Приоритезация генов нейронального апоптоза по их структурной роли в ассоциативной генной сети нарушений аутического спектра с помощью подходов ANDSystem

Автор: Лаврик И.Н., Иванисенко В.А., Деменков П.С., Сайк О.В., Янкина М.А.
Журнал: Международный журнал гуманитарных и естественных наук @intjournal
Рубрика: Биологические науки
Статья в выпуске: 9 (12), 2017 года.
Бесплатный доступ
Нарушения аутистического спектра (НАС) по оценкам эпидемиологических исследований могут поражать до 1% населения в мире. НАС сопровождается проблемами в социальной коммуникации, наличием ограниченных интересов, стереотипного и повторяющегося поведения. К числу важных факторов патогенеза НАС относится нейрональный апоптоз. В данной работе проведена приоритезация генов, вовлеченных в биологический процесс Gene Ontology нейрональный апоптоз, по их потенциальной важности для патогенеза НАС. Были использованы стандартные методы приоритезации ToppGene и Endeavor, а также подходы, реализованные в системе ANDSystem, учитывающие структуру генной сети НАС. Анализ показал, что наиболее перспективными для дальнейшего исследования в экспериментах по генотипированию могут быть гены GRIN1, NTRK2, GRIK5, PTK2B, CTNNB1 и ADORA2A.
Нейрональный апоптоз, нарушения аутистического спектра (нас), ассоциативные генные сети, приоритезация генов, гены-кандидаты
Короткий адрес: https://sciup.org/170190348
IDR: 170190348
Prioritization of neuronal apoptosis genes by their structural role in the associative gene network of autism spectrum disorder using ANDSystem approaches
Autism spectrum disorder (ASD) can affect up to 1% of the world's population according to epidemiological studies. ASD is accompanied by problems in social communication, the presence of limited interests, stereotyped and repetitive behavior. Among the important factors of ASD pathogenesis is neuronal apoptosis. In this work, genes involved in the Gene Ontology biological process neuronal apoptosis were prioritized according to their potential importance for ASD pathogenesis. For gene prioritization were used standard methods ToppGene and Endeavor, as well as approaches, taking into account the structure of the ASD gene network, implemented in the ANDSystem tool. The analysis showed that genes GRIN1, NTRK2, GRIK5, PTK2B, CTNNB1 and ADORA2A could be the most promising for further research in genotyping experiments.
Текст научной статьи Приоритезация генов нейронального апоптоза по их структурной роли в ассоциативной генной сети нарушений аутического спектра с помощью подходов ANDSystem
НАС [3]. Исследования показывают, что одним из важных факторов патогенеза НАС выступает нейрональный апоптоз [46].
Нейрональный апоптоз является одной из форм клеточной гибели и характеризуется блеббингом и потерей асимметрии плазматической мембраны, конденсацией хроматина и расщеплением ДНК. Во время эмбрионального развития этот процесс является критическим для обеспечения элиминации избыточного числа нейронов [7]. При ряде заболеваний, таких как болезнь Альцгеймера, эпилепсия, судорожные расстройства, нейротравмы и др. наблюдаются нарушения нейронального апоптоза [810].
Целью данной работы была приоритезация генов нейронального апоптоза по их потенциальной важности для НАС с помощью стандартных методов ToppGene [11] и Endeavor [12], а также критериев специфичности и центральности, реализованных в разработанной нами ранее системе ANDSystem [13, 14]. Критерии специфичности и центральности ANDSystem позволяют учитывать структуру генной сети НАС. На основе проведенного анализа были выявлены новые гены-кандидаты, перспективные для дальнейшего исследования в экспериментах по генотипированию.
Материалы и методы
Список генов, ассоциированных с НАС, был экстрагирован из базы данных MalaCards [15] по запросу «Autism Spectrum Disorder». Список генов нейронального апоптоза был экстрагирован из базы данных AmiGO 2 [16] по запросу «neuron apoptotic process» (GO:0051402).
Ассоциативные генные сети были реконструированы с помощью системы ANDSystem [13, 14]. Система ANDSystem позволяет строить сети молекулярногенетических взаимодействий между генами, белками, микроРНК, метаболитами и другими биологическими объектами в норме и при различных заболеваниях [1725].
Для приоритезации использовались программы ToppGene [11] и Endeavour version: 3.71 [12]. На вход программам подавался тренировочный и тестовый наборы генов. Тренировочный набор включал гены, ассоциированные с НАС, за исключением генов, одновременно ассоциированных с НАС и входящих в нейрональный апоптоз. Тестовый набор включал все гены нейронального апоптоза. Настройки программ использовались по умолчанию. Для ранжирования генов использовались ранги, рассчитываемые программами ToppGene и Endeavour.
Приоритезация с помощью системы ANDSystem проводилась по двум показателям: центральности CTC (cross-talk centrality) и специфичности CTS (cross-talk specificity), которые были рассчитаны с помощью функции “Intelligent Filtration”, реализованной в программе “ANDVisio” системы ANDSystem. Для расчета центральности CTC использовалась формула: CTCi=Ni/M, где Ni – число связей i-го гена с участниками ассоциативной генной сети НАС, в глобальной сети (базе знаний) системы ANDSystem; M – число участников (вершин) ассоциативной генной сети НАС. Специфичность CTS рассчитывалась как CTSi=Ni/Ki, где Ki – общее число связей i-го гена в глобальной сети (базе знаний) ANDSystem. Гены ранжировались согласно убыванию показателей центральности CTC и специфичности CTS. Наименьшее значение ранга (наибольший приоритет гена) соответствовало наибольшему показателю центральности CTC (специфичности CTS).
Средний ранг рассчитывался как среднее арифметическое значение четырех рангов критериев ToppGene, Endeavor, центральность CTC и специфичность CTS.
Обогащенность списка наиболее приоритетных генов генами, ассоциированными с НАС, оценивалась согласно гипергеометрическому распределению с помощью веб-ресурса GeneProf .
Анализ перепредставленности участниками биологических процессов для наборов генов проводился с помощью системы DAVID 6.8 [26], p-value с поправкой Бенджамини—Хохберга на множественность сравнений менее 0.05.
Результаты и обсуждение
В настоящей работе рассматривался список из 98 генов, ассоциированных с НАС согласно информации из базы данных Malacards [15]. С помощью системы ANDSystem [13, 14] между этими генами и их продуктами была реконструирована ассоциативная генная сеть НАС. Из 98 генов 40 оказались не связанными в ассоциативной генной сети НАС. Между остальными 58 генами/белками было найдено 377 взаимодействий, включая 248 ассоциативных связей, 58 связей типа «экспрессия гена», 53 белок-белок взаимодействия и 18 регуляторных связей (см. рисунок).
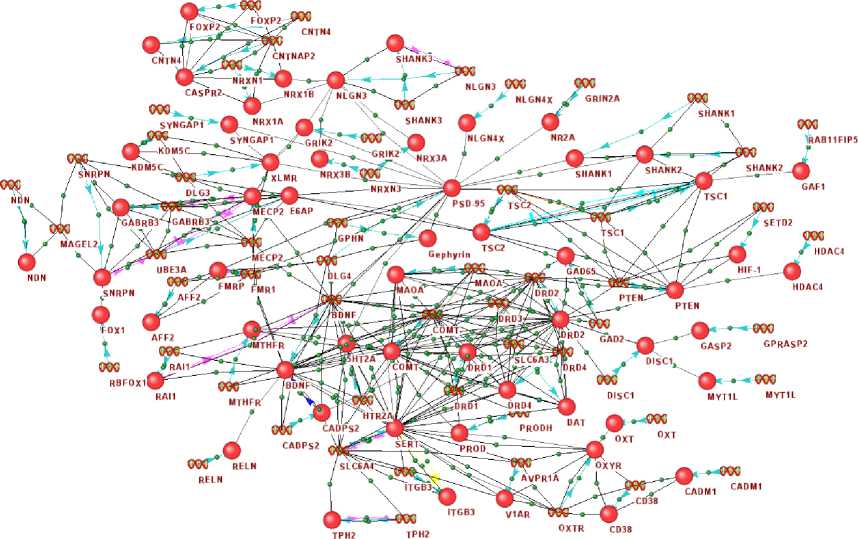
Рисунок 1. Ассоциативная генная сеть НАС, реконструированная с помощью системы ANDSystem
На основании информации из базы данных AmiGO 2 [16] был сформирован список из 218 генов, вовлеченных в нейрональный апоптоз. Оказалось, что из них 11 генов (ADNP, BDNF, DLX1, EN2, GABRA5, GABRB3, GRIK2, HDAC4, MECP2, SCN2A и SYNGAP1) входят в ассоциативную генную сеть НАС. Эти гены принимают участие в различных биологических процессах. Например, гены ADNP, BDNF и MECP2 участвуют в биологических процессах Gene Ontology, отвечающих за процессы, связанные с формирова- нием памяти; BDNF, DLX1, EN2, GABRA5, GABRB3, HDAC4, MECP2 и SYNGAP1 – в развитии нервной системы и структур мозга; GRIK2 – в механизмах ответа на страх; SCN2A – в миелинизации и др.
Приоритезация генов нейронального апоптоза проводилась с помощью стандартных методов ToppGene [11] и Endeavor [12], а также показателей центральности CTC и специфичности CTS системы ANDSystem. Список 10 наиболее приоритетных генов приведен в таблице.
Таблица 1. Наиболее приоритетные гены нейронального апоптоза, выявленные соглас- но четырем критериям (ToppGene, Endeavor, центральность CTC и специфичность CTS)
|
Ген |
GeneId |
Ранг ToppGene |
Ранг Endevaour |
Ранг CTC |
Ранг CTS |
Средний ранг |
|
GRIN1 |
2902 |
2 |
1 |
4,5 |
8 |
3,875 |
|
GABRB3* |
2562 |
1 |
4 |
22,5 |
2 |
7,375 |
|
NTRK2 |
4915 |
6 |
2 |
22,5 |
17 |
11,875 |
|
BDNF* |
627 |
8 |
26 |
2 |
19 |
13,75 |
|
GRIK2* |
2898 |
3 |
3 |
56,5 |
7 |
17,375 |
|
MECP2* |
4204 |
20 |
7 |
22,5 |
26 |
18,875 |
|
GRIK5 |
2901 |
11 |
6 |
56,5 |
9 |
20,625 |
|
PTK2B |
2185 |
13 |
38 |
15 |
27 |
23,25 |
|
CTNNB1 |
1499 |
34 |
30 |
3 |
38 |
26,25 |
|
ADORA2A |
135 |
18 |
11 |
56,5 |
20 |
26,375 |
|
TP53# |
7157 |
77 |
27 |
1 |
80 |
46,25 |
|
TFAP2B# |
7021 |
65 |
141 |
56,5 |
1 |
65,875 |
Примечание: * гены, ассоциированные с НАС; # гены, не входящие в список из 10 наиболее приоритетных по среднему рангу, но обладающие наилучшим рангом по отдельно взятым критериям центральность CTC и специфичность CTS.
Оказалось, что в числе 10 наиболее приоритетных генов нейронального апоптоза статистически значимо перепредстав-лены гены (BDNF, GABRB3, GRIK2, MECP2), ассоциированные с НАС, (p-value< 0.001). Наиболее интересными для дальнейшего изучения являются шесть генов (GRIN1, NTRK2, GRIK5, PTK2B, CTNNB1, ADORA2A), которые оказались в числе 10 наиболее приоритетных и не были ассоциированы с НАС по данным MalaCards. Анализ сверхпредставленности биологических процессов DAVID показал, что эти гены преимущественно вовлечены в отрицательную регуляцию нейронального апоптоза (GO:0043524), глутаматерги-ческий синаптическую передачу сигнала (GO:0035235, GO:0035249) и появление возбуждающего постсинаптического потенциала (GO:0060079). Анализ литературы показал, что для генов GRIN1, NTRK2, PTK2B, CTNNB1 и ADORA2A уже есть сведения о возможной взаимосвязи мутаций в этих генах с НАС [27-31]. Для гена GRIK5 данных об ассоциации с НАС в системе PubMed найти не удалось. Этот ген кодирует рецептор глутамата и вовлечен в синаптическую передачу [32]. Мута- ции в гене GRIK5 ассоциированы с шизофренией [33].
Гены TP53 и TFAP2B не попали в список из 10 наиболее приоритетных по среднему рангу, но имели первый ранг по отдельно взятым критериям центральность CTC и специфичность CTS, соответственно. Ген TP53 является супрессором опухолей и вовлечен в регуляцию клеточного цикла [34]. Ген TFAP2B кодирует сайт-специфичный ДНК-связывающий белок, который участвует в широком спектре важных биологических процессов, включая развитие глаз, лица, тела, конечностей, почечного эпителия и нервной трубки [35]. Дальнейшее изучение этих генов может быть интересным для выявления молекулярно-генетических механизмов патогенеза НАС.
Заключение
Анализ приоритезации генов нейронального апоптоза с учетом их структурной роли в генной сети НАС, проведенной с помощью методов ToppGene, Endeavor и ANDSystem, позволил выявить шесть генов (GRIN1, NTRK2, GRIK5, PTK2B,
CTNNB1, ADORA2A), наиболее перспективных для их дальнейшего экспериментального изучения в патогенезе НАС.
Список литературы Приоритезация генов нейронального апоптоза по их структурной роли в ассоциативной генной сети нарушений аутического спектра с помощью подходов ANDSystem
- Grice D. E., Buxbaum J. D. The genetics of autism spectrum disorders//Neuromolecular medicine. -2006. -Т. 8. -№4. -С. 451-460.
- Developmental D. M. N. S. Y. et al. Prevalence of autism spectrum disorder among children aged 8 years-autism and developmental disabilities monitoring network, 11 sites, United States, 2010//Morbidity and mortality weekly report. Surveillance summaries (Washington, DC: 2002). -2014. -Т. 63. -№2. -С. 1.
- Bourgeron T. From the genetic architecture to synaptic plasticity in autism spectrum disorder//Nature Reviews. Neuroscience. -2015. -Т. 16. -№9. -С. 551.
- Ghanizadeh A. Targeting neurotensin as a potential novel approach for the treatment of autism//Journal of neuroinflammation. -2010. -Т. 7. -№1. -С. 58.
- Malik M., Sheikh A. M., Wen G., Spivack W., Brown W. T., Li X. Expression of inflammatory cytokines, Bcl2 and cathepsin D are altered in lymphoblasts of autistic subjects//Immunobiology. -2011. -Т. 216. -№1. -С. 80-85.
