Природно-географические особенности экологии барсука (Meles Brisson, 1762)
")
Автор: Дворников М.Г., Чащин П.В.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Наземные экосистемы
Статья в выпуске: 2 т.10, 2008 года.
Бесплатный доступ
Исследованы некоторые аспекты экологии барсука (Meles Brisson, 1762) в Заволжье, Урале и Зауралье. Выявлены особенности поселений, питания, воздействия на среду и плотности населения животных в лесной и лесостепной зонах.
Экология, барсук
Короткий адрес: https://sciup.org/148198114
IDR: 148198114 | УДК: 591.5:599.742.43
Biogeographic features of the badger (Meles Brisson, 1762) ecology
Some aspects of badger (Meles Brisson, 1762) ecology in the Trans-Volga, Urals and Trans-Urals regions are investigated. The features of settlements, diet, influence on environment and population density of animals in the forest and forest-steppe zones are revealed. They are connected to the hydrothermal, geomorphological and anthropogenous factors.
Текст научной статьи Природно-географические особенности экологии барсука (Meles Brisson, 1762)
Барсуки ( Meles Brisson, 1762) – норные животные. Они занимают постоянный участок обитания, обеспечивающий их кормовыми ресурсами, убежищами и, главное, условиями для устройства выводковых нор. В настоящее время представители рода Meles широко распространены в Евразии. В Заволжье, Урале и Зауралье они обычны и местами многочисленны. Датировка костных остатков барсуков, обнаруженных в пещерах и на стоянках древнего человека, позволяет считать, что барсуки появились здесь в конце плейстоцена – начале голоцена [5, 13, 14, 19, 23, 25, 26, 27]. Произошедшие в голоцене глобальные изменения климата [15] предопределили сложившуюся к настоящему времени природно-географическую зональность и разнообразие современных экосистем. Это разнообразие сформировалось в рассматриваемом регионе не только под воздействием вековых изменений климата, но и в ряде случаев вследствие антропогенного воздействия в исторический период [31, 36]. Логично предположить, что на столь обширной территории, охватывающей несколько природно-географических зон, существуют определенные различия в экологии барсуков.
Основной целью нашего исследования было выявление региональной (природногеографической) специфики некоторых аспектов экологии барсука, наиболее тесно связанных с норной и кормодобывающей деятельностью. Следует отметить, что в Заволжье обитают два вида барсуков: европейский
Meles meles L. и азиатский Meles leucurus Нodgson [1, 2]. Предполагается, что имеется определенная видоспецифичность в экологии этих видов [6]. Однако эта сторона вопроса остается за рамками нашего исследования, и мы рассматриваем проблему в масштабах рода.
Материал и методы
Сбор материала велся в течение 1972-2006 гг. в Заволжье, Урале и Зауралье. Работы проводили на условно ненарушенных, особо охраняемых природных территориях (ООПТ) различного ранга. Для сравнения обследовали прилегающие к ООПТ антропогенно нарушенные территории охотничьих хозяйств, на землях которых ведется охота, сельскохозяйственная и лесохозяйственная деятельность. Район основных исследований охватывал лесную и лесостепную зоны (табл. 1).
На указанных территориях через 250-300 м прокладывали линейные маршруты, суммарная протяженность которых составила 553 км. На маршрутах регистрировали жилые и покинутые поселения (n = 282), временные норы и другие следы жизнедеятельности барсуков, в том числе кормовые покопки, «туалеты» и другое, рассматривая их как сигнальные территориальные метки [22]. В заповедниках, в дополнение к оригинальным, использовали архивные данные о встречах барсуков и следов их жизнедеятельности, зафиксированные в карточках встреч животных и их следов (n = 585). Все полученные сведения наносили на планы лесонасаждений М
1:25 000.
Спектр питания определяли по остаткам пищи в экскрементах, пробы которых брали в июне-сентябре (n = 426). В Ильменском заповеднике определяли наличие в составе питания дождевых червей. Для этого использовали общепринятые и оригинальные методики [28, 35, 37]. Пробы брали на постоян- ном маршруте в апреле-октябре (n = 254).
Для определения индивидуального (семейного) участка у нор выкладывали кормовые приманки из теста, обжаренного в растительном масле с начинкой из мелких фрагментов цветного пластика. По содержанию пластика в экскрементах оценивали освоение территории барсуками [9].
Таблица 1. Районы основных исследований
|
Природно-географическое деление |
Административная принадлежность, широта |
Наименование |
Площадь, км2 |
|
Лесная зона |
|||
|
Подзона южной тайги |
Кировская область 58º00" с.ш. |
Заповедник «Нургуш» и прилегающая территория |
900 |
|
Подзона сосново-березовых лесов восточного склона Урала |
Челябинская область 55º10" с.ш. |
Ильменский заповедник |
300 |
|
Лесостепная зона |
|||
|
Подзона южной лесостепи Европейской равнины |
Республика Башкортостан 53º50" с.ш. |
Бижбулякский заказник |
145 |
|
Подзона южной лесостепи зауральского пенеплена |
Челябинская область 54º00" - 54º40" с.ш. |
Варламовский, Санарский, Троицкий, Уйский заказники |
557 |
В длительно используемых, «вековых» поселениях оценивали воздействие барсуков на травостой. Для этого на территории барсучьих «городков» определяли видовой состав, проективное покрытие и запас фитомассы. Полученные данные сравнивали с аналогичными участками, на которых следы деятельности барсука отсутствовали [12].
Кластерный анализ методом невзвешенных средних (UPGMA) проводили с помощью пакета Statistica 6.0 for Windows.
Результаты и обсуждение
Плотность населения барсуков, количество и мощность поселений. Показатели, характеризующие население барсуков в пределах рассматриваемых природно-географических зон, заметно различаются (табл. 2). Для визуализации информации на основе табличных данных была построена дендрограмма (рис.).
На дендрограмме хорошо выделяются три кластера. Видно, что первый кластер образуют пойменные, террасные и водораздельные участки заповедника «Нургуш» и горный участок Ильменского заповедника. Здесь наиболее низкая плотность населения и наимень- шее количество потенциально жилых поселений (табл. 2). Наиболее обособлен кластер предгорной части Ильменского заповедника. На этой территории показатели, характеризующие численность животных и степень развития поселений, в целом самые высокие (табл. 2). Отдельный кластер образуют заказники лесостепной зоны. Характеристики участков, относящихся к лесостепной зоне, имеют промежуточные значения (табл. 2).
Таким образом, плотность населения барсуков, количество и мощность их поселений в направлении с севера на юг в пределах лесной зоны увеличиваются. Далее, в лесостепной зоне, отмечено снижение показателей. Выявленные различия мы связываем с несколькими основными факторами – климатическим, геоморфологическим и антропогенным.
Климат. Известно, что биопродуктивность экосистем (фитомасса и биомасса животных) связана с гидротермическим фактором. В границах лесной зоны повышение количества поступающей солнечной радиации в направлении с севера на юг [3] обусловливает увеличение в том же направлении био- продуктивности [33] и, в конечном счете, кормовых ресурсов. Далее, в лесостепной зоне, по мере снижения увлажненности биопродуктивность начинает снижаться.
Как видно из табл. 2, изменение плотности населения барсуков соответствует указанной закономерности. В направлении с севера на юг плотность населения барсуков сначала возрастает, а затем снижается. В рассматриваемом ряду данных, с точки зрения обеспеченности кормовыми ресурсами, наиболее благоприятны районы на границе лесной и лесостепной зон (Ильменский заповедник). Как уже отмечалось, здесь зарегистрирована наибольшая плотность населения животных. Необходимо заметить, что в лесной зоне, вектор повышения биопродуктивности экосистем не всегда совпадает с направлением север – юг. Например, в Ильменском заповеднике (низкогорные предлесостепные сосново-березовые леса Южного Урала) в свежих типах леса у подножий и нижних частах склона гор обилие видов и биомасса беспозвоночных выше, чем в средних и верхних [16, 17, 21]. Однако высотная поясность здесь отсутствует. В данном случае биопродуктивность возрастает в соответствии с улучшением режима увлажнения [39].
Таблица 2. Плотность населения барсуков и характеристика поселений в лесной и лесостепной зонах
|
Природные зоны и подзоны |
Район исследований |
8 5 5 i м il i|T |
hi |
|^^ E Ills |
h E |
|
Лесная зона, подзона южной тайги |
Заповедник «Нургуш» (пойма р. Вятка) Охранная зона заповедника (терраса) Охотхозяйство (водораздел) |
0,54 0,76 0,50 |
1,1 0,44 0,3 |
5-11 / 7,7 3-9 / 7,2 + 0,9 6-10 / 6,9 + 0,2 |
150-300 |
|
Лесная зона, подзона сосновоберезовых лесов восточного склона Урала |
Заповедник Ильменский, горная часть Предгорная часть |
1,8 2,5 |
5,6 10,0 |
2-6 / 3,5 + 0,4 22-43/29,3 + 1,3 |
100-150 150-600 |
|
Лесостепная зона, подзона южной лесостепи |
Заказники Башкортостана и Челябинской области |
2,85 |
1,2-6,0 |
8-24 / 19,8 + 0,9 |
500-600 |
Сходное направление вектора биопродуктивности отмечено в сопряженных экосистемах вдоль поперечного профиля долины р. Вятка [11]. Здесь биопродуктивность повышается в направлении водораздел – припойменная терраса – пойма. С более высокой продуктивностью напрямую связана плотность населения барсуков.
Как в лесной, так и в лесостепной зонах, барсуки используют кормовой ярус, состоящий главным образом из травостоя, опада и подстилки. Иногда (Ильменский заповедник) в кормовой ярус входят подлесок (вишня, рябина) и нижний ярус древостоя (рябина, че- ремуха). При этом кормовые покопки зверей в основном приурочены к мозаичным лесонасаждениям с сочетанием лугов и редколесий.
Анализ встречаемости в экскрементах остатков различных видов кормов показал, что барсуки европейской тайги менее плотоядны [10], чем на Урале, что можно объяснить более низкой биопродуктивностью среднетаежных экосистем. В бассейне р. Вятка остатки мелких млекопитающих в экскрементах барсуков составляли 1,6%, птиц – 1,6%, амфибий – 18,7%, пресмыкающихся – 6,3%, насекомых – 20,3%, растительных кормов – 51,5%;
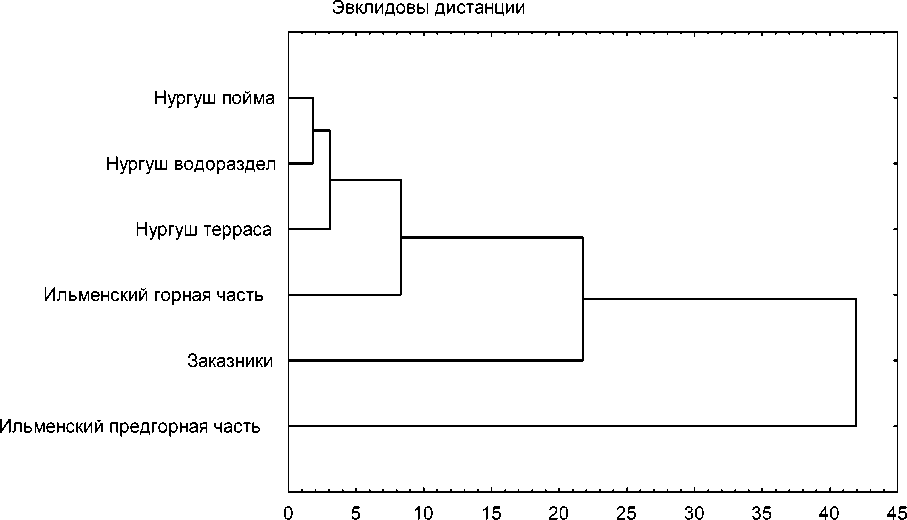
Рис. Кластерный анализ (UPGMA-метод) характеристик поселений и плотности населения барсуков в лесной и лесостепной зона в юго-восточной части Урала – 7,8%, 2,2%, 20,5%, 1,1%, 31,2% и 37,2% соответственно.
В Ильменском заповеднике, кроме перечисленных видов кормов, в составе питания существенную роль играют дождевые черви. Например, в 2001 г. остатки (щетинки) дождевых червей в большей или меньшей степени встречались в 73% проб, а в 2004 г. – в 81% проб [28, 35].
Биопродуктивность используемых барсуками экосистем определяет размер индивидуального (семейного) участка. В Ильменском заповеднике наибольший радиус удаления зверей от обитаемых нор составляет 3,5 км. В заповеднике «Нургуш» он намного больше и составляет в среднем около 8 км. В редких случаях мы отмечали удаление барсуков от поселения до 18 км.
Геоморфология. Как уже упоминалось, барсуки занимают определенный участок обитания, обеспечивающий их кормовыми ресурсами и условиями для устройства выводковых нор. Возможность устройства последних является лимитирующим фактором [34] и объясняет длительное существование отдельных поселений в течение нескольких тысячелетий [13]. Возможность устройства таких поселений обуславливается природны- ми условиями, прежде всего геоморфологией местности [20, 34]. Например, в условиях Ильменского заповедника поселения естественного происхождение расположены в самых различных условиях: на вершинах, различной крутизны склонах и у подножий хребтов и холмов, в межгорных понижениях, долинах рек и ручьев, на склонах оврагов и т.д. При этом большая часть поселений устроено животными в камнях – скальных обнажениях, каменных россыпях и т.п. Это объясняется небольшой мощностью аллювиального слоя на склонах гор и заболоченностью долин. В отличие от европейской территории страны [20] расположение поселений меньше связано с высотой над уровнем моря.
Антропогенное воздействие. В прошлом, в период возникновения большинства посе лений, хозяйственная (охотничья) деятельность человека заметно не ограничивала эти возможности. В настоящее время ситуация изменилась. По этой причине на территориях ООПТ количество «потенциально жилых» поселений заметно больше, чем на сопредельных территориях. Здесь прослеживается и более высокая плотность населения животных.
В Ильменском заповеднике, расположен- ном на границе лесной и лесостепной зон, большая часть «вековых» поселений находится в скальных образованиях и в большинстве случаев не могут быть разрушены человеком. В окрестностях заповедника также имеются большие древние поселения, как в лесной зоне, так и в лесостепи. Однако барсук здесь редок, так как подвергается постоянному преследованию. Норы зверей, где было возможно, раскапывались, либо животных выкуривали дымом [32].
На территориях ООПТ лесостепной зоны Зауралья количество поселений выше, чем в горно-лесной зоне Южного Урала (табл. 2). Например, в Троицком заказнике (экотон южной лесостепи и северной степи) на площади 1200 га выявлено 8 поселений, 4 из них были отнесены к «потенциально жилым». Сходные результаты были получены нами и для других лесостепных ООПТ Зауралья. В лесостепной зоне к западу от Урала (в Заволжье) также отмечается большое число поселений и высокая численность барсука [8].
В зоне средней тайги плотность населения барсука значительно ниже [10]. Всего в заповеднике и на прилегающей территории выявлено 54 поселения (в среднем 0,6 на 1000 га). Причем в несвойственных, пойменных сообществах (территория ООПТ, заповедник «Нургуш») плотность населения барсука выше, чем на сопредельных территориях. Очевидно, сказывается природоохранный статус этой территории.
Установлено, что ежегодно барсуками используются и «ремонтируются» не все «потенциально жилые» (выводковые) поселения. Однако все они носят следы раскопок человеком. В заповеднике «Нургуш» и его охранной зоне, прежде всего в пойме, поселения располагаются на незаливаемых половодьем гривах. При появлении в сентябре-октябре следов человека у поселений барсуки покидают подготовленные к зимовке норы. В то же время в обжитой человеком местности единично, с плотностью 0,09 на 1000 га, встречаются крупные поселения на естественных субстратах. Расстояние между этими поселениями значительно больше, чем на Урале, и составляет 15-50 км. Аналогичная ситуация наблюдается в условиях поймы заповедника. Здесь постоянно, в 15 км друг от друга, существуют 2 жилых поселения. Общение зверей с весны до середины лета возможно только через боровую террасу и кратковременно заливаемые половодьем луга. В больших массивах елово-пихтовой тайги поселения барсука редки. Для безопасных перемещений звери устраивают временные норы на гривах верховых болот. Интересно, что при перемещении по припойменной территории барсук редко отрывает временные норы и, в большинстве случаев пользуется старыми бобровыми жилищами (поселениями). При этом в пойме, на боровой террасе и на водораздельном участке территориальные метки (кормовые покопки, «туалеты», заготовки под норы и т.д.) слабо выражены.
Как уже говорилось выше, возможность устройства выводковых нор является для барсука основным лимитирующим фактором. В силу особенностей геоморфологии Ильменского заповедника и еще в большей степени заповедника «Нургуш» возможности устройства поселений ограничены. Это вынуждает животных использовать для устройства нор антропогенно нарушенные ландшафты. В Ильменском заповеднике около 16% поселений устроены барсуками в отвалах траншей, угольных ямах, копях. В заповеднике «Нур-гуш» большая часть поселений устроена в старых курганах смолокурен (52%), в гребнях траншей (22%), в заброшенных усадьбах бывших деревень (11%) и только 15% поселений устроены в естественных субстратах (гривах).
Воздействие барсуков на среду. Обследование состояния растительности на участке обитания барсуков показало, что их роющая деятельность оказывает влияние на структуру и состав травостоя. Особенно это выражено на территории старых, длительно используемых поселений.
На Урале выделяются два типа поселений. Первый тип встречается в предгорном районе и характерен для большинства областей обитания барсука. Поселения первого типа расположены неподалеку от воды на различного рода возвышенностях с песчано-глинистыми почвами. Здесь барсуки проявляют высокую роющую активность. В поселениях второго типа, приуроченных к горам и местам выходов горных пород, роющая деятельность животных не достигает больших масштабов, так как в качестве нор и отнорков используются естественные расщелины и пустоты.
В поселениях первого типа изменения состава травостоя значительны. Коэффициент видового сходства [38] контрольного и опытного участков здесь составил 51%. Это происходит за счет выпадения части видов, обычных для нетронутых участков остепнен-ного луга, и внедрения новых видов, несвой- ственных исходному сообществу – пустырника пятилопастного (Leonurus quinquelobatus Gilib.), крапивы двудомной (Urtica dioica L.), бодяка щетинистого (Cirsium setosum Bess.), марьянника гребенчатого (Melampyrum cristatum L.), будры плющевидной (Glechoma hederacea L.) и др. В поселении второго типа отмечено существенное сокращение числа видов по сравнению с контролем, но большее сходство видового состава (65%), так как здесь присутствуют только два вида, не отмеченные в контроле – крапива двудомная и звездчатка злаковая (Stellaria graminea L.).
Таблица 3. Структура надземной фитомассы травянистого яруса на поселениях барсуков (воздушно-сухой вес, г/0,25 м2)
|
Место расположения поселения |
Показатели |
Контроль |
Барсучье поселение |
||
|
M + m |
Доля от общ. запаса, % |
M + m |
Доля от общ. запаса, % |
||
|
Предгорные экосистемы |
Общие запасы фитомассы, в том числе: |
143,9 + 9,3 |
100 |
131,8 + 10,7 |
100 |
|
Биомассы |
49,1 + 3,9 |
34 |
60,7 + 4,9 |
46 |
|
|
Некромассы |
94,8 + 8,1 |
66 |
71,2 + 7,1 |
54 |
|
|
Горные экосистемы |
Общие запасы фитомассы, в том числе: |
18,7 + 2,3 |
100 |
7,0 + 0,9 |
100 |
|
Биомассы |
15,0 + 1,9 |
80 |
5,9 + 0,9 |
84 |
|
|
Некромассы |
3,7 + 0,6 |
20 |
1,1 + 0,32 |
16 |
|
|
Долинные экосистемы р. Вятки |
Общие запасы фитомассы, в том числе: |
22,7 + 0,8 |
100 |
14,9 + 0,9 |
100 |
|
Биомассы |
17,4 + 0,7 |
76,6 |
11,4 + 1,1 |
76,5 |
|
|
Некромассы |
5,3 + 1,3 |
23,4 |
3,5 + 0,5 |
23,5 |
|
В обоих типах поселений вновь появившиеся виды типично синантропные. В поселениях первого типа некоторые из них (крапива, бодяк) становятся доминантами. В целом присутствие синантропных видов в поселениях связано с наличием здесь удобных экотопов, какими являются выбросы грунта из нор, обогащенные органикой. Структура и запасы надземной фитомассы изученных сообществ приведены в табл. 3
Видно, что продуктивность фитомассы на поселениях барсука и на контроле в предгорной части Ильменского заповедника (поселения первого типа) намного выше, чем в заповеднике «Нургуш». В горной части Ильменского заповедника (поселения второго типа), напротив эти показатели ниже, что как уже оговаривалось, связано с выходами скальных пород.
В пределах поселений первого типа запасы фитомассы выше, чем на контроле за счет мощного разрастания синантропных видов. Так, на долю только крапивы и бодяка здесь приходится 50% всей биомассы. Несколько меньше показатели запасов некромассы в поселении, что связано с более интенсивными темпами ее разложения, вытаптыванием. В поселении второго типа отмечено резкое сокращение всех показателей запасов фитомассы по сравнению с контролем, что можно, видимо, объяснить незначительным участием (менее 10% по биомассе) в травяном покрове поселения синантропных видов, предпочитающих рыхлый субстрат. Однако струк- тура фитомассы существенно не отличается от контроля. В лесостепи (Троицкий заказник) около поселения барсуков было 29 видов растений, по встречаемости доминировали Helictotrichon schellianum Hack., Artemisia pontica L., Veronica spuria L., Salvia stepposa Shost. и т.д. На поселении отмечено только 13 видов растений: Euphorbia virgata Waldst. et Kit., Salvia stepposa Shost., Leonurus quinquelobatus Gilib., Seseli libanotis L., Helictotrichon schellianum Hack., Fragaria viridis West. и т.д.
В долине р. Вятка на территории заповедника «Нургуш» сохранились останцы древних террас. Именно здесь располагаются барсучьи «городки». Здесь места при входах в норы и вершины холма поселения на общей площади 15 мІ лишены травянистого покрова и местами подстилки. На остальной части холма произрастают Oxalis acetosella L., Aegopodium podagraria L., Rubus idaeus L., Urtica dioica L., Pteridium aquilinum L.. С 1997 по 2005 г. в этом поселении барсуки обитали и зимовали не ежегодно. Несмотря на то, что в результате антропогенных причин заселенность барсучьих «городков» в долине р. Вятка не регулярна, в местах поселений, как и на Урале, происходят те же процессы в запасах фитомассы у нор и на тропах, отмечается присутствие видов, появившихся в результате жизнедеятельности барсуков. Подобная ситуация прослеживается в тайге Архангельской области. На барсучьем поселении с 14 входами, которое имеет возраст более 8 тыс. лет и занимает 400 мІ площади, выбросы почвы поросли Rosa sp., Calamagrostis sp., Chamaenerium angustifolium L., Urtica urens L. в окружении сосняка лишайникового с участками Vaccinium vitis-idaea L. [13].
Выводы
Таким образом, специфика экологии барсуков лесной и лесостепной зон связана в первую очередь с гидротермическим (природ- но-географическим или зональным) фактором. В целом характер изменения показателей плотности населения барсука, степени развития и количества «потенциально жилых» поселений соответствует характеру изменений общей биопродуктивности экосистем соответствующих природно-географических зон и подзон. Эти показатели в пределах лесной зоны возрастают в направлении север-юг (оптимизация термического режима), затем в лесостепной зоне снижаются в том же направлении (ухудшение режима увлажнения).
Список литературы Природно-географические особенности экологии барсука (Meles Brisson, 1762)
- Абрамов А.В., Вехник В.П. Таксономический статус барсука (Mustelidae, Meles) Самарской Луки//Заповедное дело России: принципы, проблемы, приоритеты: Материалы Междунар. науч. конф. Бахилова Поляна, 2003. Т. 1.
- Абрамов А.В., Савельев А.П., Сотников В.Н., Соловьев В.А. Распространение двух видов барсуков (Mustelidae, Meles) в европейской части России//Систематика, филогения и палеонтология мелких млекопитающих. СПб., 2003.
- Атлас СССР. М.: ГУГК, 1985.
- Благовещенская Н.В. Динамика лесных экосистем верхнего плато Приволжской возвышенности в голоцене//Экология. 2006. № 2.
- Верещагин Н.К., Громов И.М. Формирование териофауны СССР в четвертичном периоде//Успехи современной териологии. М.: Наука, 1977.
- Гептнер В.Г., Наумов Н.П., Юргенсон П.Б. и др. Морские коровы и хищные//Млекопитающие Советского Союза. Т. 2. Ч. 1. М.: Высш. шк., 1967.
- Горчаковский П.Л. Основные проблемы исторической фитогеографии Урала//Тр. Ин-та экологии растений и животных УФ АН СССР. Вып. 66. Свердловск, 1969.
- Горшков П.К. Барсук в биоценозах Республики Татарстан. Казань, 1997.
- Дворников М.Г. Оценка воздействия млекопитающих на почвенный покров в лесных биогеоценозах//Горные экосистемы Урала и проблемы рационального природопользования. Свердловск: УНЦ АН СССР, 1986.
- Дворников М.Г. К питанию хищных зверей Южной тайги//Териофауна России и сопредельных территорий (VII съезд териологического общества). М., 2003.
- Дворников М.Г. Комплексные экологические исследования в пойменных биогеоценозах р. Вятка//Бюл. Самарская Лука: Бюл. 2004. № 15.
- Дворников М.Г., Дворникова Н.П., Коробейникова В.П. Особенности растительности в поселениях барсука (Meles meles L.) на Южном Урале//Экология. 1996. № 5.
- Динесман Л.Г. Изучение истории биогеоценозов по норам животных. М.: Наука, 1968.
- Каталог млекопитающих СССР (плиоцен-современность). Л.: Наука, 1981.
- Кинд Н.В. О методах изучения климатов голоцена//Частные методы изучения истории современных экосистем. М.: Наука, 1979.
- Коробейников Ю.И. Численность и биомасса дождевых червей (Limbricidae) в лесах Ильменского заповедника//Биогеоценологические исследования на Южном Урале. Свердловск: УНЦ АН СССР, 1978.
- Коробейников Ю.И. Жужелицы Ильменского заповедника и их сезонная динамика активности//Структурно-функциональные взаимосвязи в биогеоценозах Южного Урала. Свердловск: УНЦ АН СССР, 1979.
- Косарев М.Ф. К истории взаимоотношений человека и природы в Западной Сибири (по материалам археологических исследований)//Антропогенные факторы в истории развития современных экосистем. М: Наука, 1981.
- Кузьмина Е.Н. Сравнение видового состава позднеплейстоценовых млекопитающих западных и восточных склонов Среднего Урала//Млекопитающие Уральских гор (информационные материалы). Свердловск: УНЦ АН СССР, 1979.
- Лихачев Г.Н. Некоторые черты экологии барсука в широколиственном лесу Тульских засек//Сб. материалов по результатам изучения млекопитающих в государственных заповедниках. М., 1956.
- Миронов Б.А. Поступление и разложение опада в сосняках и березняках в зависимости от гидротермического режима почв//Биогеоценологические исследования на Южном Урале. Свердловск: УНЦ АН СССР, 1978.
- Наумов Н.П. Биологические (сигнальные) поля и их значение в жизни млекопитающих//Успехи современной териологии. М.: Наука, 1977.
- Петренко А.Г. Древнее и средневековое животноводство среднего Поволжья и Предуралья. М.: Наука, 1984.
- Рябова Т.П. Развитие растительности Башкирского Предуралья в голоцене//Науч. доклады Высш. шк. Биол. науки. 1965.
- Смирнов Н.Г., Большаков В.Н., Косинцев Л.А. и др. Историческая экология животных гор Южного Урала. Свердловск: Из-во УрО АН СССР, 1990.
- Смирнов Н.Г., Косинцев Л.А., Бородин А.В. Влияние хозяйственной деятельности древнего населения на экосистемы Западной Сибири и Зауралья//Антропогенные факторы в истории развития современных экосистем. М.: Наука, 1981.
- Смирнов Н.Г., Уразова А.В., Косинцев П.А., Бородин А.В., Петрин В.Т. Игнатьевская пещера -новое местонахождение ископаемой териофауны Южного Урала//Исследования актуальных проблем териологии (информационные материалы). Свердловск: УНЦ АН СССР, 1983.
- Смирнов В.В., Чащин П.В. Сезонные изменения встречаемости дождевых червей (Lumbricidae) в рационе барсука (Meles, Mustelidae)//Экология: от генов до экосистем. Материалы конф. молодых ученых. Екатеринбург: Изд-во «Академкнига», 2005.
- Старков В.Ф. Хозяйство населения лесного Зауралья в эпоху неолита и энеолита//Антропогенные факторы в истории развития современных экосистем. М.: Наука, 1981.
- Терновский Д.В. Биология куницеобразных (Mustelidae). Новосибирск: Наука, 1977.
- Туганаев В.В. Многовековая динамика агроценозов Волжско-Камского края//Антропогенные факторы в истории развития современных экосистем. М.: Наука, 1981.
- Ушков С.Л. Звери и птицы Ильменского заповедника. Екатеринбург, 1993.
- Ходашева К.С. Динамика биомассы позвоночных животных и ее связь с зональными особенностями фитомассы и водно-теплового режима//Биологическая продуктивность и круговорот химических элементов в растительных сообществах. Л.: Наука, 1971.
- Чащин П.В. Некоторые факторы, определяющие пространственно-временную организацию поселений барсука в Ильменском заповеднике//Материалы конф. мол. ученых-экологов Уральского региона «Современные проблемы популяционной, исторической и прикладной экологии». Екатеринбург, 1998.
- Чащин П.В. Роль дождевых червей в питании барсука на Южном Урале//Материалы междунар. конф. «Экологические проблемы горных территорий». Екатеринбург, 2002.
- Шаландина В.Г. Растительный покров северо-запада Татарии в позднем голоцене и его изменения под влиянием антропогенных факторов//Антропогенные факторы в истории развития современных экосистем. М.: Наука, 1981.
- Kruuk H., Parish T. Feeding specialization of the European badger Meles meles in Scotland//J. of Animal Ecology. 1981. V. 50, № 3.
- Sorensen T. A method of establishing groups of equal amplitude in pland sociology based on similarity of species content//K. danske vidensh Selsh. 1948. S(4).
- Vigros E., Casanovas J.G. Environmental constraints at the edge of a species distribution, the Eurasian badger (Meles meles L.): a biogeographic approach//J. of Biogeography. 1999. V. 26.
