму промежуток… между существующей и желаемой системой...» [24; 92]. Предполагается, что хорошее соответствие этих массивов позволяет осмысленно интерпретировать модельные параметры, то есть дает понимание законов функционирования (какой-то определенной стороны) реальной биосистемы. Различие между наблюдениями и прогнозами по модели количественно оценивается. «В основу <моделирования> могут быть положены разные критерии подобия, основным из которых является изоморфизм» [2; 10]. «Критерий является средством, с помощью которого измеряются или выбираются альтернативы… Критерий есть стандарт, с помощью которого может быть вынесено суждение об относительной выгоде выбора» [24; 181]. Одни критерии, встроенные в процедуру проверки соответствия, служат для автоматического перебора и поиска лучших значений модельных параметров в процессе количественной настройки. Другие критерии оценивают общую адекватность модели наблюдаемым данным и являются основаниями для вывода о том, соответствует ли сама конструкция модельных формул объекту исследования, нужна ли структурная перестройка модели – возврат на второй этап (задачи идентификации). Наконец, третья группа критериев позволяет делать вывод о том, в состоянии ли данная модель описывать весь класс явлений, представленных объектом исследования (задачи верификации) [9]. Здесь возникает проблема точности всех этих критериев, целесообразность введения жестких или относительно мягких требований к модели. Основой для решения этого вопроса служит предварительное статистическое исследование феномена. Обычно в связи с высокой сложностью организации биосистем от биолого-экологических моделей не ожидается высокой точности. Можно полагать, что параметры модели обретают биологический смысл, когда коэффициент детерминации становится выше 0.6 (лучше – около 0.8). Даже если модель оказалась неадекватной, сама попытка ее построения всегда улучшает общее понимание научной проблемы и служит основанием для новых теоретических формулировок. В конце концов, основная цель научного исследования – приближение к знанию законов природы, а неудачные модели являются неизбежными шагами в этом направлении. Иными словами, вопрос об успешности исследования решается с привлечением дополнительных критериев. «…Случаи, когда единственный критерий удачно отображает цель, скорее исключения» [25; 319].
Если построенная модель соответствует выбранным критериям, то можно считать, что цель достигнута – технический процесс моделирования завершен. Теперь остается дать ее интерпретацию, теоретическое объяснение полученным коэффициентам пропорциональности – параметрам модели. Именно параметры являются важнейшим результатом модельного исследования, по скольку они с максимальной определенностью характеризуют изучаемый процесс. Эти показатели используются для придания большей определенности ранее построенным теоретическим конструкциям. Процесс интерпретации естественно включается в ранее построенную схему системного исследования [15] (рис. 1, 4 ).
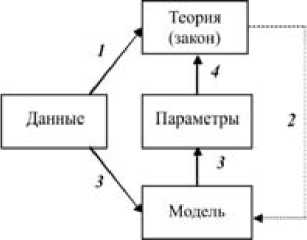
Рис. 1. Логическая схема модельного исследования:
1 – Обобщение накопленных данных в форме первичных гипотез и теорий.
2 – Построение структуры моделей.
3 – Настройка модели (оценка параметров).
4 – Интерпретация параметров модели, включение в теорию.
В качестве примера можно рассмотреть наши исследования по термобиологии обыкновенной гадюки [16]. Теоретически понятная картина (пойкилотермная рептилия нагревается на солнцепеке до оптимальных температур, чтобы активизировать обменные процессы) была существенно дополнена несколькими точно оцененными модельными параметрами (максимальная и минимальная добровольная температура, температурная чувствительность). В результате изменились и теоретические представления об этом процессе. Максимальная температура тела гадюки (35 °С), изредка наблюдаемая в природе, не является в норме оптимальной, но выполняет роль целевого параметра, к которому животное стремится, но достичь которого в условиях Карелии не может. Однако само стремление к этому порогу обеспечивает поддержание температуры тела на оптимальном уровне (28–30 ºС).
Дальнейшая работа с моделью состоит в том, чтобы выяснить, как она реагирует на варьирование исходных данных или значений параметров . Этот этап называют «модельный эксперимент». Модель выступает полным заместителем природного объекта и характеризует его возможную реакцию на разного рода «возмущения среды». В нашем примере были исследованы кривые динамики температуры тела разновозрастных и разнополых животных, терморегуляторное поведение которых, как выяснилось, контролируется только одним названным параметром, максимальной добровольной температурой (у взрослых она выше, у молодых ниже, у беременных самок выше всех) [16].
Здесь появляется возможность для наиболее интересного варианта интерпретации – получить прогноз поведения биосистемы при определенном воздействии. Модельные эксперименты могут послужить прообразом тех действий, которые затем в реальности позволят оптимизировать биосистему (рис. 2).
Эта схема соответствует и прикладным проблемам экологии, если в качестве объекта исследования рассматривать угрожаемые экосистемы и процессы антропогенного вмешательства в природу.
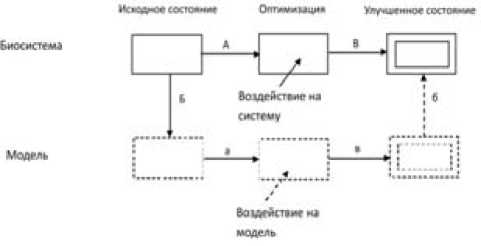
Рис. 2. Соотношение целей системы и модели в процессе моделирования:
А – существование системы,
Б – целесообразная проекция структуры системы на структуру модели, а – существование модели, в – улучшение состояния модели после экспериментального воздействия, б – проекция поведения модели на поведение системы.
В – улучшение состояния системы после воздействия по рекомендациям моделирования.
В заключение следует еще раз подчеркнуть, что на любое системное представление объекта исследования (в форме фрейма и модели) накладывают свой отпечаток как объективные цели изучаемой биосистемы, так и собственные (часто непонятые) цели исследования.
Чаще всего наблюдается бессознательное подчинение доминирующей парадигме. Для создания оригинальных моделей необходимо ознакомиться с разнообразными взглядами на предмет исследований, с разными теориями, концепциями, подходами, то есть явно определиться с собственными теоретическими воззрениями.
Не менее обычно и стремление описать «очевидные» объективные «биологические цели», которые видны при поверхностном (привычном, традиционном) подходе к биологическим процессам. Для того чтобы разработать наиболее адекватный фрейм биосистемы, следует выполнить полноценное системное исследование – рассмотреть разные уровни иерархии объектов и процессов, способы интеграции и поддержания целостности, гомеостаза, выявить эмерджентности и т. п.
Наконец, предзаданной целевой установкой является использование привычных приемов моделирования. Чтобы не ограничивать себя в средствах моделирования, важно изучить различные его методы, в том числе криволинейную регрессию, многомерную статистику, анализ временных рядов, методы шкалирования, алгебраические и стохастические имитационные модели, автоматы, фракталы и пр. и на этой основе выбрать подходящую основу для формализации.
Практика целеполагания при моделировании состоит в том, чтобы выявлять и записывать все возможные цели исследования и только после этого анализа определяться со списком действительно важных целей.
Список литературы Принцип целесообразности и моделирование биосистем
- Анохин П. К. Системные механизмы высшей нервной деятельности. М.: Наука, 1979. 453 с.
- Антомонов Ю. Г. Моделирование биологических систем: Справочник. Киев: Наукова думка, 1977. 260 с.
- Беловски Г. Е. Модели выживания и продолжительность существования популяций//Жизнеспособность популяций. Природоохранные аспекты. М.: Мир, 1989. С. 53-80.
- Берг Л. С. Труды по теории эволюции. 1922-1930. Л.: Наука, 1977. 387 с.
- Бэкон Ф. Сочинения: В 2 т. Т. 1. О достоинстве и приумножении наук. М.: Мысль, 1977. 567 с.
- Винер Н. Творец и Голем, INC//Винер Н. Творец и будущее. М.: Изд-во АСТ, 2003. С. 225-292.
- Гаазе-Раппопорт М. Г., Поспелов Д. А. От амебы до робота: Модели поведения. М.: Наука, 1987. 288 с.
- Гиляров А. М. Динамика численности пресноводных планктонных ракообразных. М.: Наука, 1987. 190 с.
- Иванищев В. В., Михайлов В. В., Тубольцева В. В. Инженерная экология. Л.: Наука, 1989. 144 с.
- Ивантер Э. В., Коросов А. В., Якимова А. Е. Статистический анализ многолетних изменений численности мелких млекопитающих северо-восточного Приладожья//Вопросы популяционной биологии. Труды ПетрГУ. Сер. биол. Вып. 2. Петрозаводск: Изд-во ПетрГУ, 2008. С. 5-24.
- Кант И. Критика чистого разума. М.: Мысль, 1994. 591 с.
- Коросов А. В. Имитация экологических объектов в среде пакета Microsoft Excel//Экология. 2002. № 2. С. 144-147.
- Коросов А. В. Имитационное моделирование в среде MS Excel (на примерах из экологии). Петрозаводск: Изд-во ПетрГУ, 2002. 212 с. (http://elibrary.karelia.ru/book.shtml?levelID=017002&id=612&cType=1)
- Коросов А. В. Двухконтурная отрицательная обратная связь и модель терморегуляции гадюки//Ученые записки Петрозаводского государственного университета. Сер. Естественные и технические науки. 2008. № 1. С. 74-82.
- Коросов А. В. Системы и экосистемы//Ученые записки Петрозаводского государственного университета. Сер. Естественные и технические науки. 2008. № 2. С. 58-67.
- Коросов А. В. Простая модель баскинга обыкновенной гадюки (Vipera berus L.)//Современная герпетология. 2008. Т. 8. Вып. 2. С. 118-136.
- Коросов А. В., Зорина А. А. Исследование динамики численности рыжей полевки с помощью функций последования//Экология. 2007. 1. С. 49-54.
- Криксунов Е. А. Теория динамики промыслового стада рыб. М.: Изд-во МГУ, 1991. 77 с.
- Любищев А. А. Проблемы формы, систематики и эволюции организмов. М.: Наука, 1980. 280 с.
- Меннинг О. Поведение животных. Вводный курс. М.: Мир, 1982. 360 с.
- Меншуткин В. В. Эволюционный подход к имитационному моделированию популяций//Математические методы в экологии: Тезисы докладов Третьей Всероссийской школы молодых ученых. Петрозаводск, 24-29 августа 2008 г. Петрозаводск: КарНЦ РАН, 2008. C. 21-23.
- Мина М. В., Клевезаль Г. А. Рост животных. Анализ на уровне организма. М.: Наука, 1976. 291 с.
- Мосияш С. С., Шашуловский В. А. Использование итерационного моделирования для прогнозирования допустимой промысловой эксплуатации популяции рыб//Поволжский экологический журнал. 2003. № 2. С. 190-194.
- Оптнер С. Л. Системный анализ для решения деловых и промышленных проблем. М.: Советское радио, 1969. 216 с.
- Перегудов Ф. И., Тарасенко Ф. П. Введение в системный анализ. М.: Высшая школа, 1989. 367 с.
- Пойа Д. Как решать задачи. Львов: Квантор, 1991. 216 с.
- Разумовский О. С. Бихевиоральные системы. Новосибирск: Наука, 1993. 240 с.
- Саати Т., Кернс К. Аналитическое планирование. Организация систем. М.: Радио и связь, 1991. 224 с.
- Сеннер Дж. В. Инбредная депрессия и выживание популяций в зоопарках//Биология охраны природы. М.: Мир, 1983. С. 238-254.
- Тараканов К. В., Овчаров С. А., Тырышкин А. Н. Аналитические методы исследования систем. М.: Сов. Радио, 1974. 239 с.
- Уиттекер Р. Сообщества и экосистемы. М.: Прогресс, 1975. 328 с.
- Цигичко В. Н., Клоков В. В. Основные принципы описания сложных организационных систем//Диалектика и системный анализ. М.: Наука, 1986. С. 121-136.
- Шварц С. С., Смирнов В. С., Добринский Л. Н. Метод морфофизиологических индикаторов в экологии наземных позвоночных. Свердловск, 1968. 387 с.
- Berry R. J. Epigenetic polymorphism in wild population of Mus muscullus//Genetical Reseach. Cambr. 1963. Vol. 4. P. 193-220.
