Прижизненные захоронения морских донных сообществ в венлокских штормовых отложениях на поднятии Чернова

Автор: Безносова Т.М., Лукин В.Ю., Матвеев В.А.
Журнал: Вестник геонаук @vestnik-geo
Рубрика: Научные статьи
Статья в выпуске: 12 (264), 2016 года.
Бесплатный доступ
Результаты изучения разреза Безымянный на поднятии Чернова позволяют предположить штормовой режим седиментации с преобладанием волновой турбулентности в венлоке и относительно высокие темпы осадконакопления. В разрезе выделены четыре интервала с прижизненными захоронениями донных сообществ и остатков минерализованной биоты, замещенных пиритом в строматолитах. Образование пирита связывают с аноксидными условиями, создавшимися сразу после гибели организмов. Полученные результаты расширяют возможность реконструкции эволюции древних экосистем в раннем силуре.
Захоронение фауны, штормовые отложения, брахиоподы, табуляты, строматолиты
Короткий адрес: https://sciup.org/149129191
IDR: 149129191 | УДК: 550.7: | DOI: 10.19110/2221-1381-2016-12-31-36
In-situ burials of benthic communities in wenlock storm sediments of the Chernov swell
The represented data suggest storm mode of sedimentation with predominance of wave turbulence in the Wenlock. We found unique in-situ burials of benthic communities and remains of mineralized biota in stromatolites, replaced by pyrite, which formation had been caused by anoxic conditions after the organism death. The obtained results support reconstruction of evolution of the Early Silurian ecosystems.
Текст научной статьи Прижизненные захоронения морских донных сообществ в венлокских штормовых отложениях на поднятии Чернова
Анализ характера захоронений фаунистических сообществ в изученных Тимано-Уральских разрезах силура свидетельствует о значительном влиянии изменений седиментационных обстановок на развитие и территориальное распространение бентосных организмов в венлокском морском бассейне. Смена обстановок была связана с возможностью миграции или изолированностью бассейна, течениями, штормами, увеличением поступления терригенного материала, изменениями рельефа дна и глубины бассейна. По мере масштабности проявления абиотических факторов выделяются различные типы захоронений бентосных организмов в изученном разрезе Безымянный [2].
Захоронения органических остатков в прижизненном положении или вблизи места их обитания встречаются редко. Одной из возможных причин возникновения таких захоронений считаются штормы [7], во время которых происходило погребение части донного сообщества в прижизненном положении. Например, в этих захоронениях присутствуют скопления целых сочлененных раковин разных размеров и различных возрастных стадий, у которых сохраняются тонкая скульптура, шлейфы, тонкие иглы, элементы внутреннего строения. Другая часть раковин при этом может переотлагаться и смешиваться с фрагментами других сообществ — мелких остракод, гастропод.
Обсуждение результатов
Венлокский возраст толщи, слагающей разрез по ручью Безымянному, впервые был определен по многочисленным остаткам брахиопод и остракод, которые присутствуют почти от его основания до кровли [1]. Последующее изучение состава отложений этого разреза и характера захоронений органических остатков позволили сделать вывод о неста-
Рис. 1. Местонахождение разреза венлокских отложений по руч. Безымянному (широта 67°53'29.94"С, долгота 62°22'23.30"В)
Fig. 1. Location of Wenlockian outcrop near Bezymyanny Brook (latitude 67°53'29.94"N, longitude 62°22'23.30"E)
бильных условиях осадконакопления в венлокское время, что подтвержается периодическими остановками роста биостромов, интенсивным выпадением илового, био- и литокластового материала и его переотложений [3, 10, 13].
Разрез по ручью Безымянному мощностью 197 м (рис. 1, 2, Г) характеризуется таксономическим разнообразием ископаемой биоты, представленной брахи-оподами, гастроподами, остракодами, пелициподами, члениками криноидей, реже ортоцератидами и трилобитами, а также конодонтами Ozarkodina kozhimica Melnikov , Oz. waugoolaensis (единичный уровень) , Oulodus rectangulus , Oul. kozhimicus , Nudibelodina sensiti-va, Pterospathodus sp. A, Apsidognathus sp. лландоверий-ско-венлокского возраста, которые определены в толще мощностью 125 м [3]. В отдельных прослоях были обнаружены фораминиферы [1] и кораллово-строма-топоровая фауна, которая формировала два биострома в нижней и верхней части разреза (рис. 3, А).
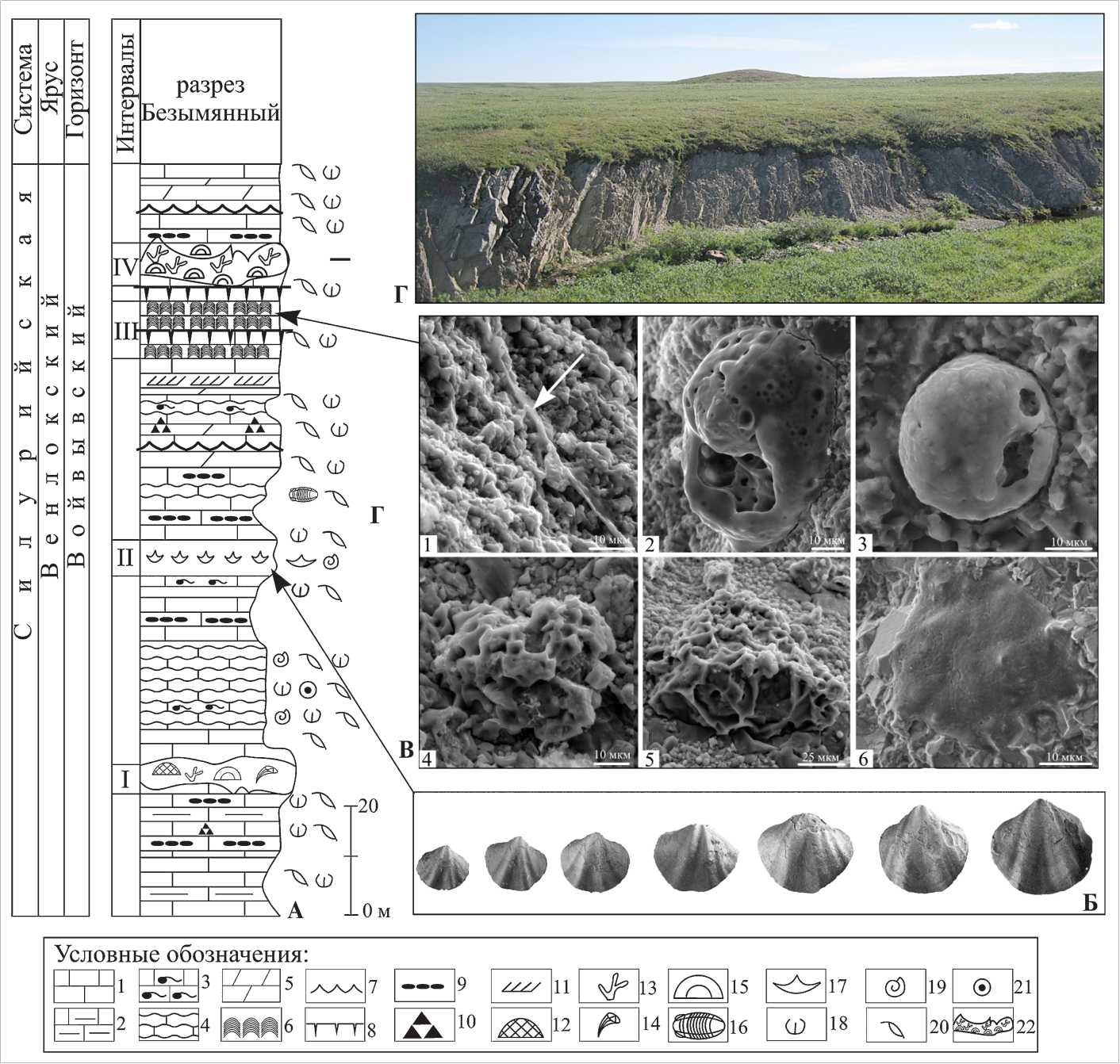
Рис. 2. Стратиграфическое распространение фаунистических остатков и захоронений in situ в разрезе Безымянный: А — схематичное строение разреза Безымянный; Б — брахиоподы Spirinella nordensis (Ljashenko) — возрастной ряд (раковины из линзы in situ). Образец 2/007, х1; В — остатки фоссилизированных микроорганизмов в венлокских строматолитах: фиг. 1 — минерализованный чехол трубчатого нитевидного образования; фиг. 2 — овальная форма; фиг. 3 — шаровидная форма со сглаженной мелкобугристой поверхностью; фиг. 4 — шаровидная форма с губчатой поверхностью; фиг. 5 — куполообразные фоссилии; фиг. 6 — фрагмент микробной биопленки; Г — Выходы пород войвывского горизонта. Условные обозначения: 1 — известняк, 2 — глинистый, 3 — биотурбированный, 4 — комковатый, 5 — мергели, 6 — строматолиты, 7 — знаки волновой ряби, 8 — трещины усыхания, 9 — плоскогалечники, 10 — конгломерат, 11 — косая слоистость, 12 — массивные табуляты, 13 — ветвистые табуляты, 14 — ругозы, 15 — строматопороидеи, 16 — трилобиты, 17 — брахиоподы, 18 — фрагменты брахио-под, 19 — гастроподы, 20 — остракоды, 21 — криноидеи, 22 — биостром
Fig. 2. Stratigraphic development of fauna remains and burials in situ in Bezymyanny section: A — Schematic structure of section Bezymyanny; Б — Brachiopods Spirinella nordensis (Ljashenko) — age series (shells from lens in situ). Sample 2/007, x1; B — remains of fossilized microorganisms in Wenlockian stromatolites: Fig. 1 — mineralized cover of tubular filamentary form; Fig. 2 — oval form; Fig. 3 — spherical form with smooth hummocky surface; Fig. 4 — spherical form with a spongy surface; Fig. 5 — dome-like fossils; Fig. 6 — fragment of a microbial biofilm; Г — outcrops of oyvyvsky horizon in section Bezymyanny. Legend: 1 — limestone 2 — clay, 3 — bio-turbated, 4 — lumpy, 5 — marls, 6 — stromatolites, 7 — signs of wave ripples, 8 — mud cracks, 9 — flat pebbles, 10 — conglomerate, 11 — cross-bedding, 12 — massive tabulates, 13 — branching tabulates, 14 — rugoses, 15 — stromatoporoids, 16 — trilobites, 17 — brachiopods, 18 — fragments of brachiopods, 19 — gastropods, 20 — ostracods, 21 — crinoids, 22 — biostrom
Отложения, слагающие разрез Безымянный, и захоронения органических остатков свидетельствуют об активной гидродинамической среде неглубокого морского бассейна. Это подтверждает характер слоистости, обилие окатанных галек и створок раковин брахиопод, обломков гастропод и трилобитов, переслаивание тонкоплитчатых известняков, известковых глин со следами илоедов, известняков иловых с тонким детритом и неотчетливой слойчатостью, мергелей и известняковых гравелито-песчаников, биокластовых известняков с крупными гальками (рис. 2, Д, Ж, 3), остракодово-иловых известняков, брахиоподовых и остракодово-водорослевых (микробиальных) известняков, ракушечников острако-довых (рис. 3, Г), плоскогалечниковых конгломератов, мощность прослоев которых изменяется от 3 см до нескольких десятков см, известняков с пелоида-ми, биокластовых известняков с текстурами взвихривания — турбулентности среды (рис. 3, И), известняков строматолитовых с уплощенными гальками,

Рис. 3. Колония табулят и микроструктуры известняков в разрезе Безымянный: А — табуляты Aulocystella aseptata (Barskaja, 1967), продольное сечение, обр. 149; Б — пелоидный известняк с плоскими интрокластами; В — строматолитовый известняк с «линзами» плоскогалечников; Г — пелитоморфный известняк с личинками остракод; Д — известняк биокластовый с фрагментами криноидей, остракод и брахиопод; Е — известняки полосчатые с градационной слоистостью; Ж — известняк биокластовый с крупными фрагментами криноидей, гастропод, остракод и брахиопод; 3 — известняк литобиокластовый; И — известняк детритовый, биоморфный с полуокатанными хаотичными штормовыми выносами
Fig. 3. Tabulate colony and limestone microstructures in Bezymyanny section: A — tabulates Aulocystella aseptata (Barskaja, 1967), longitudinal section, sample 149; Б — peloid limestone with flat introclasts; B — stromatolite limestone with pebble «lenses»; Г — pelitomor-phic limestone with ostracod larvae; Д — bioclastic limestone with fragments of crinoids, ostracods and brachiopods; E — banded limestone with graded bedding; Ж — bioclastic limestone with large fragments of crinoids, gastropods, ostracods and brachiopods; 3 — litho-bioklastic limestones; И — detrital limestone, biomorphic with subrounded chaotic storm cone имеющими признаки взмучивания осадка (рис. 3, В), известняков с «линзами» плоско галечников (рис. 3, Б), известняков полосчатых с градационной слоистостью (рис. 3, Е).
В этом разрезе исключение составляют четыре интервала, которые формировались в условиях нормализации морских условий в бассейне и благодаря которым можно наблюдать последовательную смену экологических доминантных сообществ от нижних слоев разреза до его кровли, а также изменения глубин обитания сообществ. Эти захоронения фиксируют периоды стабилизации условий осадконакопления, во время которых формировались сообщества кораллов, брахиопод, а также строматолитовые постройки в приливно-отливной зоне [3, 9, 11].
Первый интервал мощностью 0.5 м представлен кораллово-строматопоровым биостромом в нижней части разреза, в котором доминируют тонковетвистые Parastriatopora и Striatopora, колонии табулят Riphaeolites lamelliformis Klaamann и мелкотрубчатые Romingeria sp., а также колонии стромотопороидей и одиночные ругозы.
Второй интервал мощностью 4.4 м с захоронением брахиопод Spirinella nordensis (Ljashenko) выделен в средней части разреза. Изучавший разрез Безымянный в 1971 г. Н. И. Тимонин отметил в своем полевом дневнике этот интервал как «брахиоподовый горизонт». Брахиоподы монотаксонного сообщества S. nordensis отличаются хорошей сохранностью целых раковин различных стадий роста от ювенильных до зрелых (рис. 2, Б) с сочлененными створками, с сохранившимися элементами внутреннего строения, тонкими пластинами нарастания и иглами на поверхности раковин. Проведенные биохимические исследования раковин брахиопод S. nordensis из этого захоронения зафиксировали отчетливо выраженный в них максимум содержания аминокислот, достигающий более 100 мкг/г [4]. Хорошая сохранность раковин позволила исследовать их морфологические особенности , которые свидетельствуют о возможности адаптации этих брахиопод к различным условиям существования. Раковины S. nordensis относятся к экологическому якорному типу и батиметрически — к третьему бентосному комплексу А. Буко [5]. Загнутая макушка наряду с ножкой и развитой ареей способствовали фиксации раковины в нужном положении на твердом субстрате, а тонкие иглы и складки на поверхности раковины помогали удерживаться ей на мягком субстрате. Брахиоподы S. nordensis в разрезе Безымянном являются доминирующими остатками бентосной фауны по численности раковин и распространены в разрезе с первого слоя до его кровли. Но в отличие от описанного захоронения во втором интервале, в прослоях известняков с линзами плоскогалечного конгломерата, известняков биокластовых и с текстурами взвихривания (среды турбулентности) (рис. 3, И) они представлены преимущественно отдельными створками раковин, а также ракушечниками в виде лома раковин совместно с остракодами, гастроподами, пелециподами, члениками криноидей и трилобитами (рис. 3, Ж).
Третий интервал мощностью 22.4 м, заключающий строматолитовые постройки, сложен чередованием маломощных прослоев известняков тонко- 34
слойчатых, плитчатых, с остатками раковин остра-код, брахиопод, гастропод и известняков со строматолитами. В этом интервале впервые на территории Европейского Северо-Востока были обнаружены остатки строматолитообразующей биоты (рис. 2, В). Микрозондовый анализ показал, что эти остатки замещены пиритом [11]. Уникальная сохранность обнаруженных остатков биоты, обусловлена, вероятно, мгновенным захоронением части строматолитовых построек. Свидетельством такой сохранности также служит установленный элементный состав шаровидных фоссилий, замещенных пиритом (рис. 2, В, фиг. 3). Образование пирита связано с аноксидны-ми условиями, которые создавались сразу после гибели организмов. Известно, что в пирите могут быть законсервированы химические соединения, аминокислоты и некоторые другие органические молекулы [8]. Экспериментальное изучение образования пирита путем замещения организмов была осуществлена группой исследователей во главе с С. Дэрроу [18].
Четвертый интервал представляет собой биостром в верхней части разреза, в основании которого залегает прослой (0.35 м) с колониями строма-топороидей. Выше биостром мощностью 0.9 м сложен колониями трубчатых табулят Aulocystella aseptata (Barskaja) (рис. 3, А) и массивными Riphaeolites lamelliformis Klaamann. Биостром перекрывают известняки тонкоплитчатые с трещинами усыхания осадка (2.5 м) и знаками волновой ряби, в кровле которых залегают известняки со строматопороидеями.
Заключение
В разрезе Безымянном установлены интервалы с уникальными захоронениями сообществ донных организмов обитавших «in situ», которые фиксируют батиметрические изменения условий осадконакопления и важны для стратиграфических выводов и корреляции изученных отложений по ручью Безымянному с разрезами западного склона Приполярного Урала, поднятия Чернышева, островов Долгий, Сев. Земля, Сааремаа, Готланд, а также Таймыра, Южного Верхоянья, Зеравшано-Гиссарской области, Сибирской платформы [6, 16].
Известно, что ранний венлок в разрезах Эстонии, Сибирской платформы, на островах Таймыр, Северная Земля, Долгий, на Аляске знаменуется расцветом коралловой фауны [12, 14, 17, 19]. Но на территории западного склона Урала, поднятиях Чернова и Чернышева коралловая фауна представлена только поздневенлокскими ассоциациями табулят Riphaeolites, Parastriatopora и Aulocystella [3, 9] . Этот факт может служить палеонтологическим подтверждением установленного нами ранее перерыва в осадконакоплении на рубеже лландовери и венлока в разрезе западного склона Приполярного Урала и отсутствия нижней части венлока в североуральских разрезах [2].
Работа выполнена при поддержке Программы фундаментальных исследований РАН, проект № 15-18-537 «Тимано-Североуральский палеобассейн в палеозое. Закономерности динамики биотических комплексов, палеобиогеография и палеоклиматология».
Список литературы Прижизненные захоронения морских донных сообществ в венлокских штормовых отложениях на поднятии Чернова
- Антошкина А. И., Безносова Т. М. Новые данные по стратиграфии венлокских отложений Большеземельской тундры//Бюл. Моск. об-ва испытателей природы. Отд. геол. 1988. Вып. 6. Т. 63. С. 32-39.
- Безносова Т. М. Сообщества брахиопод и биостратиграфия верхнего ордовика и нижнего девона северо-восточной окраины палеоконтинента Балтия. Екатеринбург: УрО РАН, 2008. 218 с.
- Безносова Т. М., Мянник П., Майдль Т. В., Лукин В. Ю., Матвеев В. А. Условия осадконакопления и биота на рубеже лландовери и венлока (поднятие Чернова)//Вестник Института геологии Коми научного центра УрО РАН. 2014. № 3. С. 14-18.
- Безносова Т. М., Шанина С. Н. Биохимические индикаторы геобиологических событий в раннем палеозое на Европейском Северо-Востоке//ДАН. 2010. Т. 434. № 1. С. 76-78.
- Буко А. Эволюция и темпы вымирания. М.: Мир, 1979. 378 с.
